 2022, Vol. 42
2022, Vol. 42文章信息
- 吴旭, 牛耀彬, 荀梦瑶, 金俊逸, 李娜艳, 唐亚坤, 陈云明
- WU Xu, NIU Yaobin, XUN Mengyao, JIN Junyi, LI Nayan, TANG Yakun, CHEN Yunming
- 黄土丘陵区优势造林树种水分来源对季节性干旱的响应
- Responses of water source to seasonal drought of dominant afforestation tree species in the loess hilly region of China
- 生态学报. 2022, 42(10): 4101-4112
- Acta Ecologica Sinica. 2022, 42(10): 4101-4112
- http://dx.doi.org/10.5846/stxb202106221657
-
文章历史
- 收稿日期: 2021-06-22
- 网络出版日期: 2022-01-10





2. 西北农林科技大学水土保持研究所, 杨凌 712100;
3. 中国科学院水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100
2. Institute of Soil and Water Conservation, Northwest Agriculture Northwest Agriculture and Forestry University and Forestry University, Yangling 712100, China;
3. State Key Laboratory of Soil Erosion and Dry Land Farming on the Loess Plateau, Institute of Soil and Water Conservation, Chinese Academy of Sciences, Ministry of Water Resources, Yangling 712100, China
由于天然植被的严重破坏及不合理的土地利用导致黄土丘陵区成为中国水土流失最严重的地区之一。不仅成为困扰该区社会经济可持续发展的主要问题, 而且也给黄河下游地区带来一系列生态环境问题。为改善生态环境, 再造一个山川秀美的大西北, 1999年以来该区开展了大规模退耕还林工程, 然而, 受温带半干旱气候的影响, 当地多年平均降水量(≤600 mm)远小于潜在蒸发量(1000 mm)[1], 且主要集中在7-9月, 植物生长易受季节性干旱的影响。土壤水分季节性亏缺是部分人工林出现“三低”问题、“小老头树”现象和碳汇转为碳源的主要原因[2]。因此, 亟需深入分析人工林水分利用稳定性, 才能全面评价该区水土流失治理和生态恢复的可持续性。
油松(Pinus tabuliformis)和刺槐(Robinia pseudoacacia)具有较强适应性和抗逆性, 是黄土丘陵区造林优势树种, 在区域水土流失治理和景观维持方面发挥着重要作用。然而, 由于生态恢复初期缺乏科学指导, 营造了大面积人工纯林, 导致林木生长不良、形成土壤干层、碳汇功能减弱等生态效益低下等问题日益突出。经过长期造林实践, 沙棘(Hippophae rhamnoides)因具有较强固氮能力和较高经济价值, 且沙棘和油松、沙棘和刺槐混交林的水土保持效果和土壤肥力等明显高于纯林, 该混交模式被广泛应用[3]。目前, 已有研究多通过盆栽试验或根系分布的调查, 分析水分环境变化对纯林中沙棘、油松和刺槐的影响[4-5]。然而, 根系分布并不总决定植物的水分利用状况[6], 此外, 受季节性干旱的影响, 当地水分的供给能否满足植物生长需求, 混交林中沙棘和油松、沙棘和刺槐是否存在水分竞争, 以及沙棘+油松和沙棘+刺槐混交林是否比纯林更具有水分利用的稳定性和可持续性还需进一步分析。
植物主要通过活性根吸收水分, 传统的根系调查法存在费时耗力, 对植被及其生态环境破坏性大, 有一定误差等问题, 且对于深根系植物来说, 在有多个可能水分来源时, 通过直接方法来确定不同水源对植物的贡献率就显得更加困难[7]。氢氧稳定同位素分馏理论以及基于“稳定同位素质量守恒”多元线性混合模型的发展, 被认为是对植物和土壤破坏较低而精确度较高的方法, 已广泛应用于环境脆弱区植物水分来源的研究[8-9]。黄土丘陵区地处干旱半干旱过渡带, 降水、土壤储存水是植物的主要水分来源[10]。研究表明, 纯林中植物水分来源主要受根系分布和水分环境条件变化的影响[11]。草本植物多使用浅层土壤水且易受干旱的影响, 但木本植物往往有能力从更深土层获取水源。此外, 一些植物为了适应季节性干旱而发展出二态性根系, 旱季主要使用深层土壤水, 湿润季则主要使用浅层土壤水或降水[12]。而在水分受限的混交林中, 共生植物之间可能存在水分竞争, 也可能通过“水分利用生态位分离”避免水分竞争[13]。因此, 相对于纯林, 混交林共生植物之间水分利用的分析还需要考虑其水分利用协调关系, 不能将纯林中植物的水分利用结果直接应用到混交林中。
安塞国家生态试验站位于黄土高原中部典型黄土丘陵沟壑区, 土地类型多样, 资源丰富, 是水土保持与生态环境建设科学研究和试验示范推广基地。本文以试验站固定样地为基础, 选取沙棘+油松(HrPt)、沙棘+刺槐(HrRp)混交林为对象, 以沙棘(Hr)、油松(Pt)和刺槐(Rp)纯林为对照, 使用氢氧稳定同位素技术, 采用IsoSource模型量化植物水分来源, 借助当地季节性干旱事件, 分析不同水分环境下植物水分来源变化特征;结合相似性比例指数(PS), 阐明混交林中沙棘与油松、沙棘与刺槐水分利用关系, 揭示其对干旱环境的适应机制, 为进一步评估区域生态恢复过程中人工林水分利用稳定性提供基础数据。
