 2022, Vol. 42
2022, Vol. 42文章信息
- 王凯, 王聪, 冯晓明, 伍星, 傅伯杰
- WANG Kai, WANG Cong, FENG Xiaoming, WU Xing, FU Bojie
- 生物多样性与生态系统多功能性的关系研究进展
- Research progress on the relationship between biodiversity and ecosystem multifunctionality
- 生态学报. 2022, 42(1): 11-23
- Acta Ecologica Sinica. 2022, 42(1): 11-23
- http://dx.doi.org/10.5846/stxb202105141263
-
文章历史
- 收稿日期: 2021-05-14
- 网络出版日期: 2021-08-17





2. 中国科学院大学, 资源与环境学院, 北京 100049
2. College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China
随着全球性的物种灭绝加快, 物种减少如何影响生态系统功能成为一个备受关注的问题, 生物多样性与生态系统功能关系研究愈发重要[1-2]。生物多样性是生物及其环境形成的生态复合体以及与此相关的各种生态过程的综合, 包括动物、植物、微生物和它们所拥有的基因以及它们与其生存环境形成的复杂的生态系统[3]。生态系统功能是指生态系统所体现的各种作用, 主要包括物质循环、能量流动和信息传递等方面[4], 是通过生物群落来实现的。生物多样性与生态系统功能关系研究最早可以追溯到19世纪初, 乔治·辛克莱用草地实验探究了物种多样性与生物量的关系, 其结果被达尔文引用到物种起源中[5]。直到今天, 生物多样性与生态系统功能关系仍然是一个研究热点。生物多样性作为生态系统功能与动态的主要决定因素已经在诸多研究中得到证实[2], 然而大多数研究集中于单一功能, 忽略了不同功能之间的权衡关系, 例如促进土壤养分周转通常会导致二氧化碳的释放, 从而促进作物生产, 但同时也会减少碳储存[6]。而生态系统功能本质上是多维的, 生物多样性会同时影响多个生态系统功能, 例如森林中树种多样性的增加会同时提高生产力、水土保持能力、温室气体减排等多种功能, 仅考虑单一生态系统功能会低估生物多样性的重要性[7]。因此有学者逐渐认识到应该同时考虑多种生态系统功能, 并随之提出生态系统多功能性(ecosystem multifunctionality, EMF)的概念。EMF考虑了生物多样性如何同时影响多个生态系统功能, 政策制定者和管理人员可以据此通过各种措施使生态系统功能维持在最合适水平, 从而实现资源利用最大化和生态系统的可持续发展。
生态系统多功能性(ecosystem multifunctionality, EMF)指的是“生态系统的整体功能”或“生态系统同时提供多种功能和服务的能力”[8-10]。这一概念最初源自Sanderson等[11]在植物多样性与牧场生态系统牧草生产力的关系研究中, 他们认为为了揭示牧场管理中生物多样性的巨大利用空间, 应该从更广泛的角度看待牧场的生态系统功能, 并总结了对人类和环境有价值的15种生态系统功能, 首次提出了多功能性(multifunctionality)的概念。2007年, Hector与Bagchi[8]首次定量分析了植物物种多样性同时对多个生态系统过程的影响, 发现维持EMF需要比维持单个生态系统功能需要更多的物种。Hector与Bagchi的研究是真正意义上第一项生物多样性与生态系统多功能性(biodiversity and ecosystem multifunctionality, BEMF)研究, 开创了BEMF研究的先河。在这之后, He等[12]、Zavaleta等[10]、Isbell等[13]分别从微观层面、不同时空尺度等角度开展了BEMF研究。时至今日, BEMF研究已经成为生态学领域的新的研究热点, 并初步形成了不同干扰下不同维度、不同尺度生物多样性对生态系统多功能性影响的概念框架(图 1)。

|
| 图 1 干扰下生物多样性对生态系统多功能性影响的概念框架 Fig. 1 Conceptual framework of impacts of biodiversity on ecosystem multifunctionality under disturbance |
物种多样性是生物多样性的简单度量, 指得是生态系统中动物、植物、微生物种类的丰富性, 通常用物种数(丰富度)来衡量, 有时也用物种分布的均匀程度(均匀度)来衡量, 也可以通过结合两种度量的比例统计量(如Shannon-Winner指数)来衡量[14]。在传统的生物多样性与生态系统功能研究中, 常使用植物物种多样性表示生物多样性探究其与生产力的关系[15], 随着研究的深入, 动物、植物、微生物物种多样性与EMF的关系也受到越来越多的关注。
