 2022, Vol. 42
2022, Vol. 42文章信息
- 施栋涛, 赵冲, 尹东红, 陈阳, 常亚青
- SHI Dongtao, ZHAO Chong, YIN Donghong, CHEN Yang, CHANG Yaqing
- 流速对光棘球海胆行为和生长的影响
- Effects of flow velocity on behaviors and growth of the sea urchin Mesocentrotus nudus
- 生态学报. 2022, 42(10): 4160-4168
- Acta Ecologica Sinica. 2022, 42(10): 4160-4168
- http://dx.doi.org/10.5846/stxb202006101508
-
文章历史
- 收稿日期: 2020-06-10
- 网络出版日期: 2022-01-11

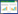




流速作为重要的环境物理因子, 对海洋生物的生存、生长及种群分布等均有重要的影响[1-4]。海胆是一类常见的棘皮动物, 多生活在水流变化较大的浅海和潮间带区域, 具有重要的生态和经济价值[5]。研究表明流速对海胆的影响在个体和种群层面都有表现。在个体层面, 海胆的移动[6-9]、觅食[10-11]及生殖[12-13]等行为在不同流速下均会表现出显著的差异。在种群层面, 水流速度影响海胆的栖息区域, 不同种海胆对栖息地的选择不同[14-16]。
正形海胆幼胆和成胆以大型藻类为食(如海带和石莼)。该类海胆的觅食过程主要涉及管足活动[17], 移动[18-19], 口器咬合[20-21]等行为。海胆利用管足进行移动, 管足的粘附能力可以帮助海胆粘附在大型海藻上, 海胆进而利用活动口器进行咬合来完成觅食活动[17, 22]。Tamaki等[23]的野外研究表明流速限制了海胆的觅食区域[24]。这可能是由于高流速的增加严重影响了海胆的运动和摄食[19]。实验室研究也证明了这一点, 在24 h的单向水流作用下, 海胆的摄食率随着流速的增加而下降[11]。翻正行为是海胆在口面向上的情况下, 利用管足使其口面向下的行为模式, 是测试海胆活力的有效性状[25-26]。本研究之前的短期实验结果表明, 海胆的翻正行为受到高流速抑制[9]。海胆的体尺也受到不同区域流速的影响。Cohen Rengifo等[27]在不同流速海区采集的同种海胆在体尺大小, 管足的数目和韧性均出现了显著差异。目前关于流速对海胆影响的报道主要集中在实验室短期(0-2 d)研究[9, 11, 19]和野外观察上[19, 23, 26]。短期研究难以阐明流速的长期影响, 而野外海区环境复杂, 海胆易受到流速之外的多种非实验因素的干扰。因此, 开展相对长时间的实验室研究对揭示流速对海胆行为和生长的影响具有重要意义。
光棘球海胆(Mesocentrotus nudus)作为我国北方沿海潮间带地区的常见物种, 具有重要的生态和经济价值。国家海洋信息中心流速数据表明, 大连海洋牧场相关海域大部分时间流速稳定(< 20 cm/s)且有良好的水流交换, 适合海洋经济生物的底播增殖[28]。Kawamata[19]和Tamaki[23]的短期实验表明20 cm/s流速下, 海胆的摄食率显著下降, 超过20 cm/s流速区域海胆数目显著减少。依据Shi等[9]之前的结果, 流速稳定时进行底播可减小流速对海胆增殖放流的影响, 但其在长期水流影响下的生长和摄食能力仍缺乏研究。因此, 本文根据现有研究和海区流速环境确定10 cm/s和20 cm/s为本研究的中和高流速, 2 cm/s为自然海区较低的流速。本实验主要研究了光棘球海胆1)在不同流速下的生存和生长情况;2)在不同流速下的觅食和摄食行为;3)在不同流速下管足附着和翻正行为。本研究有助于进一步理解流速对海胆的长期影响, 为进一步阐明海胆底播增殖的适宜流速提供理论参考。
