 2022, Vol. 42
2022, Vol. 42文章信息
- 赵芳, 张明伟, 王春雯, 梅续芳, 解李娜, 马成仓
- ZHAO Fang, ZHANG Mingwei, WANG Chunwen, MEI Xufang, XIE Lina, MA Chengcang
- 锦鸡儿属灌丛化对草原化荒漠区土壤线虫群落组成和代谢足迹的影响
- Effects of Caragana shrub encroachment on soil nematode community composition and metabolic footprints in steppe desert region
- 生态学报. 2022, 42(10): 4124-4136
- Acta Ecologica Sinica. 2022, 42(10): 4124-4136
- http://dx.doi.org/10.5846/stxb202106081517
-
文章历史
- 收稿日期: 2021-06-08
- 网络出版日期: 2022-01-11

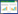




灌丛化是指木本植物的密度、盖度和生物量增加的现象。近年来, 由于全球气候变化和过度放牧等因素的影响, 全球草原区灌丛化现象越来越普遍[1], 而且灌丛扩张的速度逐年递增。内蒙古草原占我国国土面积的八分之一, 是我国北方重要的畜牧产区和生态屏障。但是, 以锦鸡儿属(Caragana)灌木为主的灌丛扩张日益成为该地区的普遍现象[2]。灌丛化显著提高灌丛内土壤含水量[3]和土壤肥力, 从而对林下植物物种丰富度、生物量和物种多样性[4]产生正效应。灌丛化对地下生物群落(包括土壤微生物群落和土壤动物群落)也有显著影响[5-6]。
土壤线虫是土壤动物的重要组成部分, 其食性多样, 分布在土壤食物网中多个营养级, 是有机质分解和养分循环等重要土壤过程的关键驱动因子, 很容易受到灌木入侵的影响[7]。目前关于灌木对土壤线虫群落的研究主要集中在对其多样性和功能指数的影响[8], 尤其是物种多样性。事实上, 生物多样性包括物种多样性、功能多样性和系统发育多样性三个维度。与物种多样性相比, 功能多样性和系统发育多样性是近年来才被引入线虫生态学研究中[9-10]。因此, 关于灌木对土壤线虫群落功能多样性和系统发育多样性的影响依然知之甚少。
多样性指数和功能指数被认为是描述土壤食物网结构和功能、评价土壤健康状况的有效指标, 但它们是建立在各类线虫相对比例的基础上, 没有考虑到土壤线虫的绝对数量, 从而导致数量不同的土壤线虫群落可能具有相同的土壤食物网结构[11]。为此, Ferris在2010年提出用线虫代谢足迹来反映线虫的碳代谢过程, 即以线虫生物量碳和呼吸碳含量度量线虫的代谢活性和土壤食物网功能。线虫代谢足迹为估算线虫对生态系统功能的贡献提供了有效的方法[12]。近年来, 有关线虫代谢足迹的研究开始受到关注。但目前主要在群落水平, 如:研究植被演替过程[13-14]和灌丛化过程[15-16]中土壤线虫代谢足迹的变化, 从灌丛局域水平上的研究较少。事实上, 研究不同种灌木的灌丛个体对线虫代谢足迹的影响更能揭示灌丛影响土壤线虫群落的机制。
土壤理化性质(湿度、pH、有机质等)和植物群落特征(植物种类、多样性、生物量等)被认为是影响土壤线虫群落组成的重要因素[17]。目前的研究主要基于土壤线虫丰度, 从营养类群、cp类群或种属水平上探讨影响土壤线虫组成和结构的环境因素。研究表明:植物寄生线虫、食微线虫和捕食杂食线虫的丰富度随植物种类和功能类群多样性的增加而增加[18];食细菌线虫和食真菌线虫相对丰度与土壤有效磷含量相关, 植物寄生线虫和捕/杂食线虫相对丰度与有机碳、铵态氮和全氮含量相关[19]。线虫功能团将其生活史策略和食性相结合, 可以更好地反映土壤食物网对外界环境的响应[20]。因此, 从功能团水平探究影响群落组成的环境因子能更好的揭示灌丛影响土壤食物网的机制。
土壤线虫群落组成(营养类群、cp类群和特殊种属)以及对环境的响应与地上植物种有关。例如西藏北部高寒草甸3种典型植物群落土壤线虫优势种存在明显差异, 委陵菜群落的优势属为矮化属(Tylenchorhynchus)和螺旋属(Helicotylenchus), 高山嵩草群落的优势属为垫咽属(Tylencholaimus)和矮化属(Tylenchorhynchus), 而藏北嵩草群落的优势属为鹿角唇属(Cervidellus)和丝尾垫刃属(Filenchus), 藏北嵩草群落cp3类群的个体数量及相对多度均高于高山嵩草群落, 而其它cp类群则低于高山嵩草群落[21];固氮树种(桤木)的食细菌线虫、植物寄生线虫、杂食线虫的丰度均显著高于非固氮树种(大桉)[22];干旱显著增加了花椒和紫花苜蓿混合培养中植物寄生类、食细菌类和食真菌类线虫的多度, 但显著降低了花椒和大豆混合培养中食细菌类和食真菌类线虫的多度[23]。因此, 为了更好的理解灌丛对土壤线虫群落组成的影响, 弄清营养类群、cp类群以及特殊种属线虫在灌丛内外及灌木种间的差异及其影响机制至关重要。
基于以上背景, 本研究在内蒙古草原化荒漠区选择3种锦鸡儿属灌木-垫状锦鸡儿(Caragana tibetica)、荒漠锦鸡儿(C. roborovskyi)和狭叶锦鸡儿(C. stenophylla), 调查了灌丛内、外的土壤线虫群落多样性、组成和代谢足迹, 以及相关的土壤理化性质和植物群落特征, 探讨锦鸡儿属灌丛对土壤线虫群落的影响, 同时在线虫功能团水平上分析灌丛土壤线虫群落组成与非生物因素和生物因素的关系。试图回答以下科学问题:(1)灌丛如何影响土壤线虫群落多样性、组成和结构?(2)这种影响是否具有灌木种间差异?(3)灌丛如何通过非生物因素和生物因素影响土壤线虫群落组成?