1.1.1 动植物物种多样性与EMF最早的BEMF研究同样也是使用物种多样性代替生物多样性探究BEMF关系, Hector与Bagchi[8]最早进行了植物物种多样性—生态系统多功能性的定量研究, 开创了BEMF研究的先河, 在这之后又出现了大量相关研究。Maestre等[16]首次利用全球旱地的群落调查数据集, 揭示了物种多样性作为旱地EMF驱动因素的重要性。这一观点在水生生态系统、动物群落中同样也有发现[17-19]。此外, 除了物种丰富度, 有学者发现物种均匀度与EMF显著负相关, 并且物种丰富度高、空间格局随机、均匀度低的群落EMF较高[20]。
1.1.2 微生物物种多样性与EMF由于高通量测序、宏蛋白质组学等生物信息技术的出现, 微生物的多度、丰度与EMF的关系研究也日渐增多。贺纪正等[12]利用高通量分子生物学方法在微观尺度探究了土壤生态系统中细菌多样性与EMF的关系, 发现更多的生态系统功能需要更多的细菌分类群来维持。除了细菌, 也有研究探究不同微生物类型例如细菌、真菌对EMF的相对影响[21-24]。Li等[23]发现在北方森林土壤中, 真菌丰富度和群落组成与EMF呈显著正相关, 而细菌丰富度与其不相关, 并确定了腐生真菌对于维持EMF至关重要。
1.2 功能多样性与EMF在大多数生物多样性—生态系统功能关系的研究中, 人们常用物种多样性来代替生物多样性。但是不同的物种在生理、生态、形态特征等方面存在较大差异, 简单的物种多样性指标难以真实地体现物种的特征对生态系统功能的重要作用[25]。生态系统的功能不仅依赖于物种的数目, 而且依赖于物种所具有的功能性状。与功能性状相对应的生物多样性维度即是功能多样性, Tilman[26]定义功能多样性为影响生态系统功能的群落中所有物种以及有机体的功能特征值及其变动范围, 它是影响生态系统动态、稳定性、生产力、营养平衡和其他生态系统功能的多样性的组成部分。
功能多样性对生态系统功能的作用机制基于两个假说, 一是质量比假说[27], 一是生态位互补假说[28]。基于质量比假说, Garnier[29]提出了群落加权平均指数(CWM), 基于生态位互补假说, 学者们提出了诸多功能多样性指数, 常用的是Villeger等[9]提出的功能丰富度指数(FRic)、功能均匀度指数(FEve)、功能分异度指数(FDiv)等三个功能多样性指数。Mouillot等[30]利用欧洲草原生态系统数据首次探究了植物叶性状功能多样性对EMF的影响。之后, 学者们基于两种假说对植物性状功能多样性与EMF关系展开了更深入的探讨[31-33]。Valencia等[34]比较了旱地生态系统中植物结构、叶性状功能多样性在两种假说下对EMF的影响, 发现质量比假说与生态位互补假说对于解释EMF同等重要, 然而Mensah等[35]在南非森林生态系统中发现了相反的结论。当前研究的研究区域的生态系统类型不同, 干扰强度不同, 所考虑的生态系统功能也不同, 因此对于何种假说决定EMF还有有待探究。上述研究都是基于植物地上部性状, 也有研究探究植物地下根系性状功能多样性对EMF的影响, 如Fry等[36]发现细须根与更高的养分循环速率相关, 根结构的复杂性、生根深度的FDiv与EMF正相关。
1.3 系统发育多样性与EMF功能多样性较物种多样性可以更好地解释生态系统功能的变异, 但评估所有与生态系统相关的功能性状往往是不现实的, 所以随着研究的深入, 研究者基于遗传学的理论和方法, 提出了系统发育多样性。系统发育多样性描述了一个群落中物种间系统发育距离的总量, 它既受物种间亲缘关系的影响, 也受物种数量的影响[37]。因为近亲缘关系的物种常被假设具有相似的功能性状, 所以可以用系统发育信息来代替物种间功能性状的信息, 也可以解决功能性状过多或无法被测量的情况[38]。系统发育多样性较低的群落, 功能性状冗余, 种间竞争作用强烈;而系统发育多样性较高的群落, 功能性状重叠少, 可以通过生态位互补或通过增加具有重要功能特性的优势种来增加对生态系统功能的贡献[37]。
Tucker等[39]将70余种系统发育多样性指标分为了丰富度、发散度和规律性三类, 对于系统发育多样性与EMF的研究, 学者一般使用系统发育多样性(PD)指数来表征系统发育多样性, 发现其与EMF正相关[40-41]。Le Bagousse-Pinguet等[42]发现增加早期分化谱系的丰富度(PSV)可以增加EMF, 而最近进化谱系的丰富度(MNTD)与EMF的相关性较弱。但也有研究发现, 植物的系统发育多样性对EMF没有显著的直接影响, 而是通过增加林分结构的复杂性间接影响EMF[43]。
综上, 生物多样性的3个维度均是驱动EMF的重要因素, 大量研究结果表明生物多样性的增加有助于提高EMF。然而, 近期的几项研究发现物种多样性对生态系统多功能性的影响与对单个生态系统功能的影响相同, 增加物种多样性并不会提高生态系统多功能性[44-46]。此外, 大多数研究基于选择效应选取群落内的优势种来探究物种多样性与EMF的关系, 但近来有研究指出群落内的稀有种而不是优势种是EMF的主要驱动力, 这一结论被归因于“保险假说”[41, 47]。因此, 物种多样性增加是否有利于提高EMF还存在争议, 未来需要加强这一方面的探索。目前关于功能多样性与生态系统多功能性的关系研究集中于探究植物性状功能多样性对EMF的影响, 对于植物性状的选取大多侧重于物理性状, 很少选择物候或节律性状, 而并不是所有的物理性状都适合预测生态系统功能[48], 所以对于植物性状的选取应该谨慎, 应适当考虑植物物候、节律性状, 并考虑动物、微生物功能多样性对EMF的影响。在BEMF关系研究中系统发育多样性与生态系统多功能性的研究相对较少, 将系统发育多样性作为预测生态系统功能的重要生物因素纳入到BEMF研究中有助于厘清生物多样性对于EMF的作用关系。此外, 用来表征系统发育多样性的指标多达70余种, 各个指标之间的差异不甚明确, 需要进一步对指标进行归纳, 选择合适的指标与EMF结合进行下一步研究。