1 材料与方法 1.1 研究区概况研究区位于陕西安塞区境内, 试验地选在中国科学院安塞水土保持综合试验站山地试验场(109°19′E, 36°51′N, 海拔1068-1309 m), 属典型黄土丘陵沟壑区;该区具有典型暖温带半干旱气候, 1985-2017年均气温8.8 ℃, 年均降水量(458±172.5)mm, 年际年内分布不均;年均潜在蒸发1000 mm左右。延河在安塞境内贯穿南北, 流域面积占总面积的89.8%。地下水位埋深较大, 含水层厚度一般100-150 m。土壤类型以黄绵土为主, 质地为砂壤, 土壤颗粒粒径2.00-0.02 mm, 0.02-0.002 mm和 < 0.002 mm含量依次为65%, 24%和11%;土壤pH 8.4-8.6。主要木本植物有沙棘、油松、刺槐、侧柏(Platycladus orientalis)等;草本主要有白羊草(Bothriochloa ischaemum)、铁杆蒿(Artemisia gmelinii)、甘草(Glycyrrhiza uralensis)等。
1.2 研究方法 1.2.1 试验材料以山地试验场野外长期固定监测林地为基础, 选取沙棘+油松(HrPt)、沙棘+刺槐混交林(HrRp)及沙棘(Hr)、油松(Pt)、刺槐(Rp)纯林为对象。各样地均为2000年左右栽植的人工林, 样地调查于生长季前进行(4月), 基本特征见表 1:
| 样地类型 Types of sample plots |
树种 Species |
海拔 Altitude/m |
坡度 Slope/(°) |
坡向 Aspect |
土壤容重 Soil bulk density/(g/cm3) |
树高 Tree height/m |
胸径/地径 DBH/DGH/cm |
冠幅 Crown width/m2 |
| HrPt | 沙棘H. rhamnoides | 1270 | 12 | 阳坡 | 1.25±0.02 | 3.08±0.14 | 5.43±0.33 | (2.01±0.09)×(2.04±0.11) |
| 油松P. tabuliformis | 4.01±0.17 | 7.11±0.48 | (2.62±0.12)×(2.60±0.14) | |||||
| HrRp | 沙棘H. rhamnoides | 1302 | 16 | 阳坡 | 1.29±0.04 | 2.69±0.10 | 4.63±0.32 | (1.79±0.10)×(1.80±0.09) |
| 刺槐R. pseudoacacia | 5.92±0.52 | 7.80±1.02 | (3.36±0.35)×(3.25±0.34) | |||||
| Hr | 沙棘H. rhamnoides | 1267 | 16 | 阳坡 | 1.33±0.06 | 3.46±0.12 | 5.96±0.33 | (2.40±0.12)×(2.29±0.11) |
| Pt | 油松P. tabuliformis | 1189 | 12 | 阳坡 | 1.33±0.05 | 3.69±0.09 | 7.43±0.27 | (2.96±0.12)×(3.36±0.11) |
| Rp | 刺槐R. pseudoacacia | 1287 | 18 | 阳坡 | 1.34±0.02 | 7.59±0.65 | 9.40±1.50 | (4.36±0.39)×(4.24±0.32) |
| HrPt:沙棘+油松混交林Mixed forest of H. rhamnoides and P. tabuliformis;HrRp:沙棘+刺槐混交林Mixed forest of H. rhamnoides and R. pseudoacacia;Hr:沙棘纯林Pure forest of H. rhamnoides;Pt:油松纯林Pure forest of P. tabuliformis;Rp:刺槐纯林Pure forest of R. pseudoacacia;DBH:胸径Diameter at breast height;DGH:地径Diameter at ground height | ||||||||
(1) 降水样品:主要借助安塞山地自动气象站的雨量器。单次雨量超过5 mm的降水作为样品, 3次重复;采集时间为降水后10 min或夜间降水的次日清晨, 以减少蒸发作用的影响。水样装入采样瓶中(20 mL), 用Parafilm密封, 带回室内冷藏保存(1-4 ℃), 用于测定降水δD和δ18O。样品采集日期为2017年5-10月, 共采集到降雨28次。
(2) 植物茎秆:每种植物选取3株与表 1中所列生长指标相近且长势良好的植株, 采集其非绿色栓化小枝(直径0.3-0.5 cm, 长3-5 cm)。去皮后立即装入采样瓶中, 用Parafilm密封, 放入随身携带的冰盒中, 带回室内冰冻保存(低于-20℃), 用于测定植物茎秆水δD和δ18O。
(3) 土壤样品:在收集植物样品附近, 距离树基50 cm处按照不同深度(0-10、10-20、20-40、40-100、100-150、150-200 cm)使用土钻采集土壤样品, 重复3次。将其迅速放入采样瓶中, 用Parafilm密封, 放入随身携带的冰盒中, 带回室内冰冻保存, 用于测定土壤水δD和δ18O。
(4) 植物、土壤样品采集时间:于7月(旱季)和9月(湿润季), 选择在晴天、无云条件下的中午时刻(11:00-14:00)对植物、土壤等进行连续3天的样品采集, 以明确植物水分来源对季节性干旱事件的响应。
1.2.3 样品分析利用低温真空抽提系统(LI-2100), 对茎秆和土壤样品中的水分进行抽提。本研究中, 所有样品在80℃下经过40-90 min完成抽提, 抽提时间在规范范围之内。此外, 将部分抽提后的样品在80℃下烘干至恒重, 计算样品抽提效率。研究所使用的真空抽提系统抽提效率在98%以上, 水分抽提完全, 抽提过程中未发生同位素分馏[14]。利用该方法提取得到的各树种茎秆水及土壤水能够满足氢、氧稳定同位素测定要求。
抽提完之后, 利用质谱仪(Finnigan, MAT-253), 对所有水样进行氢氧稳定同位素分析(TC-AE法分析)。由于自然丰度的稳定性同位素含量极低, 因此国际上通常使用相对量表示稳定同位素的组成[15]。计算公式如下:
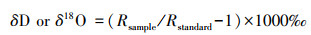
|
(1) |
式中, Rsample和Rstandard分别是样品和标准平均海水(SMOW)的2H/1H和18O/16O的摩尔比值。δ > 0, 表示样品的重同位素比标准物富集, 反之则比标准物贫化。
1.2.4 量化不同深度土层水分贡献率除极少数盐生或旱生植物以外, 大部分植物在根系吸水、运输至叶片蒸腾之前无同位素分馏现象[16]。因此, 通过比较不同深度土壤水和植物茎秆水的δD和δ18O值, 即可推断出不同树种对各土层土壤水的利用比例。然而, 通过直观法只能初步判断主要水源区域, 无法量化其贡献率。在源同位素比值能够测定情况下, 可利用二源混合模型来区分对每一个源的利用状况[17]。模型如下:
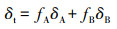
|
(2) |
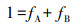
|
(3) |
式中, δt表示植物木质部的δD或δ18O(已知量), δA和δB表示A和B水源的δD或δ18O值(已知量), fA和fB表示来自A源和B源的比例(未知量)。两个未知量两个方程, 方程组有唯一解。同样, 可以同时利用δD或δ18O来确定植物对3种不同水源的利用比例。本研究抽提了6层土壤水, 由于0-20 cm和150-200 cm土壤层贡献较小, 因此, 在后期数据处理时, 对相邻土壤层进行合并, 即分析0-40 cm、40-100 cm及100-200 cm土层土壤水的贡献率。对于3个或3个以上不同水源, 计算每一个水源的利用比例会较为繁琐。因此, 本研究采用IsoSource模型[17], 确定各树种对不同水源的相对利用比例(具体步骤详见http://www.3mbang.com/p-3112242.html)。
1.2.5 相似性比例指数(PS)相似性比例指数是表征生态系统中共生植物生态位重叠的指标, PS指数越大, 表明共生植物生态位重叠的越多, 对某一资源的竞争越大;反之, 生态位重叠不明显, 对某一资源的竞争越小[18]。本研究利用PS指数评估混交林物种间对水资源的竞争现象。

|
(4) |
式中, PS表示相似性比例指数;p1i和p2i分别表示相比较的植物对第i种水源的利用比例。
1.2.6 根系调查以标准木主干为中心, 在其半径1 m的弧线正方位上选点, 用根钻(内径9 cm)对各树种根系分布特征进行调查, 基于前期已有研究基础, 初步确定取样深度为200 cm。根系分层取样(10 cm/层)并拣出相应土层的根系, 将细根归类编号后装入自封袋带回实验室。室内处理, 用蒸馏水清洗干净后用根系扫描仪(EPSON)对各层根进行扫描, 通过CIAS(Computer Image Analysis System)软件进行分析。最后置入烘箱75℃48 h, 烘干至恒重, 再分别称重和记录。
1.2.7 环境因子(1) 气象因子:主要来源于试验站的山地自动气象观测点。观测指标重点关注大气温/湿度(Vaisala, HMP45D)和降水量(Vaisala, DRD11A Rain Detector)等, 输出数据为30 min平均值。
(2) 土壤水分:在2块混交林样地及Hr、Pt样地内分别布设一套ECH2O土壤含水量监测系统, 长期监测土壤水分的动态变化。分层测量(0-10、10-20、20-40、40-100、100-150和150-200 cm), 输出数据为30 min平均值。此外, 采用土钻法对ECH2O数据进行校正以及Rp样地土壤含水量的测定。
1.2.8 数据处理采用SPSS 16.0在对氢、氧同位素数据进行正态分布和方差齐性检验的基础上, 采用One-Way ANOVA对其进行方差分析, 使用Sigma plot 10.0完成制图。
2 结果与分析 2.1 降水量、土壤水分及降水稳定同位素特征研究区2017年生长季(5-10月)总降水量为442.8 mm, 同期多年平均降水量(438.07±24.24)mm。其中7月22日-10月31日的降水量为312.6 mm, 占全年的70.6%;仅有3.16%的降水发生在6月6日-7月21日, 春末和夏初降水明显不足(图 1)。受降水年内分配不均的影响, 旱季取样时期(7月12-14日), Hr、Pt、Rp、HrPt、HrRp样地0-200 cm土层土壤含水量均值分别为:0.067、0.090、0.064、0.087、0.084 m3/m3。湿润季取样时期(9月17-19日), 各样地土壤含水量较旱季显著增加(P < 0.05)分别为:0.113、0.151、0.125、0.118、0.123 m3/m3。此外, 不同时期随土层深度的增加, 各样地土壤含水量整体呈现先增加后降低的趋势(图 2)。

|
| 图 1 研究区降水量和降水δ18O和δD值季节变化 Fig. 1 Seasonal variations of precipitation and the delta D, 18O isotope values of rainwater in the study area |

|
| 图 2 旱季和湿润季各样地0-200 cm土层土壤体积含水量剖面分布 Fig. 2 Vertical profiles (0-200 cm) of soil volumetric moisture in the sample plots during dry and wet seasons |
降水δD值的变化范围较大, 具有明显的季节变化规律。6月6日-7月21日, δD值较高, 均值为(-24.70‰);7月22日-8月31日降水频繁, δD相对贫化, 均值为(-57.97‰);9月-10月δD值平均为(-36.84‰)。降水δ18O值也具有较大变化, 介于(-10.22‰)-(2.76‰)之间(图 1)。进一步根据降水样品的δD和δ18O值, 拟合得出研究区大气降水线:δD=6.30δ18O-4.97(R2=0.902, P < 0.01)(图 3)。与全球大气降水线δD=8δ18O+10相比, 研究区降水的δ18O-δD坐标点基本落在其右下方, 即区域大气降水线的斜率和截距都较低, 且与全球大气降水线差异显著。此外, 植物茎秆水和土壤水线斜率和截距均显著低于区域大气降水线。