1 材料与方法 1.1 海胆本实验用光棘球海胆全部来源于大连市金州某育苗场(壳径=(21.57±0.14)mm, 壳高=(10.12±0.10)mm, 体重=(4.19±0.08)g)。实验前海胆暂养于农业农村部北方海水增养殖重点实验室。实验使用海胆均健康, 无疾病症状。暂养池为玻璃钢水槽(长×宽×高=180 cm×100 cm×70 cm), 期间饲以新鲜海带(足量投喂, 每2 d约10 g/只)。此外, 本研究每两日换水1/3, 清理粪便。实验室光照为自然光, 暂养期间连续充气确保海胆不会缺氧。
1.2 实验设计为了研究流速对海胆的长期影响, 实验设置在低(2 cm/s), 中(10 cm/s)和高(20 cm/s)3种流速下进行。
本实验使用环形跑道(图 1)进行, 跑道由网分割出5个体积相等小室(长×宽×高=20 cm×15 cm×10 cm)。每一小室底部放置一块底质模拟板。底质板由沙子(采集于黑石礁海边, 由1 mm孔径筛网筛出, 约50 g)和石头(采集于黑石礁海边, 约40 g, 约15个)作为底质, 使用环氧树脂(20 g)经行粘结。水流为单向流动(如图 1), 由入口进入后, 经过各小室流至出水口。出水口挂置一个过滤网袋过滤海胆粪便。食物由绑在底质板上的塑料夹子夹住。本研究对实验过程中其它发现也进行记录。海胆体尺性状使用游标卡尺(Mahr, 德国)测量, 重量使用电子天平(G & G, USA)测量。实验开始前利用流速仪(JDC, Swiss)调整流速至目标流速, 并保证每个小室的流速差距在±0.5 cm/s。为保证试验期间水质, 保证充气充足并每日换水1/3, 日常测量其温度, pH和盐度。实验进行时, 每一小室放置5只海胆, 每一流速和每一小室间的海胆在体尺测量上均无统计学差异。

|
| 图 1 长期流速实验装置示意图 Fig. 1 Diagram of the equipment for the long-term flow velocity experiment 水流为单向, 由入水口进入后, 经整流板进入实验区后自出口流出。实验区有5个小室, 每一个小室为一个实验区(长×宽×高=20 cm×15 cm×10 cm), 其间用网相隔, 底部放置有底质模拟板 |
为了评估长期水流对海胆摄食的影响, 本研究测量了海胆每日的摄食量。每个小室每次投食均使用相同部位并切成大小约11 cm×5 cm的新鲜海带(约20 g)。每日定时投喂海带, 并称量前一日投喂海带的剩余重量, 以确定其每日的摄食情况, 测量共39次(n=5)。在测量摄食期间, 对每一流速下的海胆的粪便留存量进行了3次测量。
1.4 生长实验进行中, 为了研究海胆在不同流速下的生长的情况, 每周对其进行生长相关性状的测量。本实验中, 测量性状包括海胆的壳径(mm), 壳高(mm)和体重(g)。每个小室中的5个海胆平均值作为一个独立样本数值进行分析(n=5)。最后, 本研究中计算了海胆的增重率(WGR), 特定生长率(SGR)和饲料转化率(FCR), 公式如下:
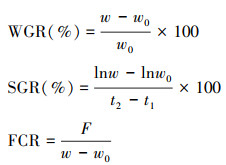
|
式中, w0表示海胆初始体重, w表示养殖后体重, t1表示开始养殖的时间, t2表示终末时间, F表示总摄食量。
1.5 翻正行为在长期流速影响的养殖实验结束后, 立即对海胆的翻正行为进行测试。这一实验分别在原位(摄食实验时所暴露的流速)和静水中进行。将海胆口面朝上放置于装置底部, 记录海胆利用其管足和棘调整至反口面朝上的正常体态所需时间, 若海胆超过300 s仍未恢复到口面向下的姿态, 则翻正时间记为300 s[9]。
1.6 觅食时间和附着时间在长期流速影响的养殖试验后, 本研究测量了海胆在不同流速下的觅食能力。在原流速下进行了7 d禁食处理后, 将新鲜海带置于上游区域, 并将海胆放置于下游区域中间(两个小室大小, 长×宽=40 cm×15 cm), 观察海胆的移动和觅食情况。本实验全程使用摄像机(SONY, FDX-55)进行摄像, 实验后进行计算和分析。在共5个小室中, 每个小室内随机挑选2只海胆进行实验(n=10)。觅食行为时长持续为480 s, 若超过这一时间海胆仍未到达食物点, 则将觅食时间记为480 s[29]。