2 多营养级BEMF研究生态系统由多个营养级组成, 不同的营养级之间可能相互作用, 例如生产者利用分解者提供的无机物而分解者从生产者获取碳, 并且不同营养级对生态系统功能的效应可能相反或者互补, 此外, 不同营养级对生态系统功能的相对贡献也不同[49-51]。因此, 仅仅关注于单一营养级可能会低估生物多样性对EMF的影响。
近年来, 逐渐有学者关注多营养级生物多样性对EMF的影响。Eisenhauer等[52]最早探究了多营养级生物多样性对EMF的影响, 他们发现蚯蚓、等足类动物等分解者可以通过改变氮吸收、植物根系深度分布和地表凋落物影响植物间的互补作用, 从而提高植物物种丰富度和EMF之间的积极关系。
多营养级生物多样性如何影响生态系统多功能性的相关研究主要有以下3种方式。第一, 通过z-标准化或归一化方法综合地上、地下群落生物多样性, 分析其与EMF的关系。Jing等[53]采用z-均值法探究了青藏高原草原生态系统地上和地下群落的生物多样性如何影响EMF, 发现地上和地下物种多样性的综合效应比单独的任一因素能更好地解释不同地点间EMF的变异。第二, 通过构建不同营养级之间的级联效应探究BEMF关系。Schuldt等[54]在亚热带森林生态系统中发现促进分解和养分循环的异养生物的多样性, 以及植物功能性状的多样性和组成对多功能性的影响大于树种丰富度对其的影响, 并且通过较高营养级生物多样性对较低营养级生物多样性的级联效应表明, 多营养级的生物多样性是理解EMF驱动因素的关键。第三, 对各个营养级生物多样性对生态系统多功能性的效应进行比较。Lefcheck等[55]通过阈值法量化EMF, 探究了不同生境、不同营养级下生物多样性与EMF的关系, 结果表明死有机物、食腐者在不同阈值水平下物种多样性对EMF均有较弱的作用, 植物多样性对EMF有更强的效应, 但在高阈值时逐渐减弱, 而食草动物多样性在低阈值、高阈值下对EMF均有较强的效应, 并且上述物种多样性对EMF的效应在陆地生境和水生生境一致。
综上, 多营养级生物多样性相较于单一营养级生物多样性可以更好地解释EMF的变异。但是目前的研究还停留在探究多营养级物种多样性对EMF的影响, 功能多样性、系统发育多样性作为EMF的重要驱动因素受到关注较少。此外, 当前研究局限于陆地生境, 忽略了水生生境中的多营养级BEMF研究, 需要进一步扩大研究范围。
3 BEMF的时空尺度效应生态系统的功能, 如养分循环, 不仅受当前气候条件、生物多样性驱动, 也受过去生物、非生物因素驱动, 过去条件的遗留效应也可以通过土壤质地、植物功能性状、微生物群落等间接影响EMF。Ye等[56]探究了末次盛冰期(Last Glacial Maximum, LGM)以来的植被和气候条件的遗留效应对EMF的影响, 发现LGM时期植被覆盖度较低、物种多样性较少的群落对当前的EMF有直接的负面影响, 并且LGM以来经历了较高升温速率的生态系统, 每增温1℃, 其当代EMF降低约9%。此外, 当前BEMF关系的探究多基于1年内的短时间尺度, 长时间序列下BEMF关系的探究相对较少。有学者探究了温带松树林120年的次生演替过程中生物多样性与生态系统多功能性的变化, 发现随着演替的进行, 森林内的物种多样性增加进而提高了生态系统多功能性[57]。
随着BEMF研究的深入, 有研究发现群落的空间格局会影响资源的可利用性、生物多样性等[58-59], 进而影响EMF。已有学者发现群落组成的差异(β多样性)也会影响EMF[60-61], β多样性主要通过以下3种机制促进生态系统的多功能性。首先, 如果不同的物种在不同的地点发挥不同的功能, 那么对功能起重要作用的物种间的差异可以提高EMF[13, 62]。其次, 局地群落之间物种组成的差异影响群落之间的生态过程, 包括对生态系统功能至关重要的生物和资源的移动[63], 进而提高EMF;最后, 性状变异使不同的物种对不同的生态系统功能做出贡献, 可能导致功能权衡, 群落组成的差异将功能的表现在群落之间进行空间分离, 避免功能之间的权衡, 从而提高EMF[61]。Pasari等发现更大尺度的多样性(γ多样性)同样会对EMF有显著的正效应, 但α多样性对EMF的影响仍居于主导地位, 并远大于β多样性与γ多样性。Yan等[64]也发现α多样性、β多样性对草原土壤多功能性有显著的正效应, 但α多样性的作用更大。尽管如此, β多样性、γ多样性对EMF的影响同样重要。此外, 近来有研究表明α多样性和β多样性的协同作用会影响EMF, 在一个尺度上高水平的生物多样性放大了另一个尺度生物多样性对EMF的贡献[63]。