1 材料与方法 1.1 研究区概况研究地点位于内蒙古自治区鄂尔多斯市西部鄂托克旗棋盘井镇(38°17′ N, 106°42′ E, 海拔1800 m)。该地区属温带草原化荒漠区, 典型大陆型气候, 以风沙大、干旱少雨、日照充足、蒸发强烈为主要特点。年均降雨量222 mm, 主要集中在4-9月;年均蒸发量2359 mm, 是降水量的8倍以上;年日照时间3000 h左右, 年均气温7.0℃, 年均无霜期129 d。该地区植被稀疏单一, 主要优势灌木为垫状锦鸡儿(C. tibetica)、荒漠锦鸡儿(C. roborovskyi)和狭叶锦鸡儿(C. stenophylla);主要草本植物有沙生针茅(Stipa glareosa)、无芒隐子草(Cleistogenes songorica)和刺沙蓬(Salsola ruthenica)。
1.2 研究方法 1.2.1 样品采集于2016年7月在研究地点选取植被生长较好的灌丛化草地作为实验样地。在样地中建立一个分布有垫状锦鸡儿、荒漠锦鸡儿和狭叶锦鸡儿灌丛的样方(100 m×100 m), 样方内选取立地条件基本一致的垫状锦鸡儿、荒漠锦鸡儿和狭叶锦鸡儿灌丛各3株作为样株, 在每个样株内设置50 cm×50 cm小样方, 小样方位于灌丛下的中心位置(灌丛内), 同时在样方内均匀选择3个点, 设置同样大小的小样方(灌丛外), 并确保灌丛外小样方距离灌丛边缘2 m以上。对每个小样方, 首先剪取各灌丛样株地上部, 然后调查小样方内草本植物种类和多度并收获其地上部分, 带回实验室于65℃烘干至恒重, 测定锦鸡儿属灌丛和草本植物生物量。同时, 分别采集样方内0-10 cm和10-20 cm土层的土壤样品;每层土样混合均匀后取300克放入自封袋, 4℃保存迅速带回实验室。每份土壤样品一部分立刻用于土壤线虫的分离鉴定, 一部分用于土壤理化性质的测定。
1.2.2 土壤线虫分离与鉴定土壤线虫的分离提取采用改良的Baermann法[24]。从每份土壤样品中取20 g鲜土分离土壤线虫, 分离时间为24 h, 用TAF固定液(三乙醇胺-福尔马林固定液)固定保存线虫。根据《中国土壤动物检索图鉴》[25]、《植物线虫分类学》[26]和《De Nematoden van Nederland》[27]在光学显微镜下鉴定到属, 并记录每个属线虫数。鉴定中, 如果样品中的土壤线虫数低于100条, 则全部鉴定;如果多于100条, 则随机抽取其中的100条进行鉴定。将每个小样方内0-10 cm和10-20 cm两个土样统计鉴定结果合并表示0-20 cm的线虫数量特征, 物种丰富度用上述40克鲜土样的物种数表示, 线虫多度用每100 g干土(用土壤含水量换算)中土壤线虫的个体数表示。
1.2.3 土壤理化性质测定土壤含水量采用烘干法;土壤有机质采用重铬酸钾加热法;硝态氮和铵态氮采用流动分析仪法;速效磷采用碳酸氢钠浸提-钼锑抗比色法;速效钾采用醋酸铵浸提-火焰光度法;pH值采用水浸提电位法(水土比为2.5 ∶ 1);电导率采用水土5 ∶ 1浸提后DDS-LLa型电导仪测量[28]。用每个小样方内0-10 cm和10-20 cm两个土样平均值表示0-20 cm的土壤理化性质。
1.3 数据处理 1.3.1 土壤线虫生态指数(1) 多样性指数:采用物种多样性、功能多样性和系统发育多样性3个维度分析土壤线虫群落的多样性。
物种多样性用土壤线虫多度(总个体数)、丰富度(物种数)和Shannon-Wiener指数表示。

|
式中, Pi表示群落第i个物种的个数占生物总数的比例。
功能多样性用功能丰富度(FRic)表示。基于3种性状将土壤线虫划分成不同的功能类群:①营养类群:根据土壤线虫的食性特征、食道的结构和捕食方法等, Yeates等[29]将线虫划分为8个取食类群, 其中食细菌类群、食真菌类群、植物寄生类群和捕食杂食类群是土壤生态系统中4个主要营养类群;②cp类群:根据土壤线虫生活史策略将线虫划分为r-对策向K-对策过渡的5个cp(colonizer-persister)类群, cp1到cp5, 其中cp1类群为典型r-对策者, cp 5类群为典型K-对策者[30-31];③线虫个体生物量(μg):特定属中所有种的平均生物量。以上线虫性状特征(食性、cp, 体重)均从Nematode Ecophysiological Parameter (http://nemaplex.ucdavis.edu)数据库中获得。功能多样性采用R软件“FD”程序包计算。
系统发育多样性是基于本研究鉴定的所有线虫分类系统, 利用Open Tree of Life (OTL)建立系统发育树, 然后量化群落系统发育总枝长, 即系统发育多样性指数。系统发育多样性采用R软件“rotl”、“ape”和“picante”程序包计算[32]。
(2) 土壤线虫成熟度指数:采用自由生活线虫成熟度指数(MI)和植物寄生线虫成熟度指数(PPI)评价土壤线虫对外界扰动的响应。
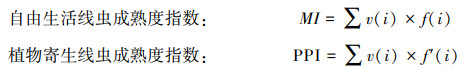
|
式中, v(i)为线虫的生活史cp值;f(i)为第i类自由生活线虫(不包括植食性线虫)占自由生活线虫总数的比例;f′(i)为第i类植物寄生线虫占植物寄生类线虫总数的比例[29-30]。
1.3.2 土壤线虫组成采用土壤线虫各营养类群、cp类群和特殊属多度反映线虫群落组成特征, 并用非度量尺度分析法(Non-metric multidimensional scaling, NMDS;基于Bray-Curtis相异系数)将其组成可视化, 比较群落组成的相似性;NMDS采用PAST3软件进行。
1.3.3 土壤线虫代谢足迹利用“线虫-植物专家信息系统”(http://plpnemweb.ucdavis.edu/nemaplex)中已经列出的各类群线虫的生物量(鲜重, W)计算线虫代谢足迹。
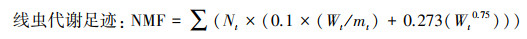
|
式中, Nt、Wt和mt分别表示t类线虫个体数量、鲜重和cp值。
根据线虫的营养类群计算食细菌线虫代谢足迹、食真菌线虫代谢足迹、植物寄生线虫代谢足迹和捕食杂食线虫代谢足迹, 其中前三者指示通过细菌通道、真菌通道和植物通道进入土壤食物网的碳和能量, 而捕食杂食线虫代谢足迹指示进入较高营养级的捕食杂食线虫类群的碳和能量。
根据线虫的生活史策略计算富集代谢足迹、结构代谢足迹和功能代谢足迹。富集代谢足迹和结构代谢足迹分别反映较低营养级线虫和较高营养级线虫的碳代谢过程[12]。功能代谢足迹是富集代谢足迹和结构代谢足迹的总面积, 即(Fs×Fe)/2, 反映土壤食物网调控和维持代谢平衡的能力。功能足迹越大, 反映线虫对土壤食物网的调控能力越强, 被捕食者与捕食者之间维持代谢平衡的能力越强[12]。
1.3.4 土壤线虫区系分析计算线虫群落的富集指数(EI)和结构指数(SI), 并结合EI和SI指数进行线虫区系分析, 评估土壤食物网的结构与功能变化, 从而评价土壤食物网对外界干扰的响应和恢复状况。

|
式中, b代表食物网中的基础成分, 主要指cp值为2的食细菌线虫和食真菌线虫;e代表食物网中的富集成分, 主要指cp值为1的食细菌线虫和cp值为2的食真菌线虫;s代表食物网中的结构成分, 包括cp3-5的食细菌线虫、食真菌线虫和捕食杂食线虫[31]。