|
| 图 3 降水、土壤水、植物茎秆水的δD和δ18O的线性回归关系 Fig. 3 The linear regression relationship between δD and δ18O in precipitation, soil water and plant xylem water |
表 2为旱季和湿润季各样地不同层次土壤水氢、氧稳定性同位素的统计特征。不同时期各样地在0-40 cm土层土壤水的δ18O(δD)值较高, 且随土层深度增加, δ18O(δD)值逐渐降低(表 2)。旱季, 除HrRp外, 其余林分不同土层土壤水δ18O(δD)值差异显著(P < 0.05);湿润季, 仅Hr的δ18O(δD)、Pt的δD和HrRp的δ18O值不同土层间差异显著(P < 0.05)。研究区地下水埋深超过100 m, 土壤水的补给来源仅有天然降水。因此, 40 cm土层以下, 土壤水δ18O和δD值均显著小于降水δ18O和δD值(表 2)。
| 水分条件 Moisture conditions |
水源 Source |
土层深度 Soil depth/cm |
Hr | Pt | Rp | |||||
| δ18O/‰ | δD/‰ | δ18O/‰ | δD/‰ | δ18O/‰ | δD/‰ | |||||
| 旱季 | 降水 | -3.69 | -24.70 | -3.69 | -24.70 | -3.69 | -24.70 | |||
| Dry season | 土壤水 | 0-40 | -4.41±0.03 | -41.74±1.45 | -6.15±0.19 | -46.58±0.35 | -4.55±0.25 | -47.72±0.77 | ||
| 40-100 | -7.47±0.18 | -56.82±0.74 | -9.06±0.10 | -62.53±1.33 | -8.15±0.14 | -59.58±0.97 | ||||
| 100-200 | -9.14±0.27 | -68.40±1.26 | -12.64±0.36 | -73.70±0.39 | -9.45±0.26 | -68.92±1.04 | ||||
| 湿润季 | 降水 | -6.38 | -38.78 | -6.38 | -38.78 | -6.38 | -38.78 | |||
| Wet season | 土壤水 | 0-40 | -7.74±0.06 | -60.74±0.69 | -9.53±0.40 | -58.89±0.50 | -7.69±0.67 | -55.11±3.24 | ||
| 40-100 | -8.75±0.27 | -68.66±0.11 | -10.49±0.24 | -63.72±0.35 | -8.82±0.42 | -62.29±2.44 | ||||
| 100-200 | -10.49±0.39 | -76.62±0.25 | -12.98±0.59 | -69.42±1.01 | -9.18±0.05 | -69.12±0.40 | ||||
| 水分条件 Moisture conditions |
水源 Source |
土层深度 Soil depth/cm |
HrPt | HrRp | ||||||
| δ18O/‰ | δD/‰ | δ18O/‰ | δD/‰ | |||||||
| 旱季 | 降水 | -3.69 | -24.70 | -3.69 | -24.70 | |||||
| Dry season | 土壤水 | 0-40 | -3.72±0.20 | -43.29±1.47 | -6.17±0.58 | -55.79±1.94 | ||||
| 40-100 | -8.54±0.33 | -57.82±0.73 | -9.07±0.34 | -58.12±2.50 | ||||||
| 100-200 | -10.18±0.50 | -71.98±1.05 | -10.31±0.28 | -70.97±1.33 | ||||||
| 湿润季 | 降水 | -6.38 | -38.78 | -6.38 | -38.78 | |||||
| Wet season | 土壤水 | 0-40 | -9.20±0.29 | -67.17±2.23 | -9.09±0.04 | -60.73±0.28 | ||||
| 40-100 | -9.56±0.35 | -67.17±2.23 | -10.26±0.22 | -73.62±3.10 | ||||||
| 100-200 | -10.29±0.53 | -74.16±1.32 | -11.29±0.24 | -79.28±0.40 | ||||||
旱季, 除Rp和HrRp(刺槐)茎秆水δ18O和δD值外, 其余树种纯林和混交林之间差异显著(P < 0.05)。各树种茎秆水旱季的δ18O和δD值均小于同时期降水值, 表明干旱时期植物很少利用同时期降水。湿润季, 各树种茎秆水δ18O和δD值在纯林和混交林之间无明显差异(P > 0.05), 且同位素较干旱时期贫化。此外, 湿润季各树种茎秆水的同位素值均低于同时期降水值, 与7月下旬和8月降水同位素均值相近, 表明植物主要利用这一时期入渗到土壤中的混合雨水。
| 物种 Species |
旱季Dry season | 湿润季Wet season | |||
| δ18O/‰ | δD/‰ | δ18O/‰ | δD/‰ | ||
| Hr | -7.59±0.38 | -59.82±2.02 | -8.63±0.19 | -68.56±0.67 | |
| Pt | -6.79±0.06 | -52.91±0.70 | -9.62±0.29 | -68.25±1.19 | |