综上, 在不同的时空尺度需要不同的物种组合和丰富度水平以提供高水平的EMF[10], 在时间尺度上, 生物多样性与EMF的关系除受到累计影响外, 还应该考虑“遗留效应”;在空间尺度上, 群落间的物种差异相比于群落内的物种差异对于维持EMF同样重要, BEMF研究的时空尺度效应对于指导全球变化下生物多样性保护与生态系统管理具有重要的指导意义。如“遗留效应”为全球旱地留下了很大的“功能性债务”[56], 而在当今全球变化加剧的情况下, 极端气候、生物多样性丧失等可能会对未来的EMF造成不利影响, 需要引起重视。此外, 生物多样性保护和生态系统管理行动通常在不同的空间尺度上施行, 不同空间尺度的生物多样性及其协同交互均对生态系统多功能性有显著的正效应。因此未来的生物多样性保护和生态系统管理行动需要考虑BEMF的时空尺度效应, 以达到预期效果。
4 干扰下的BEMF研究干扰是存在于自然界中无时无处不在的一种现象, 会影响生态系统的结构和功能[65]。干扰按产生的来源可分为自然干扰和人为干扰, 自然干扰包括洪水、干旱、台风等, 人为干扰包括放牧、施肥、伐木等。在BEMF研究中, 干扰是重要的驱动因素, 会直接、间接影响生物多样性与生态系统多功能性, BEMF关系具有很强的环境依赖性[66]。
4.1 自然干扰下的BEMF研究在全球变暖的背景下, 植物的物候与生长, 土壤微生物群落、物种分布等都会随之发生变化, 从而改变生态系统的水文过程、碳氮循环和生产力的产生与分配等生态过程[67], 影响BEMF关系。Valencia等[68]通过控制实验探究了气候变化对生物多样性与生态系统多功能性的影响, 发现变暖选择了较大和最具生产力的植物物种, 增加了群落的均匀度, 并导致植物功能性状多样性的减少, 这些变化增加了细菌的丰度进而增加了原生生物的丰度, 最终促进了EMF的增加。Liu等[69]通过控制实验发现, 降雨频率的减少可以降低蓝藻的相对丰度从而间接对EMF产生负面影响。在旱地生态系统中, 近几十年发生的最重要的变化之一是灌木入侵[70]。Quero等[71]探究了灌木入侵对半干旱的地中海草地和灌丛EMF的影响, 结果表明, 入侵的萌芽灌木通过增加入侵灌木丛物种多样性增强EMF。Valencia等[34]利用相同数据进一步表明入侵的萌芽灌木通过增加灌木丛的大小和叶性状功能多样性提高EMF。
4.2 人为干扰下的BEMF研究人类活动加剧引发的土地利用变化是导致生物多样性丧失和多重生态系统功能改变的重要驱动因素, 并会直接影响BEMF关系。刈割、围封、放牧是草原生态系统常见的土地利用方式, Li等[72]探究了内蒙古草原生态系统在3种土地利用方式下物种多样性和EMF的变化, 发现相比于刈割与围封, 放牧显著降低了物种多样性进而降低了EMF。Peco等[73]进一步发现放牧还可以通过改变植物的比叶面积和植物高度性状间接影响EMF, 但在宏观尺度, 食草动物通过动物粪便和尿液沉积在栖息地之间转移养分可能是影响EMF的重要机制。对于农田生态系统, Luo等[40]发现长期施肥会增加微生物群落的系统发育多样性, 从而增加EMF, Chen等[47]也发现有机肥的长期使用可以增强细菌、真菌群落的多样性以及土壤EMF。
综上, 自然干扰与人为干扰都可以显著地影响生物多样性与EMF之间的关系。随着全球变化的近一步加剧, 生态系统会经历多项极端气候事件从而加剧生物多样性与生态系统多功能性的降低, 影响生态安全。人类活动作为全球变化的主要驱动因素[74], 显著地影响极端气候事件的发生和生物多样性的丧失, 并最终改变生态系统多功能性。合理的人类活动如适度的放牧等可以缓解生物多样性的丧失, 提高生态系统多功能性[72]。因此, 未来应加强人为干预缓解极端气候、生物多样性丧失等全球变化对生态系统多功能性带来的不利影响方面的研究, 提高模拟与预测能力, 为生态系统管理提供科学依据。
5 EMF测度方法及其对BEMF关系的影响近15年的BEMF研究中, 学者就EMF度量问题提出了多种方法。徐炜等[75]对多种方法进行了总结, 近年来针对生态系统功能加权、功能权衡以及不同测度方法无法比较等问题, 相继又发展了新的表征方法(表 1)。在大多数研究中, 使用不同的测度方法均得出“随着所考虑的生态系统功能数量的增加, 需要更多的物种来维持生态系统的整体功能”这一结论[16-17, 22]。然而, 最近的两项研究对这一结论提出了质疑。Gamfeldt与Roger[44]通过模型模拟认为, 生物多样性通过对单个功能的非累加效应影响EMF的水平, 并且对EMF的影响等于对单个功能的平均影响, 也就是说BEMF关系不会随着所考虑的功能数量而改变。这一结论在Fanin等[45]的生物多样性丧失控制实验中得到了验证, 他们发现无论植物或真菌物种多样性如何丧失, BEMF关系相对不变。通过上述研究的对比发现, 可能是EMF测度方法的不同导致得出了不同的结论(表 1)。
| 方法 Methods |
公式/步骤 Formulas/Programs |
参数 Parameters |
优点 Advantages |
缺点 Disadvantages |
BEMF关系 BEMF relationship |
| 单功能法[76] Single function |