根据计算出的Fe和Fs, 在A、B、C、D 4个象限中以菱形图的方式反映土壤食物网状况。菱形的中心点坐标为(SI, EI), 4个顶点的坐标分别为(SI-0.5Fs/k, EI)、(SI+0.5Fs/k, EI)、(SI, EI-0.5Fe/k)和(SI, EI+0.5Fe/k)。其中k为转换常数, 乘以0.5是为了使Fs和Fe能围绕中心坐标点(SI, EI)对称分布。依次连接4个坐标点, 所圈定的菱形区域的面积为线虫的功能代谢足迹。菱形横轴和纵轴方向对角线的长度可以反映Fs和Fe的相对大小, 若菱形横轴对角线的尺度大于纵轴对角线的尺度, 说明Fs大于Fe;反之亦然。当线虫区系分布在A象限时, 表明土壤环境受干扰程度较高;在B象限时, 表明土壤养分状况较好而且受干扰程度较小, 食物网稳定成熟;在C象限时, 表明土壤受干扰程度较小, 食物网处于结构化状态;在D象限时, 表明土壤受干扰程度最高, 食物网退化[31]。
1.3.5 土壤线虫物种选择性以物种对生境的特异性和适宜性为基础, 采用物种指示值分析法[33]计算土壤线虫的指示值(Indicator value;IndVal), 并通过IndVal值确定灌丛选择性物种(物种通常仅生长在灌丛内或灌丛外)和灌丛非选择性物种(物种在灌丛内和灌丛外均可生长)。如果IndVal值大于0.6, 则认为该物种是灌丛选择性物种, 否则为灌丛非选择性物种;灌丛内IndVal值大于0.6, 为灌丛正选择物种, 灌丛外IndVal值大于0.6, 为灌丛负选择物种。然后, 计算灌丛正选择性物种和负选择性物种占总物种的百分比。物种IndVal值采用R软件中“labdsv”程序包计算。
1.4 统计分析采用IBM SPSS 19软件, 通过单因素方差分析检验锦鸡儿属灌丛对各指标影响的显著性, 并用Duncan检验做多重比较, 用独立样本t检验分析灌丛内和灌丛外的差异。采用ANOSIM检验锦鸡儿属灌丛对线虫群落组成影响的显著性, ANOSIM分析采用R软件“vegan”程序包进行。采用Mantel tests分析土壤线虫群落组成与环境因子的相关性, Mantel tests采用R软件“vegan”程序包进行。
2 结果与分析 2.1 土壤线虫总多度和各营养类群多度垫状锦鸡儿灌丛共分离土壤线虫208条/100 g干土, 17个物种, 隶属于2纲3目8科17属。优势属共3属, 包括丽突属(Acrobeles)、拟丽突属(Acrobeloides)和真滑刃属(Aphelenchus), 占土壤线虫总数的63.45%(表 1);荒漠锦鸡儿灌丛共分离土壤线虫247条/100 g干土, 21个物种, 隶属于2纲3目10科21属。优势属共3属, 包括丽突属(Acrobeles)、拟丽突属(Acrobeloides)和真滑刃属(Aphelenchus), 占土壤线虫总数的76.08%(表 1);狭叶锦鸡儿灌丛共分离土壤线虫162条/100 g干土, 17个物种, 隶属于2纲3目9科17属。优势属共3属, 包括丽突属(Acrobeles)、拟丽突属(Acrobeloides)、真矛线属(Eudorylaimus), 占土壤线虫总数的54.47%(表 1)。3种锦鸡儿属灌丛土壤线虫总多度和物种丰富度灌丛内外均无显著差异, 不同灌木种之间也无显著差异(表 1)。
| 营养类群 Trophic Groups |
属名 Genus |
cp值 cp value |
垫状锦鸡儿 C. tibetica |
荒漠锦鸡儿 C. roborovskyi |
狭叶锦鸡儿 C. stenophylla |
灌丛外 Outside shrub |
| BF多度 | 丽突属Acrobeles | 2 | 50.33±6.06abiii | 104.67±29.06aiii | 26.00±7.37biii | 42.67±7.88iii |
| Abundance of BF | 拟丽突属Acrobeloides | 2 | 26.67±0.88aiii | 32.67±20.63aiii | 36.00±9.71aiii | 27.00±6.66iii |
| 真头叶属Eucephalobus | 2 | 1.00±1.00ai | 1.00±1.00ai | 0a | 0 | |
| 板唇属Chiloplacus | 2 | 8.33±4.41aii | 0a | 2.67±1.45aii | 1.00±1.00i | |
| 威尔斯属Wilsonema | 2 | 0a | 1.00±1.00ai | 0a | 1.00±1.00i | |
| 头叶属Cephalobus | 2 | 0a | 3.67±2.33aii | 0a | 0 | |
| 鹿角唇属Cervidellus | 2 | 0a | 0a | 0a | 1.67±1.67ii | |
| 类群合计Total | 86.33±5.70a | 143±50.02a | 64.67±17.64a | 73.33±13.09 | ||
| FF多度 | 滑刃属Aphelenchoides | 2 | 8.33±4.41aii | 1.67±1.67ai | 17.00±10.54aii | 6.00±3.06ii |
| Abundance of FF | 真滑刃属Aphelenchus | 2 | 52.67±15.88aiii | 51.00±12.42aiii | 3.33±1.67bii | 13.67±8.09ii |
| 伪垫刃属Nothotylenchus | 2 | 2.00±1.00ai | 1.00±1.00ai | 5.00±5.00aii | 1.00±1.00i | |
| 丝尾垫刃属Filenchus | 2 | 1.00±1.00bi | 1.00±1.00bi | 5.33±1.45a*ii | 0 | |
| 类群合计Total | 64±12.29a* | 54.67±12.35a | 30.67±13.86a | 20.67±9.21 | ||
| PP多度 | 裸矛属Psilenchus | 2 | 0a | 1.00±1.00ai | 0a | 0 |
| Abundance of PP | 垫刃属Tylenchus | 2 | 0a | 1.67±1.67ai | 0a | 0 |
| 散香属Boleodorus | 2 | 2.67±2.67aii | 0a | 2.00±1.00aii | 1.00±1.00i | |
| 盘旋属Rotylenchus | 3 | 5.00±5.00aii | 7.00±3.79aii | 0a | 7.00±3.06ii | |
| 头垫刃属Tetylenchus | 3 | 15.33±9.60aii | 1.00±1.00ai | 13.67±8.29aii | 12.67±7.67ii | |
| 矮化属Tylenchorhynchus | 3 | 11.67±6.67aii | 3.33±1.67aii | 9.33±2.33aii | 24.33±10.59iii | |