| Rp | -7.69±0.23 | -62.71±1.53 | -7.99±0.12 | -65.30±1.48 | |
| HrPt (沙棘) | -9.12±0.19 | -64.31±0.87 | -9.65±0.53 | -69.41±2.90 | |
| HrPt (油松) | -8.65±0.22 | -63.34±1.07 | -8.80±0.76 | -62.76±4.25 | |
| HrRp(沙棘) | -9.52±0.10 | -61.37±0.34 | -9.62±0.20 | -66.22±2.00 | |
| HrRp(刺槐) | -7.35±0.13 | -63.81±1.46 | -9.50±0.24 | -68.37±1.62 | |
| HrPt(沙棘):沙棘+油松混交林中的沙棘;HrPt(油松):沙棘+油松混交林中的油松;HrRp(沙棘):沙棘+刺槐混交林中的沙棘;HrRp(刺槐):沙棘+刺槐混交林中的刺槐 | |||||
运用IsoSource模型量化各树种对不同土层土壤水的利用比例(图 4和图 5)。旱季, 不同土层土壤水对沙棘和油松的贡献率有所差异(图 4)。其中, Hr利用的水分83.8%来自40 cm以下, HrPt(沙棘)92%来自40 cm以下且主要依赖100-200 cm土层;Pt利用的水分95.1%来自100 cm以上, HrPt(油松)88.4%来自40 cm以下且主要依赖于40-100 cm土层;湿润季, Hr、Pt、HrPt(沙棘)和HrPt(油松)分别有83.9%、73.5%、98.7%和85.7%来自100 cm以上。

|
| 图 4 基于IsoSource模型的沙棘和油松在旱季和湿润季对不同土层土壤水的利用比例 Fig. 4 Proportions of water uptake from different soil layers based on IsoSource for H. rhamnoides and P. tabuliformis during dry and wet seasons |

|
| 图 5 基于IsoSource模型的沙棘和刺槐在旱季和湿润季对不同土层土壤水的利用比例 Fig. 5 Proportions of water uptake from different soil layers based on IsoSource for H. rhamnoides and R. pseudoacacia during dry and wet seasons |
旱季, 不同土层土壤水对沙棘和刺槐的贡献率有所差异(图 5)。其中, Hr利用的水分83.8%来自40 cm以下, HrRp(沙棘)90.7%来自40 cm以下且主要依赖100-200 cm土层;Rp利用的水分75.8%来自40 cm以下, HrRp(刺槐)85.6%来自40 cm以下且主要依赖于100-200 cm土层;湿润季, Hr、Rp、HrRp(沙棘)和HrRp(刺槐)分别有83.9%、89.9%、91.8%和86.9%来自100 cm以上。
根据表 2、3得出, 不同树种均主要利用土壤水, 结合图 4和图 5, 旱季HrPt和HrRp中不同树种主要依赖于40 cm以下土层土壤水, 为了进一步阐明季节性干旱对混交林共生树种水分来源的影响, 通过PS指数分析了其对土壤水的竞争关系(表 4)。旱季PS指数均显著高于湿润季, 且不同时期HrRp的PS指数均高于HrPt。
| 水分条件 Moisture conditions |
相似比例指数Proportional similarity index | |
| HrPt | HrRp | |
| 旱季Dry season | 0.77 | 0.88 |
| 湿润季Wet season | 0.56 | 0.61 |
0-100 cm为各树种根系的主要分布土层(图 6)。其中, Hr根系主要分布在40-70 cm, Pt主要分布在0-10 cm和30-70 cm, HrPt(沙棘)和HrPt(油松)根系分布随深度变化趋势基本一致, 表现为在0-20 cm、30-40 cm及50-60 cm土层分布比较密集。此外, 同一土层深度, 沙棘细根表面积总体小于油松。

|
| 图 6 各树种根系分布特征 Fig. 6 The fine root distribution characteristics of each tree species |
Rp根系主要分布在0-60 cm土层(图 6), HrRp(沙棘)和HrRp(刺槐)根系分布随深度变化趋势基本一致, 表现为在0-60 cm土层分布比较密集。此外, 除Rp在80 cm土层以下, 在相同土层深度, 沙棘细根表面积总体均小于刺槐。
3 讨论 3.1 旱季不同植物水分来源差异对比分析各样地不同土层土壤水δ18O和δD值, 其均随深度的增加而减小, 在0-40 cm土层达到最大值, 土壤水分受大气蒸发的影响最为强烈, 重同位素明显富集。40-100 cm土壤水主要受降水和原有土壤水的混合影响, 同位素相对稳定。深层(100-200 cm)土壤水分主要来自雨量大、低δ18O(δD)值的降水和原有土壤水分混合, 因此同位素分馏效应相对较弱[19]。植物主要通过活性根吸收水分, 受温带半干旱气候的影响, 研究区季节性干旱频发, 降水是该地区植物水分最初来源[20]。旱季降水偏少, 表层土壤蒸发强烈, 浅层的土壤水分不断减少, 植物只能依赖更深层根系从土壤中吸收水分[21]。本研究中, 油松根系主要分布在0-70 cm土层, 侧根不发达, 因此, Pt和HrPt(油松)主要利用0-40 cm和40-100 cm土层土壤水, 利用率分别为83.9%和47.4%。刺槐根系主要分布在0-60 cm土层, Rp和HrRp(刺槐)主要利用40-100 cm和100-200 cm土层土壤水, 利用率分别为43.8%和65.3%。沙棘根系主要分布于0-60 cm土层, Hr、HrPt(沙棘)和HrRp(沙棘)主要利用40-100 cm、100-200 cm和100-200 cm土层土壤水, 利用率分别为46.8%、58.8%和58.2%。此外, 对于同一土层深度, 油松和刺槐细根表面积整体大于沙棘。但研究表明, 油松和刺槐根系均属于直根系, 即水平根系不发达[22-23], 而沙棘水平根系发达, 在林间通常是基株、分株间紧密相连, 组成株间根系网, 当遮阴或水分营养匮乏时, 其匍匐水平根的长度和分枝数会迅速增多和增长[24]。不同树种根系分布差异在一定程度上决定了其水分利用特点。这也是干旱、半干旱地区植物为了生存而采用的水分利用策略[25], 即植物会通过自我调节最大限度地利用不同水源, 从而向着最优表现型发展[26]。