使用一般线性模型对生物多样性与EMF进行回归 | - | 直观地了解EMF随生物多样性变化的趋势 | 1.无法定量EMF;2.易受到抽样效应和互补效应的影响 | 本质是生物多样性对生态系统功能的影响, 结果可能正相关, 可能负相关, 可能无关。 |
| 周转法[8] Turnover |
1.确定影响每个生态系统功能的最小物种集合; 2.计算物种重叠度:  ; ;3.计算维持功能所需的平均物种数SE= 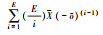
|
|Ei|、|Ej|分别表示对功能i、j有贡献的物种数目, |Ei∩Ej表示对功能i和j都有贡献的物种数目, 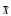 表示影响单个生态系统功能的平均物种数目, 表示影响单个生态系统功能的平均物种数目,  表示影响每对功能的物种集合的平均重叠率 表示影响每对功能的物种集合的平均重叠率 |
1.确定影响每个生态系统功能的物种集合;2.确定是否不同的物种影响不同的功能 | 1.无法定量EMF;2.数据质量要求高;3.没有考虑功能之间的权衡和相关关系;4.分析步骤繁琐 | 随着所考虑的生态系统功能数量的增加, 需要更多的物种来维持生态系统的整体功能 |
| 均值法[16, 77] Averaging |
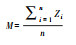
|
Zi表示生态系统功能指标i的标准化处理, n为生态系统功能指标数量。 | 定量EMF | 难以区分由相似水平的几个功能得到的EMF指数和由部分较高、部分较低功能得到的EMF指数 | 1.正相关[16];2.不相关[44] |
| 单阈值法[78] Single threshold |
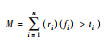
|
fi表示生态系统功能i, ti表示阈值, ri判断函数, 表示fi>ti时取1, n为生态系统功能数量 | 得到超过阈值数的物种组合 | 1.阈值的选择具有很大的主观性;2.缺少统一的标准;3.没有考虑功能之间的权衡与交互 | 正相关[10] |
| 多阈值法[79] Multiple-threshold |
1.设定0-100%的多个阈值; 2. 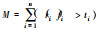
|
同单阈值法 | 1.不需要考虑阈值设定;2.蕴含信息量大;3.不同实验结果可以比较 | 1.指标较多, 流程繁琐;2. 没有考虑功能之间的权衡与交互 | 1.一般为正相关, 但在超过一定阈值后, 可能为负相关[55];2.原始数据z-标准化, 生物多样性与生态系统多功能性无关[46] |
| 多元模型法[80] Multivariate model |
1.根据公式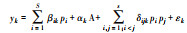
进行模型拟合, 确定最优模型;2.比较不同模型的系数β分析单作时的多功能性;3.比较不同模型的系数δ分析混种时的多功能性 |
yk表示功能k, βik表示物种i单作时, 功能k的预期表现, δijk表示物种i、j的交互作用, pipj分别表示物种i、j的相对多度。 | 1.同时评估EMF的驱动因素及其相对重要性;2可扩展到多时空尺度的EMF分析 | 1.只适合于功能数较少的研究;2.物种数量较多, 会产生大量的种间交互系数;3.不适合于相对丰度相等的实验;4.没有考虑功能的权重 | 物种均匀度与生态系统多功能性正相关[80] |
| 聚类多阈值法[81] Cluster-based multiple- threshold |
1.对生态系统功能进行聚类分析, 划分权重;2.对加权后的生态系统功能进行多阈值分析 | 同单阈值法 | 考虑功能的权重 | 指标较多, 流程繁琐 | 正相关或负相关[81] |
| PCA多变量法[82] PCA-based multivariate |
1.标准化原始数据;2.对标准化后数据进行PCA分析;3.检查PCA分析后相应的功能, 对于高值代表低功能的值乘以-1, 使得所有的高值代表高功能;4.通过特征值加权计算综合主成分得分 | - | 说明了功能之间的权衡关系 | 无法区分负值代表高功能和正值代表高功能的主成分加权得到的EMF指数与同为正值或负值代表高功能的主成分加权得到的EMF指数 | 正相关[82] |
| 缩放法[46] Scaling |
1.对原始数据进行标准化;2.对标准化后的数据进行求和或均值运算;3.对运算后的值再次z-标准化 | - | 可以比较使用不同方法进行的BEMF研究 | 无法克服均值法、阈值法等方法的缺点 | 正相关[46] |
| BEMF: 生物多样性与生态系统多功能性Biodiversity and ecosystem multifunctionality | |||||
针对是否EMF测度方法的差异会改变BEMF关系, 最近的一项研究进行了明确的比较[46]。Jing等[46]使用均值法、求和法(未进行PCA分析的多变量方法)以及缩放法量化EMF, 分别对模拟数据以及实测数据进行分析, 发现均值法下BEMF关系不会随着功能数量的增加而改变, 而求和法以及缩放法下BEMF斜率均随着功能数量的增加而增加。此外原始数据的标准化方法也会改变BEMF关系, 通过z-均值标准化方法得到的BEMF关系强度比最大值标准化方法高约10倍[46]。产生不同结果的原因主要取决于这些指标的基础统计数据, 而不是生物多样性对生态系统多功能性的真实影响。