海胆觅食实验前, 进行海胆管足附着能力的测量。本研究测量了海胆附着的时间以确定水流对管足的附着能力的影响。将3种不同流速下养殖的海胆放置于与其暴露流速相同的流速下, 记录其用管足将自身附着于底面所需的时长。实验开始时将海胆放置于底面并开始计时, 当其管足附着于底面不被水流移位且可移动时记为附着完成, 期间为附着时间。
1.7 口器咬合行为为了研究长期暴露在不同流速下的海胆的摄食能力, 在觅食行为后5 d(期间正常投喂)进行了口器咬合行为的实验。实验使用定制的亚克力器具如图 2所示。实验开始前, 根据Shi等[30]的方法配置海藻胶并铺于容器内, 待其凝固后加满相同温度海水。实验时, 将海胆放入容器中, 并在上方盖一聚乙烯塑料泡沫固定。使用摄影机(SONY, FDX-55)在容器下方进行拍摄。在共5个小室中, 每个小室内随机挑选2只海胆进行实验(n=10)。实验后通过录像计数海胆在300 s内的口器咬合次数。

|
| 图 2 口器咬合行为实验装置示意图 Fig. 2 Experimental facility for Aristotle′s lantern reflex of Mesocentrotus nudus |
本研究数据均使用SPSS(vision 22)进行分析。分析前分别用Kolmogorov-Smirnov和Levene′s的方法检查数据的正态分布情况和方差齐性。海胆的初始壳径、壳高和体重使用单因素方差分析。养殖期间海胆摄食量、生长性状和之后测试的翻正行为使用单因素方差分析, 如有显著差异则进一步使用LSD方法进行两两比较。对不符合正态分布或方差齐性的觅食行为和管足附着时间数据进行非参数检验分析(Kruskal-Wallis方法)。显著性概率水平为P < 0.05。
2 实验结果 2.1 摄食量养殖试验中测量海胆每日摄食量39次, 期间无海胆死亡。结果表明在流速为2 cm/s, 10 cm/s和20 cm/s时的每日平均摄食量分别为(5.94±1.87)g, (5.22±1.90)g和(4.59±1.81)g。其中流速为2 cm/s时的每日摄食量显著大于10 cm/s和20 cm/s(P < 0.001和P < 0.001), 而10 cm/s时的摄食量显著大于20 cm/s(P=0.001)(图 3)。

|
| 图 3 光棘球海胆在2 cm/s, 10 cm/s和20 cm/s下的每日平均摄食量(平均值±标准差) Fig. 3 Daily feeding of Mesocentrotus nudus at 2 cm/s, 10 cm/s and 20 cm/s (mean±SD) 不同小写字母表示在不同流速下日平均摄食量存在显著性差异 |
此外, 在实验中发现一定的水流对海胆的粪便有清理作用。当流速为2 cm/s时底板和海胆反口面部会残留部分粪便((4.66±1.85)g, 图 4), 而在共3次的测量中流速为10 cm/s和20 cm/s时则没有粪便残留(n=3)。

|
| 图 4 光棘球海胆于2 cm/s时, 壳面部分留有粪便 Fig. 4 Left feces on the top of the urchin test at 2 cm/s |
在对海胆壳径和体重的7周连续测量中, 自第2周和第3周, 海胆的体重和壳径就开始在不同流速组间出现显著差异(表 1)。
| 流速 Flow velocity/(cm/s) |
壳径 Test diameter/mm |
显著性 Sig. |
体重 Body weight/g |
显著性 Sig. |
|
| 实验起始Week 0 | 2 | 21.53±1.04 | 0.398 | 9.84±0.69 | 0.520 |
| 10 | 21.39±1.24 | 10.30±0.84 | |||
| 20 | 21.77±1.42 | 10.22±0.96 | |||
| 第1周Week 1 | 2 | 22.45±1.04 | 0.175 | 10.28±0.67 | 0.226 |
| 10 | 22.7±1.32 | 10.57±0.77 | |||
| 20 | 22.18±1.39 | 10.44±0.88 | |||