| 螺旋属Helicotylenchus | 3 | 0a | 1.00±1.00ai | 0a | 1.67±1.67ii | |
| 短体属Pratylenchus | 3 | 0a | 2.67±1.45ai | 0a | 6.00±3.79ii | |
| 潜根属Hirschmanniella | 3 | 0a | 1.00±1.00ai | 0a | 0 | |
| 长针属Longidorus | 5 | 0a | 0a | 1.00±1.00ai | 0 | |
| 缢咽属Axonchium | 5 | 0a | 0a | 1.67±1.67aii | 0 | |
| 类群合计Total | 34.67±23.47a | 18.67±6.36a | 27.67±10.99a | 52.67±13.30 | ||
| OP多度 | 拟桑尼属Thorneella | 4 | 0a | 0a | 0a | 1.00±1.00i |
| Abundance of OP | 盘咽属Discolaimus | 4 | 5.33±3.93aii | 4.67±1.67aii | 3.33±3.33aii | 0 |
| 真矛线属Eudorylaimus | 4 | 10.67±4.26bii | 10.33±3.93bii | 25.33±4.33aiii | 23.67±14.85iii | |
| 桑尼属Thornia | 4 | 2.67±1.45bii | 12.67±3.93aii | 4.33±0.67abii | 7.00±2.08ii | |
| 拱唇属Labronema | 4 | 2.00±1.00ai | 0a | 3.67±2.33aii | 0 | |
| 孔咽属Aporcelaimus | 5 | 2.67±1.45aii | 2.67±2.67ai | 2.00±1.00aii | 1.00±1.00i | |
| 类群合计Total | 23.33±4.67a | 30.33±2.33a | 38.67±9.39a | 32.67±16.51 | ||
| 丰富度Richness | 11.67±1.76a | 12.33±1.67a | 12.67±0.33a | 11.67±0.67 | ||
| 总多度Abundance | 208.33±16.42a | 246.67±42.56a | 161.67±37.2a | 179.33±37.56 | ||
| BF:食细菌类Bacterial feeders;FF:食真菌类线虫Fungal feeders;PP:植物寄生类Plant-parasites;OP:捕食杂食类Omnivores-predators;iii:优势属, > 10%;ii:常见属, 1%-10%;i:稀有属, < 1%;0:未出现;*表示灌丛内和灌丛外差异显著(t-test, P < 0.05);不同小写字母表示3种锦鸡儿灌丛之间差异显著(Duncan tests, P < 0.05) | ||||||
从营养类群上看, 3种锦鸡儿属灌丛均表现为食细菌类线虫多度最大, 占总数的比例约为39.23%-55.31%;其次为食真菌类线虫。垫状锦鸡儿多度最小的是捕食杂食类线虫, 占总数的11.20%;荒漠锦鸡儿和狭叶锦鸡儿多度最小的是植物寄生类线虫, 占总线虫的比例分别为7.77%和16.97%。3种锦鸡儿属灌木中, 只有垫状锦鸡儿灌丛土壤食真菌类线虫多度显著大于灌丛外, 其余各营养类群在灌丛内外和种间差异均不显著(表 1中类群合计)。
2.2 土壤线虫群落组成分析在分离得到的28属土壤线虫中, 食细菌类线虫丽突属(Acrobeles)、食真菌类线虫真滑刃属(Aphelenchus)和丝尾垫刃属(Filenchus)以及捕食杂食类线虫真矛线属(Eudorylaimus)和桑尼属(Thornia)5属线虫在灌丛内外或灌木种间存在显著差异(表 1)。其中, 荒漠锦鸡儿丽突属多度最大, 且显著大于狭叶锦鸡儿;垫状锦鸡儿和荒漠锦鸡儿真滑刃属多度相差不大, 但均显著大于狭叶锦鸡儿;狭叶锦鸡儿丝尾垫刃属多度显著大于垫状锦鸡儿和荒漠锦鸡儿, 灌丛外未分离出该属线虫;狭叶锦鸡儿真矛线属多度显著大于垫状锦鸡儿和荒漠锦鸡儿;荒漠锦鸡儿桑尼属多度最大, 且显著大于垫状锦鸡儿(表 1)。
非度量多维尺度(NMDS)分析排序了不同种锦鸡儿灌丛内和灌丛外土壤线虫群落组成。土壤线虫食细菌类、食真菌类、植物寄生类、cp2类群和总线虫群落的Stress≤0.2, 说明整体上NMDS分析3种锦鸡儿属灌丛土壤线虫群落结构拟合度高(图 1)。ANOSIM检验结果显示, 锦鸡儿属灌丛显著(或接近显著)影响了食细菌类群(P=0.048)、食真菌类群(P=0.052)、cp2类群(P=0.007)和总线虫群落(P=0.007)的组成(图 1)。

|
| 图 1 3种锦鸡儿灌丛内外土壤线虫营养类群、cp类群和总线虫群落的非度量多维尺度分析(NMDS)和ANOSIM检验结果 Fig. 1 Non-metric multidimensional scale analysis (NMDS) and ANOSIM test of soil nematode trophic groups, cp groups and nematode community outside and inside three Caragana shrub canopies BF:食细菌类Bacterial feeders;FF:食真菌类线虫Fungal feeders;PP:植物寄生类Plant-parasites;OP:捕食杂食类Omnivores-predators;Community:总线虫群落; T:垫状锦鸡儿灌丛内Inside C. tibetica canopies;R:荒漠锦鸡儿灌丛内Inside C. roborovskyi canopies;S:狭叶锦鸡儿灌丛内Inside C. stenophylla canopies;N:灌丛外Outside shrub canopies;P值为ANOSIM检验结果, P < 0.05, 表示差异显著 |
3种锦鸡儿属灌丛土壤均未分离出cp1类群线虫, 且cp2类群线虫多度大于cp 3-5类群(图 2)。垫状锦鸡儿灌丛内cp2类群线虫多度显著大于灌丛外, 其余种灌丛内外均无显著差异, 也没有种间差异, cp3-5类群线虫多度在灌丛内外和不同灌木种之间均无显著差异(图 2)。

|
| 图 2 3种锦鸡儿灌丛内外土壤线虫cp2和cp3-5类群多度 Fig. 2 Abundance of nematode cp2 and cp3-5 groups outside and inside three Caragana shrub canopies *表示灌丛内和灌丛外差异显著(t-test, P < 0.05);不同小写字母表示3种锦鸡儿灌丛之间差异显著(Duncan tests, P < 0.05) |
土壤线虫多样性指数(包括Shannon-Wiener、功能丰富度指数和系统发育多样性指数)和成熟度指数(MI和PPI)在灌丛内外及灌木种间均没有显著差异(图 3, 图 4)。

|