综上, EMF测度方法的差异会影响BEMF关系的研究结果, 而目前的测度方法大多没有将功能之间的权衡关系纳入指标中, 生态系统功能之间的权衡可能随着功能数量的增加而增加从而抵消生物多样性对EMF的影响[46]。测度方法引起的结果差异是否是表象, 其内在的功能权衡为根本原因还有待探究, 后续应基于实地观测数据开展不同EMF测度方法的对比分析, 厘清EMF测度方法对BEMF关系的影响。
6 结论与展望在全球变化背景下, 面对不同类型的干扰, 不同维度的生物多样性在单一或多营养级中均会显著影响生态系统多功能性, 不同时空尺度的生物多样性作用于生态系统多功能性形成了BEMF关系的时空尺度效应。目前, EMF测度方法的差异是否会改变BEMF关系引起了争议[46], 繁多的测度方法使得各项研究之间难以比较。此外, 多维度、多尺度生物多样性对EMF影响综合作用研究的缺少也阻碍了BEMF关系的近一步探索。基于此, 本文对EMF测度方法优化方向、多维度、多尺度生物多样性对EMF的影响路径进行了展望, 提出了BEMF关系研究的通用分析框架, 旨在促进BEMF关系的深入探究, 为生物多样性保护与生态系统可持续发展提供理论指导。
6.1 发展优化EMF测度方法EMF测度方法对于BEMF研究至关重要, 然而至今没有一个统一或最佳的方法来评估EMF。最近的研究表明EMF测度方法的差异会改变BEMF关系, 基于各种统计方法造成的差异并不能反映生物多样性对生态系统多功能性的内在机理[46]。未来研究应该深刻理解各种测度方法背后的统计基础, 进行各种方法之间的比较, 深入剖析方法差异改变BEMF关系的内在机理。此外, 虽然多种测度方法可以供研究人员适用不同的研究目的, 但却不利于不同研究之间的比较, 无法形成统一的认知, 后续研究应该基于公开数据库, 加强不同方法研究结果的对比分析, 同时, 近一步开发适用不同研究目的的通用的EMF测度方法。
6.2 探究多维度、多尺度生物多样性对EMF的影响路径目前对于生物多样性的不同维度(尺度)如何影响EMF的研究多集中于同一尺度下不同维度BEMF关系或同一维度下不同尺度BEMF关系探究, 鲜有学者探究不同维度、不同尺度生物多样性的对EMF影响的综合作用。在全球变化加剧的情形下, 面对环境胁迫与人类活动, 厘清不同维度、不同尺度的生物多样性如何相互影响综合作用于EMF, 对于合理配置资源以有效提高EMF, 实现生态系统可持续发展至关重要。未来应着手探究多维度、多尺度生物多样性对EMF的影响路径, 为阐明不同维度、不同尺度的生物多样性如何相互影响综合作用于EMF提供科学理论指导。
6.3 规范BEMF关系研究范式目前关于BEMF关系的研究主要集中在区域小尺度, 并且对于生物多样性维度的选择也主要集中在物种多样性, 关于时间遗留效应对于BEMF关系影响的研究更是缺乏。时空尺度与生物多样性维度选择的局限性阻碍了BEMF关系的深化研究。在此建立通用的BEMF分析框架(图 2)以规范BEMF关系研究范式, 为BEMF研究提供科学理论指导。

|
| 图 2 BEMF研究分析框架 Fig. 2 Analysis framework of BEMF research BEMF: 生物多样性与生态系统多功能性 Biodiversity and ecosystem multifunctionality |
未来应该扩大研究的空间尺度、延长野外观测周期、丰富群落调查内容、选择具有代表性的功能指标构建长时间尺度、多空间尺度、多生物多样性维度的BEMF数据库。此外, 目前对于BEMF关系研究的方法较为单一, 多为一般线性模型和结构方程模型[16, 19, 83], 一般线性模型只能定性判断生物多样性与生态系统多功能性的关系, 而结构方程模型对数据质量要求较高, 方法的局限性也阻碍了深入理解生物多样性对生态系统多功能性影响的内在机理。因此, 未来的研究也应该引入新的研究方法, 例如运用贝叶斯统计以解决数据量较少难以分析的问题, 例如运用随机森林、网络分析等, 比较生物多样性属性对于EMF影响的相对重要性, 深入剖析BEMF关系的内在机理, 而不能只停留于表面的定性关系。总之, 未来的BEMF关系研究应该构建统一的BEMF数据库从而可以对比各项研究深入剖析生物多样性对生态系统多功能性影响的内在机理。
| [1] |
Cardinale B J, Duffy J E, Gonzalez A, Hooper D U, Perrings C, Venail P, Narwani A, Mace G M, Tilman D, Wardle D A, Kinzig A P, Daily G C, Loreau M, Grace J B, Larigauderie A, Srivastava D S, Naeem S. Biodiversity loss and its impact on humanity. Nature, 2012, 486(7401): 59-67. DOI:10.1038/nature11148 |