| 第2周Week 2 | 2 | 23.37±1.01 | 0.743 | 11.04±0.72a | 0.015 |
| 10 | 23.19±1.17 | 11.06±0.68ab | |||
| 20 | 23.15±1.24 | 10.75±0.81b | |||
| 第3周Week 3 | 2 | 24.45±0.93a | 0.002 | 11.36±0.68a | 0.001 |
| 10 | 23.88±1.13b | 11.19±0.89a | |||
| 20 | 23.41±1.34b | 11.23±0.83b | |||
| 第4周Week 4 | 2 | 25.36±1.04a | 0.001 | 11.74±0.80a | 0.001 |
| 10 | 24.61±1.33b | 11.37±0.57b | |||
| 20 | 24.17±1.43b | 11.36±1.04c | |||
| 第5周Week 5 | 2 | 25.92±1.06a | 0.002 | 11.60±0.86a | 0.001 |
| 10 | 25.44±1.24a | 11.92±0.80b | |||
| 20 | 24.50±1.73b | 11.05±0.83c | |||
| 第6周Week 6 | 2 | 26.91±1.25a | 0.001 | 11.74±0.77a | 0.004 |
| 10 | 25.75±1.54b | 11.39±0.86b | |||
| 20 | 24.86±1.78c | 11.16±0.79b | |||
| 第7周Week 7 | 2 | 27.22±1.17a | 0.003 | 11.83±0.62a | 0.005 |
| 10 | 26.13±1.49b | 11.49±0.85b | |||
| 20 | 25.35±1.87b | 11.34±0.82b | |||
| 不同小写字母表示在各实验组中存在显著性差异 | |||||
在49 d中, 光棘球海胆在2 cm/s时的增重率(图 5)显著大于10 cm/s和20 cm/s组(P=0.015和P=0.001), 而后两者间无显著差异(P=0.099)。特定增长率(图 5)也表明流速显著地影响了光棘球海胆的生长。流速为2 cm/s时海胆的特定增长率显著大于10 cm/s和20 cm/s(P=0.007和P < 0.001), 而后两者无显著差异(P=0.060)。饲料转化率(图 5)在流速为20 cm/s时要显著大于2 cm/s(P=0.045), 而2 cm/s和10 cm/s, 10 cm/s和20 cm/s间无显著差异(P=0.057和P=0.899)。

|
| 图 5 在2 cm/s, 10 cm/s和20 cm/s流速下光棘球海胆的增重率、特定生长率、饲料转化率变化 Fig. 5 Weight gain rate, specific growth rate and food coefficient of Mesocentrotus nudus at flow velocities of 2 cm/s, 10 cm/s and 20 cm/s 不同小写字母表示在各实验组中存在显著性差异 |
流速显著影响海胆的管足的粘附时间(图 6)。在10 cm/s和20 cm/s下, 海胆的管足粘附时间显著长于在2 cm/s下(P < 0.001和P < 0.001)。在10 cm/s和20 cm/s组之间无显著差异(P=0.051)。

|
| 图 6 光棘球海胆在2 cm/s, 10 cm/s和20 cm/s流速下长期暴露后的行为 Fig. 6 The behaviors of Mesocentrotus nudus at flow velocities of 2 cm/s, 10 cm/s and 20 cm/s 不同小写字母表示在不同实验组中存在显著性差异 |
光棘球海胆在不同流速下的觅食时间存在显著差异。在20 cm/s流速下, 海胆的觅食时间显著多于在2 cm/s时(P=0.004), 但是在10 cm/s和20 cm/s组之间无显著差异(P=0.200)(图 6)。