| 图 3 3种锦鸡儿灌丛内外土壤线虫群落多样性指数 Fig. 3 Diversity index of soil nematode outside and inside three Caragana shrub canopies |

|
| 图 4 3种锦鸡儿属灌丛内外土壤线虫群落MI和PPI指数 Fig. 4 MI and PPI indexes of soil nematode outside and inside three Caragana shrub canopies MI:自由生活线虫成熟度指数Maturity index;PPI:植物寄生线虫成熟度指数Plant parasitic index |
各营养类群的代谢足迹在灌丛内外以及灌木种间均无显著差异(图 5), 但cp类群的代谢足迹具有显著差异(Fe: F3, 8= 4.95; P < 0.05)(图 6)。基于代谢足迹的区系分析显示:灌丛外土壤线虫分布在CD象限之间;狭叶锦鸡儿灌丛土壤线虫分布灌丛外右侧, C象限, 比灌丛外更靠近B象限;垫状锦鸡儿和荒漠锦鸡儿灌丛土壤线虫分布在灌丛外左侧, D象限, 比灌丛外更靠近A象限(图 6)。线虫功能足迹(菱形总面积)表现为荒漠锦鸡儿>垫状锦鸡儿>狭叶锦鸡儿和灌丛外。3种锦鸡儿属灌丛的富集指数(菱形中心位置)均高于灌丛外(图 6)。

|
| 图 5 3种锦鸡儿属灌丛内外土壤线虫代谢足迹 Fig. 5 Metabolic footprint of soil nematode outside and inside three Caragana shrub canopies BFMF:食细菌类线虫代谢足迹Metabolic footprint of bacterial feeders; FFMF:食真菌类线虫代谢足迹Metabolic footprint of fungal feeders;PPMF:植物寄生类线虫代谢足迹Metabolic footprint of plant parasitic nematodes;OPMF:捕食杂食类线虫代谢足迹Metabolic footprint of omnivores-predators nematodes |

|
| 图 6 3种锦鸡儿属灌丛内外土壤线虫区系分析(基于线虫代谢足迹) Fig. 6 Analysis of soil nematode fauna outside and inside three Caragana shrub canopies (based on nematode metabolic footprints) |
锦鸡儿属灌丛对线虫营养类群和cp类群具有选择性。3种锦鸡儿属灌丛食真菌类和cp2类线虫的正选择性物种比例均大于负选择性物种比例;但植物寄生类线虫和cp3-5恰好相反, 表现为正选择性物种比例小于负选择性物种比例(表 2)。
| 线虫类群 Nematode groups |
垫状锦鸡儿 C. tibetica |
荒漠锦鸡儿 C. roborovskyi |
狭叶锦鸡儿 C. stenophylla |
|||||
| 正选择性物种 Positive-selective |
负选择性物种 Negative-selective |
正选择性物种 Positive-selective |
负选择性物种 Negative-selective |
正选择性物种 Positive-selective |
负选择性物种 Negative-selective |
|||
| BF | 0.00 | 0.00 | 28.57 | 0.00 | 0.00 | 20.00 | ||
| FF | 25.00 | 0.00 | 25.00 | 0.00 | 50.00 | 0.00 | ||
| PP | 0.00 | 33.33 | 0.00 | 22.22 | 0.00 | 37.50 | ||
| OP | 33.33 | 33.33 | 40.00 | 20.00 | 16.67 | 16.67 | ||
| cp2 | 9.09 | 0.00 | 21.43 | 0.00 | 20.00 | 10.00 | ||
| cp3-5 | 18.18 | 36.36 | 18.18 | 27.27 | 7.69 | 30.77 | ||
| cp:殖民者-居住者Colonizer-persister;cp1-cp5:表示土壤线虫从殖民者到居住者类型过渡 | ||||||||
各营养类群线虫选择性物种在灌木种间存在明显差异。食细菌类, 荒漠锦鸡儿正选择性物种比例最大, 无负选择性物种;食真菌类, 3种锦鸡儿均只有正选择性物种, 其中狭叶锦鸡儿正选择性物种比例最大;植物寄生类, 3种锦鸡儿均只有负选择性物种, 其中垫状锦鸡儿和狭叶锦鸡儿负选择性物种比例相差不大, 荒漠锦鸡儿比例最小;捕食杂食类, 荒漠锦鸡儿正选择性物种比例最大(表 2)。丽突属(Acrobeles)和头叶属(Cephalobus)是荒漠锦鸡儿的指示物种;滑刃属(Aphelenchoides)和丝尾垫刃属(Filenchus)是狭叶锦鸡儿的指示物种;盘咽属(Discolaimus)和桑尼属(Thornia)是荒漠锦鸡儿的指示物种。
各cp类群线虫选择性物种在灌木种间也存在明显差异。cp2类群, 荒漠锦鸡儿和狭叶锦鸡儿正选择性物种比例相差不大, 但均大于垫状锦鸡儿正选择性物种比例(表 2)。cp3-5类群, 垫状锦鸡儿负选择性物种比例最大(表 2)。
2.7 生物、非生物因素与线虫群落组成的相关性分析Mantel tests是评估群落物种组成和环境之间相关性的一种检验方法。Mantel tests结果显示:土壤速效钾(P < 0.05)和土壤含水量(P < 0.01)与cp值为2的食真菌类线虫功能团组成呈显著正相关;铵态氮与cp值为2的植食线虫功能团组成呈显著正相关(P < 0.05);土壤速效磷与cp值为4的捕食杂食类线虫功能团组成呈显著正相关(P < 0.05);其它生物因素(灌丛生物量、林下植物生物量和丰富度)和非生物因素(土壤理化性质)与线虫各功能团组成无显著相关性(表 3, 表 4)。
| 因素Factors | BF2 | FF2 | PP2 | PP3 | OP4 | OP5 |
| AP | -0.0189 | -0.1186 | 0.2477 | -0.1281 | 0.2683* | -0.2198 |
| AK | 0.0367 | 0.2531* | -0.1469 | -0.1208 | 0.0467 | 0.0855 |
| pH | -0.0534 | -0.0443 | -0.0872 | 0.0260 | 0.1736 | 0.1691 |
| EC | -0.2916 | -0.0260 | 0.0267 | -0.1189 | 0.1948 | -0.1785 |
| SOC | -0.0018 | -0.0783 | 0.0486 | -0.0931 | -0.0219 | -0.1194 |
| SWC | -0.0945 | 0.4410** | -0.3039 | 0.0178 | -0.0184 | -0.0132 |
| NO3--N | -0.2206 | 0.0993 | -0.1646 | -0.0352 | -0.0187 | -0.0708 |