3.2 湿润季不同植物水分来源差异湿润季, 不同树种对各层次土壤水分的利用发生了明显的变化, 主要表现为减少了对深层(100-200 cm)土壤水分的利用, 更倾向于利用0-100 cm土壤水分, 利用率均在70%以上。一方面是由于湿润季雨水对土壤水的补给作用, 使其土壤水分有效性增加。植物通过浅层根系吸收水分时消耗的能量相对较小, 因此, 当浅层土壤水分充足时, 植物优先利用储存在这里的水分, 而不是消耗更多的能量利用更深土层土壤水[27]。另一方面由于湿润季逐渐进入生长末期, 浅层土壤中积累了一定量的由枯落物形成的有机质, 水分供给充足时, 活性根的数量显著增加, 植物优先利用浅层水分[28]。
由旱季到湿润季, 不同植物主要水分来源由深变浅, 这主要和其根系系统有关, 即植物水平根系和垂直根系在不同阶段, 可能会发挥不同的作用[29]。孙守家等[30]对太行山栓皮栎(Quercus variabilis)的研究也表明, 在降水少、浅层土壤含水量低的旱季植物更倾向于利用深层土壤水以适应干旱环境, 在降水多、土壤含水量高的湿润季, 植物根系会减少对深层水源的依赖, 优先利用浅层土壤水。朱雅娟等[31]的研究发现, 乌兰布和沙漠的柠条(Caragana korshinskii)在不同季节分别利用浅层和较深层的土壤水, 推测其可能具有根系的二态性。
3.3 混交林共生植物水分利用关系植物对资源的竞争是一种普遍存在的现象, 多数生态系统的竞争主要发生在地下根系对水分或养分竞争[32]。人工林生态系统中吸收层位相似的植物间存在对水源的竞争现象[33]。本研究中, 在旱季, 混交林中HrRp(沙棘)和HrRp(刺槐)吸水层位相似, 均主要利用100-200 cm土层土壤水, 利用率分别为58.2%和65.3%。PS指数是表征植物间生态位重叠的指标, 可以进一步量化共生植物间水分是否存在竞争关系[18]。结果表明, 旱季PS指数显著大于湿润季, 且HrRp的PS指数最大, 因此, 在旱季, 沙棘和刺槐混交存在对水分利用的竞争现象。此外, 共生植物之间还可通过水分利用在时间或空间上的错位即生态位分离来应对干旱[13, 34]。本研究中, 旱季, HrPt(沙棘)更倾向于利用100-200 cm土层土壤水, 利用率为58.8%;而HrPt(油松)主要倾向于40-100 cm土层土壤水, 利用率为47.4%。表明沙棘与油松混交主要通过水分利用空间上的差异应对干旱环境。
黄土丘陵区属于季节性干旱事件多发地, 研究混交林树种在不同水分条件下水分来源差异, 对理解土壤-植被交互作用与物种共存具有重要作用。本试验着重分析了生长季内典型时期各树种水分利用变化, 因此只选取了生长季相对干旱和湿润两个有代表性的时段进行分析。而植物也可通过生理调节应对水分来源的变化, 后续研究还需基于更多水分梯度或结合降水事件和植物生理特征的观测, 才能全面评价干旱和半干旱地区植物水分利用的稳定和可持续性。
4 结论(1) 研究区大气降水线为δD=6.30×δ 18O-4.97(R2=0.902, P < 0.01), 与全球大气降水线相比, 斜率和截距均偏小, 说明该区域受到干旱的影响, 降水不足且蒸发强烈, 重同位素富集。
(2) 旱季纯林各树种主要水分来源集中于0-100 cm(利用率 > 63%);混交林中为40-200 cm(利用率 > 75%), 其中HrPt(沙棘)对100-200 cm为58.8%, HrPt(油松)对40-100 cm为47.4%;HrRp(沙棘)和HrRp(刺槐)对100-200 cm为58.2%和65.3%。表明沙棘与油松混交主要通过水分利用空间上的差异应对干旱。湿润季不同树种主要利用0-100 cm土层土壤水(利用率 > 73%)。
(3) 旱季PS指数显著大于湿润季, 且HrRp的PS指数最大, 表明沙棘和刺槐混交旱季存在对水分利用的竞争现象。
| [1] |
Chen D Y, Wang Y K, Liu S Y, Wei X G, Wang X. Response of relative sap flow to meteorological factors under different soil moisture conditions in rainfed jujube (Ziziphus jujuba Mill.) plantations in semiarid Northwest China. Agricultural Water Management, 2014, 136: 23-33. DOI:10.1016/j.agwat.2014.01.001 |
| [2] |
Deng L, Yan W M, Zhang Y W, Shangguan Z P. Severe depletion of soil moisture following land-use changes for ecological restoration: Evidence from northern China. Forest Ecology and Management, 2016, 366: 1-10. DOI:10.1016/j.foreco.2016.01.026 |
| [3] |
白岗栓, 侯喜录, 张占雄. 油松-沙棘混交模式对生境和油松生长的影响. 林业科学, 2006, 42(8): 37-43. |
| [4] |