2.5 口器咬合海胆口器咬合在2 cm/s, 10 cm/s和20 cm/s流速下无显著差异, 但是在20 cm/s的流速下咬合次数相对更多(P=0.113)(图 6)。
2.6 翻正行为在原位翻正行为测试中, 相对于2 cm/s(P=0.02)和10 cm/s(P=0.03), 当流速为20 cm/s时海胆的翻正时间显著增加。海胆原位翻正时间在2 cm/s和10 cm/s之间没有显著差异(P=0.840, 图 6)。
在静水中测试的海胆翻正时间结果表明, 在2 cm/s, 10 cm/s和20 cm/s下长时间暴露的各组海胆翻正时间之间无显著差异(P=0.208)。
3 讨论栖息于潮间带和浅海,海胆的分布、行为和生长更容易受到水流环境变化的影响[11, 26, 31]。因此, 研究海胆在长期不同流速影响下的生存、生长和行为, 具有重要的生态和经济价值。本实验研究了长时间(49 d)不同流速(2 cm/s, 10 cm/s和20 cm/s)对光棘球海胆的生存、生长和行为的影响, 为进一步阐明海胆底播增殖的适宜流速提供理论参考。
野外调查显示流速影响海胆的生长[27]。本研究表明海胆的壳径和体重的增长在高流速下均受到了显著抑制, 这一现象在第2、3周已表现出来。海胆的增重率为2 cm/s组 > 10 cm/s组和2 cm/s组 > 20 cm/s组。这表明高流速抑制了海胆体重的增长。特定生长率与增重率结果同样为2 cm/s组 > 10 cm/s组和2 cm/s组 > 20 cm/s组。这进一步表明高流速对海胆生长的抑制作用。该结果与野外调查有一定相似性, 在流速较高的区域收集的海胆在体尺上小于流速较低区域[27]。本文推测海胆增重受到抑制可能是摄食量受到影响的结果。进一步研究发现, 海胆摄食量在不同流速下为2 cm/s组 > 10 cm/s组 > 20 cm/s组。这说明在不同流速下的摄食量对海胆生长可能具有较大影响。海胆的觅食或者摄食(口器咬合)能力下降可能进一步导致了摄食量的下降。然而, 海胆口器咬合次数并未在3种流速下产生显著差异, 这说明海胆的口器摄食功能稳定性好, 与Shi等[30]的研究结果相似。Tamaki等[23]的野外调查结果显示光棘球海胆几乎不会出现在较高流速(14.5±3.4)cm/s但食物更为丰富的区域。这与本实验结果相一致, 光棘球海胆在低流速下的觅食所需时间随着流速的升高而增加(10 cm/s组 > 2 cm/s组, 20 cm/s组 > 2 cm/s组)。这表明较高流速削弱了海胆觅食能力。因此, 高流速下每日摄食量的减少可能是由于流速对光棘球海胆觅食能力的影响, 这也与Tamaki等[11]的短期实验结果相一致。较低流速下海胆持续增长以较高的生长率为特征, 而体型的增长可能会帮助其建立抵抗水流的优势[11, 19]。因此, 中低流速区域(2-10 cm/s)更适合海胆生存、摄食和生长。低流速海域的大型藻类可能会因海胆的大量摄食而减少, 进而影响低流速区域的生态环境[24]。
海胆的觅食行为需要依靠管足的粘附-脱离-粘附过程来完成。这也是其附着、移动等行为的基础[32]。在高流速下海胆生长更缓慢, 可能是因为管足消耗额外的能量用来抵御水流影响, 而不能用于生长[33-34]。本研究发现饲料转化率在20 cm/s下显著大于低流速(2 cm/s), 这表明光棘球海胆的增重所消耗的食物(海带)随着流速的增加而增加。管足附着时间实验表明管足吸附能力受水流显著影响, 长时间流速暴露后海胆的觅食能力仍会受到水流的显著影响。海胆管足附着所需时间在3种流速下存在着显著差异, 中高流速暴露下海胆要消耗更长时间才能完成附着, 这可能是能量消耗的主要原因之一。而饲料转化率在不同流速下的海胆中无显著差异。这说明当流速升高到一定程度后, 海胆需要用于保持附着所需的消耗可能会趋于平衡[35]。此外, 较高流速(10 cm/s和20 cm/s)下海胆的粪便几乎不会在原地残留。本研究表明, 在保证食物供应的前提下, 流速为2-10 cm/s的区域有利于海胆生长。