| NH4+-N | 0.0280 | -0.1331 | 0.3772* | 0.0070 | 0.1957 | -0.0409 |
| Sbio | -0.1026 | 0.1475 | -0.1112 | 0.0872 | 0.0288 | -0.0366 |
| Herbio | -0.1079 | 0.1478 | -0.0845 | -0.1759 | 0.0113 | 0.0166 |
| Richness | -0.2130 | 0.1903 | 0.1570 | -0.1135 | 0.1038 | -0.1791 |
| AP:速效磷Available P;AK:速效钾Available K;EC:电导率Electrical conductivity;SOC:土壤有机质Soil organic material;SWC:土壤含水量Soil water content;NO3--N:硝态氮;NH4+-N:铵态氮;Sbio:锦鸡儿生物量;Herbio:林下草本植物生物量;Richness:林下草本植物丰富度;BF、FF、PP、OP下标代表cp值; * 表示显著相关, P < 0.05; ** 表示极显著相关, P < 0.01 | ||||||
| 垫状锦鸡儿 C. tibetica |
荒漠锦鸡儿 C. roborovskyi |
狭叶锦鸡儿 C. stenophylla |
灌丛外 Outside shrub |
|
| AP/(mg/kg) | 7.57±1.82 | 4.95±0.63 | 6.53±0.22 | 5.73±0.57 |
| AK/(mg/kg) | 76.00±12.58 | 80.33±6.23 | 109.33±11.46 | 84.83±11.83 |
| pH | 8.21±0.12 | 8.52±0.06 | 8.31±0.06 | 8.72±0.23 |
| EC/(μs/cm) | 260.67±27.18 | 170.00±57.04 | 134.67±3.18 | 96.00±21.70 |
| SOC/(g/kg) | 10.57±4.60 | 10.27±1.64 | 12.66±1.86 | 3.85±1.29 |
| SWC/% | 3.84±0.43 | 3.70±0.10 | 5.18±0.63 | 2.40±0.12 |
| NO3--N/(mg/kg) | 8.64±5.01 | 5.77±2.24 | 10.46±2.34 | 1.50±0.22 |
| NH4+-N/(mg/kg) | 0.09±0.02 | 0.26±0.08 | 0.07±0.01 | 0.20±0.08 |
| Sbio/g | 142.99±10.52 | 138.14±13.76 | 90.14±5.53 | 0.00 |
| Herbio/g | 10.46±0.70 | 11.27±2.11 | 12.90±1.74 | 9.98±3.20 |
| Richness | 3.67±0.17 | 3.17±0.33 | 3.17±0.17 | 3.67±0.44 |
本文研究结果表明锦鸡儿属灌丛对土壤线虫多度、丰富度和多样性无显著影响, 但却显著影响土壤线虫群落组成。5个属(占总属的17.86%)线虫多度在灌丛内外或灌木种间出现显著差异(表 1), 绝大多数线虫营养类群和cp类群组成也表现出灌丛内外或灌木种间的显著差异(图 1)。非度量多维度尺度(NMDS)分析也显示线虫群落组成在灌丛内外或灌木种间差异很大, 进一步的ANOSIM检验证实锦鸡儿属灌丛对土壤线虫群落组成, 以及一些营养类群和cp类群的组成具有显著或接近显著的影响(图 1)。以往的研究已经发现不同的灌木对土壤线虫的组成产生不同的影响, 如, Dasiphora frutico的存在不影响土壤线虫群落多度和组成[34], 但是Piliostigma reticulatum显著影响植物寄生类线虫丰度[35]。
认为灌丛通过两种机制影响土壤线虫群落、营养类群和cp类群的组成。第一, 灌丛对土壤线虫群落具有物种选择性。(1)从群落水平上, 3种锦鸡儿灌木对不同线虫属具有偏好或排斥性(表 2), 从而导致土壤线虫群落组成的差异。(2)从营养类群水平上, 3种锦鸡儿属灌丛食真菌类线虫正选择性物种比例大于负选择性物种比例, 而植物寄生类则相反, 负选择性物种比例更大(表 2)。这说明锦鸡儿属灌丛可能更有利于食真菌类线虫定居, 尤其是滑刃属(Aphelenchoides), 这可能是因为灌丛的遮阴和土壤的改善效应提高了灌丛土壤的保湿能力[36], 而滑刃属中的某些种更适合在相对湿度较高的环境中生存[37-38]。与其它营养类群线虫相比, 植物寄生类线虫可能更适合在无灌丛区域定居(表 2)。其原因是, 与灌木相比, 灌丛外的禾草类能提供质量更高的根系资源[39], 从而对一些以植物为食的线虫有更大的吸引力[40]。另外, 对于食细菌类线虫, 荒漠锦鸡儿正选择性物种比例最大, 而狭叶锦鸡儿仅有负选择性物种(表 2)。这说明荒漠锦鸡儿可能更有利于食细菌类线虫生长, 而狭叶锦鸡儿对其具有负效应, 表明线虫营养类群对灌丛的响应依赖于所关注的灌木物种, 即不同植物种对线虫的影响不同[41]。(3)从cp类群水平上, 3种锦鸡儿属灌丛cp2类群正选择性物种比例大于负选择性物种比例, cp3-5类群的负选择性物种比例更大。这说明土壤线虫机会主义者(cp2)对土壤食物资源增加的响应程度强于定居者(cp3-5)。由此我们推测灌丛内土壤线虫的生活史策略可能以偏r-对策为主[42]。这种演变的原因是较低cp值的线虫(比如:cp2类群)有较强的适应环境变化的能力, 而较高cp值的线虫(如:cp3)可能更适合比较干旱的环境[43];在锦鸡儿灌丛生长发育过程中, 外源营养不断向灌丛下聚集, 形成一个变化的营养环境, 从而可能有利于一些cp2类线虫种定居;而灌丛外的干旱环境更适宜cp3-5类线虫种定居。另外, 灌丛内cp2类群指示物种主要是丽突属、真滑刃属、丝尾垫刃属, 灌丛外cp3-5类群指示物种主要是矛线目(如矮化属、短体属、真矛线属、桑尼属)。这与Guan等[44]研究结果一致, 即丽突属和矛线属可作为灌木的指示物种。由于不同的锦鸡儿属灌木会形成特异的土壤环境, 因此线虫会迁移到适合自己生长的栖息地。这也是导致这些指示物种多度具有明显的灌木种间差异的原因(表 2)。
第二, 在草原化荒漠区, 锦鸡儿属灌丛可能主要是通过根系分泌物、凋落物质量等因素, 而非土壤理化性质和林下植物群落影响土壤线虫群落组成。以往的研究主要通过结构方程模型(SEM)表明灌丛通过影响土壤理化性质(如:pH、土壤有机质、土壤含水量等)和林下植物群落影响土壤线虫群落组成[34]。我们通过Mantel tests发现, 绝大多数土壤理化性质和所有植物群落特性对线虫功能团组成影响不显著, 而土壤速效钾、铵态氮和土壤含水量与cp值为2的线虫功能团组成呈显著正相关(表 3), 这是因为低营养级线虫直接依赖土壤营养。