Zhou Z C, Shangguan Z P. Vertical distribution of fine roots in relation to soil factors in Pinus tabulaeformis Carr. forest of the Loess Plateau of China. Plant and Soil, 2007, 291(1/2): 119-129. |
| [5] |
Wu X, Tang Y K, Chen Y M, Wen J, Xie Y L, Lu S B. Sap flow characteristics and responses to summer rainfall for Pinus tabulaeformis and Hippophae rhamnoides in the Loess hilly region of China. Ecology and Evolution, 2018, 8(1): 617-630. DOI:10.1002/ece3.3639 |
| [6] |
West A G, Hultine K R, Burtch K G, Ehleringer J R. Seasonal variations in moisture use in a piñon-juniper woodland. Oecologia, 2007, 153(4): 787-798. DOI:10.1007/s00442-007-0777-0 |
| [7] |
巩国丽, 陈辉, 段德玉. 利用稳定氢氧同位素定量区分白刺水分来源的方法比较. 生态学报, 2011, 31(24): 7533-7541. |
| [8] |
Renninger H J, Carlo N, Clark K L, Schäfer K V R. Physiological strategies of co-occurring oaks in a water- and nutrient-limited ecosystem. Tree Physiology, 2014, 34(2): 159-173. DOI:10.1093/treephys/tpt122 |
| [9] |
刘自强, 余新晓, 娄源海, 李瀚之, 贾国栋, 路伟伟. 北京山区侧柏水分利用策略. 生态学报, 2017, 37(11): 3697-3705. |
| [10] |
Li S G, Romero-Saltos H, Tsujimura M, Sugimoto A, Sasaki L, Davaa G, Oyunbaatar D. Plant water sources in the cold semiarid ecosystem of the upper Kherlen River catchment in Mongolia: a stable isotope approach. Journal of Hydrology, 2007, 333(1): 109-117. DOI:10.1016/j.jhydrol.2006.07.020 |
| [11] |
West A G, Dawson T E, February E C, Midgley G F, Bond W J, Aston T L. Diverse functional responses to drought in a Mediterranean-type shrubland in South Africa. New Phytologist, 2012, 195(2): 396-407. DOI:10.1111/j.1469-8137.2012.04170.x |
| [12] |
Wang J, Fu B J, Lu N, Zhang L. Seasonal variation in water uptake patterns of three plant species based on stable isotopes in the semi-arid Loess Plateau. Science of the Total Environment, 2017, 609: 27-37. DOI:10.1016/j.scitotenv.2017.07.133 |
| [13] |
Neumann R B, Cardon Z G. The magnitude of hydraulic redistribution by plant roots: a review and synthesis of empirical and modeling studies. New Phytologist, 2012, 194(2): 337-352. DOI:10.1111/j.1469-8137.2012.04088.x |
| [14] |
West A G, Goldsmith G R, Brooks P D, Dawson T E. Discrepancies between isotope ratio infrared spectroscopy and isotope ratio mass spectrometry for the stable isotope analysis of plant and soil waters. Rapid Communications in Mass Spectrometry, 2010, 24(14): 1948-1954. DOI:10.1002/rcm.4597 |
| [15] |
林光辉. 稳定同位素生态学: 先进技术推动的生态学新分支. 植物生态学报, 2010, 34(2): 119-122. DOI:10.3773/j.issn.1005-264x.2010.02.001 |
| [16] |
Ellsworth P Z, Williams D G. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant and Soil, 2007, 291(1/2): 93-107. |
| [17] |