翻正行为帮助海胆在不利环境(如海流)造成姿态失常后恢复正常。这一行为主要由管足活动影响, 是判断海胆环境适应性的重要指标[26]。在原位实验中, 海胆的翻正所需时间随着流速的增加而增加。这表明海胆的翻正能力会被削弱, 这也是管足吸附能力减弱的具体体现[36]。与之前的短期研究[9]结果不同, 长期的流速适应下的海胆均成功完成翻正并耗时更短。这说明海胆在长期高流速下, 或已经出现了一定适应性的表现, 但也无法排除体型差距可能造成的影响。Cohen Rengifo等[27]研究表明, 在不同流速区域生长的海胆在管足密度表现出了差别。这说明海胆对流速的适应很有可能会通过管足表现(例如, 翻正行为)来实现[26, 33]。而海胆在静水中的翻正所需时间无显著差异, 这表明这一行为稳定性好, 在10 cm/s和20 cm/s长期的流速下未受到显著影响。
综上所述, 不同流速暴露显著影响光棘球海胆(壳径:约20 mm)壳径和体重的增长, 但对其生存无显著影响。海胆的生长随着流速的升高而显著降低。高流速(20 cm/s)显著削弱海胆管足的粘附能力, 进而显著降低其翻正, 移动和觅食行为, 但并不显著影响其口器咬合行为。饲料转化率结果表明高流速(20 cm/s)下海胆需要消耗更多饲料以维系其生长。这表明流速对海胆生长的影响可能不仅源于摄食量和觅食行为的差异, 也源于其生存的基本消耗差异。根据我们结果和国家海洋信息中心关于适于海胆底播增殖海域的流速数据, 本研究建议有关生产单位在综合考虑各种环境因子和生产要素的前提下, 在流速常年在2-10 cm/s的海区开展光棘球海胆幼胆底播增殖生产, 或对相关海域进行海洋牧场建设, 投放躲避物减少流速影响。
致谢: 感谢国家科技资源共享服务平台--国家海洋科学数据中心(http://mds.nmdis.org.cn/)提供数据信息。| [1] |
Clements J C, Hunt H L. Influence of sediment acidification and water flow on sediment acceptance and dispersal of juvenile soft-fshell clams (Mya arenaria L.). Journal of Experimental Marine Biology and Ecology, 2014, 453: 62-69. DOI:10.1016/j.jembe.2014.01.002 |
| [2] |
Abelson A, Denny M. Settlement of marine organisms in flow. Annual Review of Ecology and Systematics, 1997, 28: 317-339. DOI:10.1146/annurev.ecolsys.28.1.317 |
| [3] |
Duckworth A R, Battershill C N, Schiel D R. Effects of depth and water flow on growth, survival and bioactivity of two temperate sponges cultured in different seasons. Aquaculture, 2004, 242(1/4): 237-250. |
| [4] |
Lenihan H S, Peterson C H, Allen J M. Does flow speed also have a direct effect on growth of active suspension-feeders: an experimental test on oysters. Limnology and Oceanography, 1996, 41(6): 1359-1366. DOI:10.4319/lo.1996.41.6.1359 |
| [5] |
Agatsuma Y. Strongylocentrotus nudus Developments in Aquaculture and Fisheries Science, 2013, 38: 449-460.