土壤线虫组成的变化引起线虫代谢足迹发生明显变化。灌丛内土壤线虫功能代谢足迹(菱形总面积)大于灌丛外(除狭叶锦鸡儿外), 说明灌丛内土壤线虫群落对碳的利用率更高。这可能是因为灌丛内食真菌类线虫比例较大, 且全是cp2类群线虫(表 1)。cp值较小的线虫通常为r-对策者, 个体较小, 生命周期相对短, 代谢快, 对外源养分的输入反应非常迅速, 从而导致其较高的资源利用率。不同灌木线虫代谢足迹不同。3种锦鸡儿属灌木中, 狭叶锦鸡儿的结构代谢足迹最大, 表明相比于其它类群线虫, 狭叶锦鸡儿对捕食杂食类线虫代谢活性的促进作用可能更强。这是因为狭叶锦鸡儿灌丛生物量大, 灌丛对土壤营养富集状况好, 土壤食物网结构成熟稳定, 捕食杂食类线虫比例最大(表 1), 所以更多的物质和能量从被捕食者(较低营养级)向捕食者(较高营养级)流动。富集指数可用于评价土壤养分富集状况, 值越大, 表明外界投入的养分越多, 土壤线虫食物资源越丰富[20]。我们研究发现3种锦鸡儿属灌丛的富集指数(菱形中心位置)均高于灌丛外, 说明灌丛内土壤养分富集状况好于灌丛外, 这是灌丛的资源岛效应。
土壤线虫组成的变化通过线虫代谢足迹导致土壤食物网结构显著变化。基于线虫代谢足迹的土壤线虫区系分析结果表明, 狭叶锦鸡儿灌丛在C象限, 比灌丛外更靠近B象限, 说明灌丛内土壤干扰程度低, 营养富集状况好, 食物网结构更成熟稳定;垫状锦鸡儿和荒漠锦鸡儿灌丛在D象限, 比灌丛外更靠近A象限, 说明土壤环境干扰程度高, 营养富集较好, 食物网有退化倾向。形成这些结果的原因是:(1)锦鸡儿属灌丛具有富集土壤营养和改善土壤环境效应;(2)狭叶锦鸡儿灌丛生物量大、灌丛结构致密, 资源岛效应和环境改善效应更明显, 而垫状锦鸡儿矮小、荒漠锦鸡儿稀疏, 二者资源岛效应较弱, 土壤环境更容易受外界干扰;(3)虽然灌丛具有资源岛效应和环境改善效应, 但草原化荒漠区土壤的营养仍然处于较贫瘠状态, 环境干扰和低营养依然是调控线虫群落组成的主要因素。
总之, 虽然锦鸡儿属灌丛没有增加土壤线虫多度、丰富度和多样性, 但却引起土壤线虫群落组成和结构发生显著变化, 且这种变化具有明显种间差异。土壤线虫组成的变化引起线虫代谢足迹发生明显变化, 进一步导致土壤食物网结构的显著变化。
| [1] |
Van Auken O W. Shrub invasions of North American semiarid grasslands. Annual Review of Ecology and Systematics, 2000, 31(1): 197-215. DOI:10.1146/annurev.ecolsys.31.1.197 |
| [2] |
熊小刚, 韩兴国, 鲍雅静. 试论我国内蒙古半干旱草原灌丛沙漠化的研究. 草业学报, 2005, 14(5): 1-5. DOI:10.3321/j.issn:1004-5759.2005.05.001 |
| [3] |
彭海英, 李小雁, 童绍玉. 内蒙古典型草原小叶锦鸡儿灌丛化对水分再分配和利用的影响. 生态学报, 2014, 34(9): 2256-2265. |
| [4] |
郭璞, 解李娜, 满良, 王金龙, 李清芳, 马成仓. 荒漠化草原锦鸡儿属灌丛扩增对牧草产量和植物多样性的影响. 草业科学, 2019, 36(5): 1215-1223. |
| [5] |
Na X F, Xu T T, Li M, Zhou Z N, Ma S L, Wang J, He J, Jiao B Z, Ma F. Variations of bacterial community diversity within the rhizosphere of three phylogenetically related perennial shrub plant species across environmental gradients. Frontiers in Microbiology, 2018, 9: 709. DOI:10.3389/fmicb.2018.00709 |
| [6] |
Liu R T, Zhao H L, Zhao X Y, Drake S. Facilitative effects of shrubs in shifting sand on soil macro-faunal community in Horqin Sand Land of Inner Mongolia, Northern China. European Journal of Soil Biology, 2011, 47(5): 316-321. DOI:10.1016/j.ejsobi.2011.07.006 |
| [7] |
Kudrin A A, Konakova T N, Taskaeva A A. Communities of soil nematodes of various tundra phytocenoses differing in the development level of the shrub layer. Russian Journal of Ecology, 2019, 50(6): 526-534. DOI:10.1134/S1067413619060092 |
| [8] |
Pen-Mouratov S, Barness G, Steinberger Y. Effect of desert plant ecophysiological adaptation on soil nematode communities. European Journal of Soil Biology, 2008, 44(3): 298-308. DOI:10.1016/j.ejsobi.2008.03.005 |
| [9] |
Mulder C, Maas R. Unifying the functional diversity in natural and cultivated soils using the overall body-mass distribution of nematodes. BMC Ecology, 2017, 17(1): 36. DOI:10.1186/s12898-017-0145-9 |
| [10] |
Li X P, Zhu H M, Geisen S, Bellard C, Hu F, Li H X, Chen X Y, Liu M Q. Agriculture erases climate constraints on soil nematode communities across large spatial scales. Global Change Biology, 2020, 26(2): 919-930. DOI:10.1111/gcb.14821 |
| [11] |
Ferris H, Bongers T. Indices developed specifically for analysis of nematode assemblages//Wilson M J, Kakouli-Duarte T, eds. Nematodes as Environmental Indicators. Wallingford: CABI, 2009: 124-145.