| [2] |
Tilman D, Isbell F, Cowles J M. Biodiversity and ecosystem functioning. Annual Review of Ecology, Evolution, and Systematics, 2014, 45: 471-493. DOI:10.1146/annurev-ecolsys-120213-091917 |
| [3] |
蒋志刚, 马克平, 韩兴国. 保护生物学. 杭州: 浙江科学技术出版社, 1997, 1-3. |
| [4] |
Jax K. Function and "functioning" in ecology: what does it mean?. Oikos, 2005, 111(3): 641-648. DOI:10.1111/j.1600-0706.2005.13851.x |
| [5] |
Hector A, Hooper R. Ecology. Darwin and the first ecological experiment. Science, 2002, 295(5555): 639-640. DOI:10.1126/science.1064815 |
| [6] |
Wood S A, Bradford M A, Gilbert J A, McGuire K L, Palm C A, Tully K L, Zhou J Z, Naeem S. Agricultural intensification and the functional capacity of soil microbes on smallholder African farms. Journal of Applied Ecology, 2015, 52(3): 744-752. DOI:10.1111/1365-2664.12416 |
| [7] |
Garland G, Banerjee S, Edlinger A, Oliveira E M, Herzog C, Wittwer R, Philippot L, Maestre F T, van der Heijden M G A. A closer look at the functions behind ecosystem multifunctionality: a review. Journal of Ecology, 2021, 109(2): 600-613. DOI:10.1111/1365-2745.13511 |
| [8] |
Hector A, Bagchi R. Biodiversity and ecosystem multifunctionality. Nature, 2007, 448(7150): 188-190. DOI:10.1038/nature05947 |
| [9] |
Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301. DOI:10.1890/07-1206.1 |
| [10] |
Zavaleta E S, Pasari J R, Hulvey K B, Tilman G D. Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(4): 1443-1446. DOI:10.1073/pnas.0906829107 |
| [11] |
Sanderson M A, Skinner R H, Barker D J, Edwards G R, Tracy B F, Wedin D A. Plant species diversity and management of temperate forage and grazing land ecosystems. Crop Science, 2004, 44(4): 1132-1144. DOI:10.2135/cropsci2004.1132 |
| [12] |
He J Z, Ge Y, Xu Z H, Chen C R. Linking soil bacterial diversity to ecosystem multifunctionality using backward-elimination boosted trees analysis. Journal of Soils and Sediments, 2009, 9(6): 547-554. DOI:10.1007/s11368-009-0120-y |
| [13] |
Isbell F, Calcagno V, Hector A, Connolly J, Harpole W S, Reich P B, Scherer-Lorenzen M, Schmid B, Tilman D, van Ruijven J, Weigelt A, Wilsey B J, Zavaleta E S, Loreau M. High plant diversity is needed to maintain ecosystem services. Nature, 2011, 477(7363): 199-202. DOI:10.1038/nature10282 |