|
| [6] |
Lauzon-Guay J S, Scheibling R E. Seasonal variation in movement, aggregation and destructive grazing of the green sea urchin (Strongylocentrotus droebachiensis) in relation to wave action and sea temperature. Marine Biology, 2007, 151(6): 2109-2118. DOI:10.1007/s00227-007-0668-2 |
| [7] |
James P, Siikavuopio S I. The effects of tank system, water velocity and water movement on survival, somatic and gonad growth of juvenile and adult green sea urchin, Strongylocentrotus droebachiensis. Aquaculture Research, 2015, 46(6): 1501-1509. DOI:10.1111/are.12303 |
| [8] |
Morse B L, Hunt H L. Effect of unidirectional water currents on displacement behaviour of the green sea urchin Strongylocentrous droebachiensis. Journal of the Marine Biological Association of the United Kingdom, 2013, 93(7): 1923-1928. DOI:10.1017/S002531541300060X |
| [9] |
Shi D T, Yin D H, Chen Y, Sun J N, Yang M F, Chang Y Q, Zhao C. Effects of flow velocity on fitness-related behaviours of the sea urchin Mesocentrotus nudus: new information on stock enhancement. Journal of the Marine Biological Association of the United Kingdom, 2020, 100(6): 963-967. DOI:10.1017/S0025315420000867 |
| [10] |
Kawamata S. Experimental evaluation of the anti-attachment effect of microalgal mats on grazing activity of the sea urchin Strongylocentrotus nudus in oscillating flows. Journal of Experimental Biology, 2012, 215(9): 1464-1471. DOI:10.1242/jeb.068411 |
| [11] |
Tamaki H, Muraoka D, Inoue T. Effect of water flow on grazing by the sea urchin (Strongylocentrotus nudus) in the presence of refuge habitat. Journal of Water and Environment Technology, 2018, 16(1): 30-39. DOI:10.2965/jwet.17-010 |
| [12] |
Mead K S, Denny M W. The effects of hydrodynamic shear stress on fertilization and early development of the purple sea urchin Strongylocentrotus purpuratus. The Biological Bulletin, 1995, 188(1): 46-56. DOI:10.2307/1542066 |
| [13] |
Thomas F I M, Kregting L T, Badgley B D, Donahue M J, Yund P O. Fertilization in a sea urchin is not only a water column process: effects of water flow on fertilization near a spawing female. Marine Ecology Progress Series, 2013, 494: 231-240. DOI:10.3354/meps10601 |
| [14] |
Tuya F, Cisneros-Aguirre J, Ortega-Borges L, Haroun R J. Bathymetric segregation of sea urchins on reefs of the Canarian Archipelago: role of flow-induced forces. Estuarine, Coastal and Shelf Science, 2007, 73(3/4): 481-488. |
| [15] |
Santos R, Flammang P. Estimation of the attachment strength of the shingle sea urchin, Colobocentrotus atratus, and comparison with three sympatric echinoids. Marine Biology, 2008, 154(1): 37-49. DOI:10.1007/s00227-007-0895-6 |
| [16] |
McClanahan T R. Predation and the distribution and abundance of tropical sea urchin populations. Journal of Experimental Marine Biology and Ecology, 1998, 221(2): 231-255. DOI:10.1016/S0022-0981(97)00130-5 |
| [17] |
Santos R, Hennebert E, Coelho A V, Flammang P. The echinoderm tube foot and its role in temporary underwater adhesion//Gorb S N, ed. Functional Surfaces in Biology. Dordrecht: Springer, 2009: 9-41.
|
| [18] |
James D W. Diet, movement, and covering behavior of the sea urchin Toxopneustes roseus in rhodolith beds in the Gulf of California, México. Marine Biology, 2000, 137(5): 913-923. |
| [19] |
Kawamata S. Effect of wave-induced oscillatory flow on grazing by a subtidal sea urchin Strongylocentrotus nudus (A. Agassiz). Journal of Experimental Marine Biology and Ecology, 1998, 224(1): 31-48. DOI:10.1016/S0022-0981(97)00165-2 |
| [20] |
Brothers C J, McClintock J B. The effects of climate-induced elevated seawater temperature on the covering behavior, righting response, and Aristotle's lantern reflex of the sea urchin Lytechinus variegatus. Journal of Experimental Marine Biology and Ecology, 2015, 467: 33-38. DOI:10.1016/j.jembe.2015.02.019 |
| [21] |
Ding J Y, Zheng D F, Sun J N, Hu F Y, Yu Y S, Zhao C, Chang Y Q. Effects of water temperature on survival, behaviors and growth of the sea urchin Mesocentrotus nudus: new insights into the stock enhancement. Aquaculture, 2020, 519: 734873. DOI:10.1016/j.aquaculture.2019.734873 |
| [22] |
Candia Carnevali M D, Wilkie I C, Lucca E, Andrietti F, Melone G. The Aristotle's lantern of the sea-urchin Stylocidaris affinis (Echinoida, Cidaridae): functional morphology of the musculo-skeletal system. Zoomorphology, 1993, 113(3): 173-189. DOI:10.1007/BF00394858 |
| [23] |
Tamaki H, Kusaka K, Fukuda M, Arai S, Muraoka D. Undaria pinnatifida habitat loss in relation to sea urchin grazing and water flow conditions, and their restoration effort in Ogatsu Bay, Japan. Journal of Water and Environment Technology, 2009, 7(3): 201-213. DOI:10.2965/jwet.2009.201 |
| [24] |
安鑫龙, 李雪梅, 徐春霞, 李志霞. 大型海藻对近海环境的生态作用. 水产科学, 2010, 29(2): 115-119. DOI:10.3969/j.issn.1003-1111.2010.02.014 |
| [25] |
Hagen N T. Is the righting response a useful indicator of functional well-being in the green sea urchin, Strongylocentrotus droehachiensis?//David B, Guille A, Feral J P, Roux M, eds. Echinoderms Through Time. Balkema: CRC Press, 1994: 693-698.