Wang J, Lu N, Fu B J. Inter-comparison of stable isotope mixing models for determining plant water source partitioning. Science of the Total Environment, 2019, 666: 685-693. DOI:10.1016/j.scitotenv.2019.02.262 |
| [18] |
Trogisch S, Salmon Y, He J S, Hector A, Scherer-Lorenzen M. Spatio-temporal water uptake patterns of tree saplings are not altered by interspecific interaction in the early stage of a subtropical forest. Forest Ecology and Management, 2016, 367: 52-61. DOI:10.1016/j.foreco.2016.02.018 |
| [19] |
王玉阳, 陈亚鹏, 李卫红, 王日照, 周莹莹, 张建鹏. 塔里木河下游典型荒漠河岸植物水分来源. 中国沙漠, 2017, 37(6): 1150-1157. |
| [20] |
Ohte N, Koba K, Yoshikawa K, Sugimoto A, Matsuo N, Kabeya N, Wang L H. Water utilization of natural and planted trees in the semiarid desert of Inner Mongolia, China. Ecological Applications, 2003, 13(2): 337-351. DOI:10.1890/1051-0761(2003)013[0337:WUONAP]2.0.CO;2 |
| [21] |
Ehleringer J R, Dawson T E. Water uptake by plants: perspectives from stable isotope composition. Plant, Cell & Environment, 1992, 15(9): 1073-1082. |
| [22] |
王进鑫, 罗伟祥, 刘广全, 王迪海. 黄土高原人工林根区土壤水分亏缺状况与空间分布. 西北林学院学报, 2004, 19(4): 1-4, 8-8. DOI:10.3969/j.issn.1001-7461.2004.04.001 |
| [23] |
吕春娟, 陈丽华, 周硕, 宋恒川, 盖小刚, 冀晓东, 张心平. 油松根系固土的基本力学特性. 水土保持学报, 2011, 25(5): 17-20, 25-25. |
| [24] |
王岚, 张小民, 林美珍, 张吉科. 论沙棘根系与功能Ⅰ——根系分化与分工. 沙棘, 2005, 18(4): 13-19. |
| [25] |
Wang L, Mu Y, Zhang Q F, Zhang X C. Groundwater use by plants in a semi-arid coal-mining area at the Mu Us Desert frontier. Environmental Earth Sciences, 2013, 69(3): 1015-1024. DOI:10.1007/s12665-012-2023-2 |
| [26] |
周海, 郑新军, 唐立松, 李彦. 准噶尔盆地东南缘多枝柽柳、白刺和红砂水分来源的异同. 植物生态学报, 2013, 37(7): 665-673. |
| [27] |
段德玉, 欧阳华. 稳定氢氧同位素在定量区分植物水分利用来源中的应用. 生态环境, 2007, 16(2): 655-660. DOI:10.3969/j.issn.1674-5906.2007.02.076 |
| [28] |
Sun S J, Meng P, Zhang J S, Wan X C. Variation in soil water uptake and its effect on plant water status in Juglans regia L. during dry and wet seasons. Tree Physiology, 2011, 31(12): 1378-1389. DOI:10.1093/treephys/tpr116 |
| [29] |
Williams D G, Ehleringer J R. Intra- and inter specific variation for summer precipitation use in Pinyon-Juniper woodlands. Ecological Monographs, 2000, 70(4): 517-537. |
| [30] |
孙守家, 孟平, 张劲松, 贾长荣, 任迎丰. 太行山南麓山区栓皮栎-扁担杆生态系统水分利用策略. 生态学报, 2014, 34(21): 6317-6325. |
| [31] |
朱雅娟, 贾志清, 卢琦, 郝玉光, 张景波, 李磊, 綦艳林. 乌兰布和沙漠5种灌木的水分利用策略. 林业科学, 2010, 46(4): 15-21. DOI:10.3969/j.issn.1672-8246.2010.04.003 |
| [32] |
张彦东, 沈有信, 白尚斌, 王政权. 混交条件下水曲柳落叶松根系的生长与分布. 林业科学, 2001, 37(5): 18-23. |
| [33] |
曾巧, 马剑英. 黑河流域不同生境植物水分来源及环境指示意义. 冰川冻土, 2013, 35(1): 148-155. |
| [34] |
吴旭. 黄土丘陵区沙棘与四个混交树种水分养分利用特征研究[D]. 杨凌: 中国科学院水利部水土保持研究所(中国科学院教育部水土保持与生态环境研究中心), 2019.
|