|
| [12] |
Ferris H. Form and function: metabolic footprints of nematodes in the soil food web. European Journal of Soil Biology, 2010, 46(2): 97-104. DOI:10.1016/j.ejsobi.2010.01.003 |
| [13] |
Wang Y M, Guan P T, Chen J W, Li Z X, Yang Y R, Wang P. A comparison of soil nematode community structure and environmental factors along fen-bush-forest succession in a peatland, northeastern China. Global Ecology and Conservation, 2021, 28: e01679. DOI:10.1016/j.gecco.2021.e01679 |
| [14] |
Wu Y, Chen W J, Entemake W, Wang J, Liu H F, Zhao Z W, Li Y Z, Qiao L L, Yang B, Liu G B, Xue S. Long-term vegetation restoration promotes the stability of the soil micro-food web in the Loess Plateau in North-west China. CATENA, 2021, 202: 105293. DOI:10.1016/j.catena.2021.105293 |
| [15] |
Jackson L E, Bowles T M, Ferris H, Margenot A J, Hollander A, Garcia-Palacios P, Daufresne T, Sánchez-Moreno S. Plant and soil microfaunal biodiversity across the borders between arable and forest ecosystems in a Mediterranean landscape. Applied Soil Ecology, 2019, 136: 122-138. DOI:10.1016/j.apsoil.2018.11.015 |
| [16] |
Hodson A K, Ferris H, Hollander A D, Jackson L E. Nematode food webs associated with native perennial plant species and soil nutrient pools in California riparian oak woodlands. Geoderma, 2014, 228-229: 182-191. DOI:10.1016/j.geoderma.2013.07.021 |
| [17] |
Veen G F, Olff H, Duyts H, van der Putten W H. Vertebrate herbivores influence soil nematodes by modifying plant communities. Ecology, 2010, 91(3): 828-835. DOI:10.1890/09-0134.1 |
| [18] |
Cortois R, Veen G F, Duyts H, Abbas M, Strecker T, Kostenko O, Eisenhauer N, Scheu S, Gleixner G, De Deyn G B, van der Putten W H. Possible mechanisms underlying abundance and diversity responses of nematode communities to plant diversity. Ecosphere, 2017, 8(5): e01719. DOI:10.1002/ecs2.1719 |
| [19] |
周广帆, 秦洁, 祁小旭, 王瑞琪, 赵建宁, 谭炳昌, 杨殿林. 丹江口水源涵养区退耕还草土壤线虫群落变化特征. 农业资源与环境学报, 2020, 37(3): 308-318. |
| [20] |
陈云峰, 韩雪梅, 李钰飞, 胡诚. 线虫区系分析指示土壤食物网结构和功能研究进展. 生态学报, 2014, 34(5): 1072-1084. |
| [21] |
薛会英, 胡锋, 罗大庆. 藏北高寒草甸植物群落对土壤线虫群落功能结构的影响. 生态学报, 2013, 33(5): 1482-1494. |
| [22] |
Olatunji O A, Gong S X, Tariq A, Pan K W, Sun X M, Chen W K, Zhang L, Dakhil M A, Huang D, Tan X. The effect of phosphorus addition, soil moisture, and plant type on soil nematode abundance and community composition. Journal of Soils and Sediments, 2019, 19(3): 1139-1150. DOI:10.1007/s11368-018-2146-5 |
| [23] |
Sun F, Pan K W, Olatunji O A, Li Z L, Chen W K, Zhang A P, Song D G, Sun X M, Huang D, Tan X. Specific legumes allay drought effects on soil microbial food web activities of the focal species in agroecosystem. Plant and Soil, 2019, 437(1/2): 455-471. |
| [24] |
Viglierchio D R, Schmitt R V. On the methodology of nematode extraction from field samples: baermann funnel modifications. Journal of Nematology, 1983, 15(3): 438-444. |
| [25] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
|
| [26] |
谢辉. 植物线虫分类学. 合肥: 安徽科学技术出版社, 2000.
|
| [27] |
Bongers T. The Nematodes of the Netherlands. Utrecht: Foundation Publisher of Royal Dutch Natural History Society, 1994.
|
| [28] |
鲍士旦. 土壤农化分析 ((第三版)). 北京: 中国农业出版社, 2000.
|
| [29] |
Yeates G W, Bongers T, de Goede R G, Freckman D W, Georgieva S S. Feeding habits in soil nematode families and genera—an outline for soil ecologists. Journal of Nematology, 1993, 25(3): 315-331. |
| [30] |
Bongers T. The maturity index: an ecological measure of environmental disturbance based on nematode species composition. Oecologia, 1990, 83(1): 14-19. DOI:10.1007/BF00324627 |
| [31] |
Ferris H, Bongers T, de Goede R G M. A framework for soil food web diagnostics: extension of the nematode faunal analysis concept. Applied Soil Ecology, 2001, 18(1): 13-29. DOI:10.1016/S0929-1393(01)00152-4 |
| [32] |
Michonneau F, Brown J W, Winter D J. rotl: an R package to interact with the Open Tree of Life data. Methods in Ecology and Evolution, 2016, 7(12): 1476-1481. DOI:10.1111/2041-210X.12593 |
| [33] |
Dufrêne M, Legendre P. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs, 1997, 67(3): 345-366. |
| [34] |
Wang X T, Nielsen U N, Yang X L, Zhang L M, Zhou X H, Du G Z, Li G X, Chen S Y, Xiao S. Grazing induces direct and indirect shrub effects on soil nematode communities. Soil Biology and Biochemistry, 2018, 121: 193-201. DOI:10.1016/j.soilbio.2018.03.007 |
| [35] |
Diakhaté S, Villenave C, Diallo N H, Ba A O, Djigal D, Masse D, Sembène P M, Chapuis-Lardy L. The influence of a shrub-based intercropping system on the soil nematofauna when growing millet in Senegal. European Journal of soil Biology, 2013, 57: 35-41. DOI:10.1016/j.ejsobi.2013.04.003 |
| [36] |
Tian L, Wang X A. Role of nurse shrubs for restoration planting of two conifers in southeast of Mu Us Sandland, China. Journal of Environmental Biology, 2015, 36(1): 331-336. |
| [37] |
Togashi K, Hoshino S. Distribution pattern and mortality of the white tip nematode, Aphelenchoides besseyi (Nematoda: Aphelenchoididae), among rice seeds. Nematology, 2001, 3(1): 17-24. DOI:10.1163/156854101300106847 |
| [38] |
Togashi K, Hoshino S. Trade-off between dispersal and reproduction of a seed-borne nematode, Aphelenchoides besseyi, parasitic on rice plants. Nematology, 2003, 5(6): 821-829. DOI:10.1163/156854103773040727 |
| [39] |
Lavorel S, Garnier E. Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Functional Ecology, 2002, 16(5): 545-556. DOI:10.1046/j.1365-2435.2002.00664.x |
| [40] |
De Deyn G B, Raaijmakers C E, Van Ruijven J, Berendse F, Van Der Putten W H. Plant species identity and diversity effects on different trophic levels of nematodes in the soil food web. Oikos, 2004, 106(3): 576-586. DOI:10.1111/j.0030-1299.2004.13265.x |
| [41] |
Viketoft M, Palmborg C, Sohlenius B, Huss-Danell K, Bengtsson J. Plant species effects on soil nematode communities in experimental grasslands. Applied Soil Ecology, 2005, 30(2): 90-103. DOI:10.1016/j.apsoil.2005.02.007 |
| [42] |
Zhao J, Shao Y H, Wang X L, Neher D A, Xu G L, Li Z A, Fu S L. Sentinel soil invertebrate taxa as bioindicators for forest management practices. Ecological Indicators, 2013, 24: 236-239. DOI:10.1016/j.ecolind.2012.06.012 |
| [43] |
Yeates G W. Nematodes as soil indicators: functional and biodiversity aspects. Biology and Fertility of Soils, 2003, 37(4): 199-210. DOI:10.1007/s00374-003-0586-5 |
| [44] |
Guan P T, Zhang X K, Yu J, Ma N N, Liang W J. Variation of soil nematode community composition with increasing sand-fixation year of Caragana microphylla: bioindication for desertification restoration. Ecological Engineering, 2015, 81: 93-101. DOI:10.1016/j.ecoleng.2015.04.011 |