|
| [26] |
Challener R C, McClintock J B. In situ measurements of righting behavior in the common sea urchin Lytechinus variegatus: the importance of body size, substrate type, and covering material. Aquatic Biology, 2017, 26: 33-40. DOI:10.3354/ab00669 |
| [27] |
Cohen-Rengifo M, Moureaux C, Dubois P, Flammang P. Attachment capacity of the sea urchin Paracentrotus lividus in a range of seawater velocities in relation to test morphology and tube foot mechanical properties. Marine Biology, 2017, 164(4): 79. DOI:10.1007/s00227-017-3114-0 |
| [28] |
国家海洋科学数据中心. [2021-08-21]. http://mds.nmdis.org.cn/pages/visualization.html.
|
| [29] |
Chi X M, Shi D T, Ma Z H, Hu F Y, Sun J N, Huang X Y, Zhang L S, Chang Y Q, Zhao C. Carryover effects of long-term high water temperatures on fitness-related traits of the offspring of the sea urchin Strongylocentrotus intermedius. Marine Environmental Research, 2021, 169: 105371. DOI:10.1016/j.marenvres.2021.105371 |
| [30] |
Shi D T, Ding J Y, Zhang L L, Zhang L S, Sun J N, Chang Y Q, Zhao C. Effects of UV-B radiation on fitness related behaviors of the sea urchin Strongylocentrotus intermedius. Journal of Oceanology and Limnology, 2018, 36(5): 1681-1687. DOI:10.1007/s00343-018-7211-9 |
| [31] |
杨军, 刘永虎, 田涛, 张国胜, 陈勇, 尹增强, 王月, 陈辰. 模拟海底水流和光照条件下光棘球海胆行为特征及其聚礁效果的初步研究. 大连海洋大学学报, 2016, 31(2): 219-224. |
| [32] |
Smith J E. The activities of the tube feet of Asterias rubens L. I. The mechanics of movement and of posture. The Quarterly Journal of Microscopical Science, 1947, 88(1): 1-14. |
| [33] |
Cohen-Rengifo M, Agüera A, Detrain C, Bouma T J, Dubois P, Flammang P. Biomechanics and behaviour in the sea urchin Paracentrotus lividus (Lamarck, 1816) when facing gradually increasing water flows. Journal of Experimental Marine Biology and Ecology, 2018, 506: 61-71. DOI:10.1016/j.jembe.2018.05.010 |
| [34] |
Santos R, Da Costa G, Franco C, Gomes-Alves P, Flammang P, Coelho A V. First insights into the biochemistry of tube foot adhesive from the sea urchin Paracentrotus lividus (Echinoidea, Echinodermata). Marine Biotechnology, 2009, 11(6): 686-698. DOI:10.1007/s10126-009-9182-5 |
| [35] |
Santos R, Flammang P. Intra-and interspecific variation of attachment strength in sea urchins. Marine Ecology Progress Series, 2007, 332: 129-142. DOI:10.3354/meps332129 |
| [36] |
Lawrence J M. The effect of temperature-salinity combinations on the functional well-being of adult Lytechinus variegatus (Lamarck) (Echlnodermata, Echinoldea). Journal of Experimental Marine Biology and Ecology, 1975, 18(3): 271-275. DOI:10.1016/0022-0981(75)90111-2 |