| [14] |
Stirling G, Wilsey B. Empirical relationships between species richness, evenness, and proportional diversity. The American Naturalist, 2001, 158(3): 286-299. DOI:10.1086/321317 |
| [15] |
车盈, 金光泽. 物种多样性和系统发育多样性对阔叶红松林生产力的影响. 应用生态学报, 2019, 30(7): 2241-2248. |
| [16] |
Maestre F T, Quero J L, Gotelli N J, Escudero A, Ochoa V, Delgado-Baquerizo M, García-Gómez M, Bowker M A, Soliveres S, Escolar C, García-Palacios P, Berdugo M, Valencia E, Gozalo B, Gallardo A, Aguilera L, Arredondo T, Blones J, Boeken B, Bran D, Conceição A A, Cabrera O, Chaieb M, Derak M, Eldridge D J, Espinosa C I, Florentino A, Gaitán J, Gatica M G, Ghiloufi W, Gómez-González S, Gutiérrez J R, Hernández R M, Huang X W, Huber-Sannwald E, Jankju M, Miriti M, Monerris J, Mau R L, Morici E, Naseri K, Ospina A, Polo V, Prina A, Pucheta E, Ramírez-Collantes D A, Romão R, Tighe M, Torres-Díaz C, Val J, Veiga J P, Wang D L, Zaady E. Plant species richness and ecosystem multifunctionality in global drylands. Science, 2012, 335(6065): 214-218. DOI:10.1126/science.1215442 |
| [17] |
Perkins D M, Bailey R A, Dossena M, Gamfeldt L, Reiss J, Trimmer M, Woodward G. Higher biodiversity is required to sustain multiple ecosystem processes across temperature regimes. Global Change Biology, 2015, 21(1): 396-406. DOI:10.1111/gcb.12688 |
| [18] |
Slade E M, Kirwan L, Bell T, Philipson C D, Lewis O T, Roslin T. The importance of species identity and interactions for multifunctionality depends on how ecosystem functions are valued. Ecology, 2017, 98(10): 2626-2639. DOI:10.1002/ecy.1954 |
| [19] |
Antiqueira P A P, Petchey O L, Romero G Q. Warming and top predator loss drive ecosystem multifunctionality. Ecology Letters, 2018, 21(1): 72-82. DOI:10.1111/ele.12873 |
| [20] |
Maestre F T, Castillo-Monroy A P, Bowker M A, Ochoa-Hueso R. Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern. Journal of Ecology, 2012, 100(2): 317-330. DOI:10.1111/j.1365-2745.2011.01918.x |
| [21] |
Wagg C, Bender S F, Widmer F, van der Heijden M G A. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(14): 5266-5270. DOI:10.1073/pnas.1320054111 |
| [22] |
Bastida F, Torres I F, Moreno J L, Baldrian P, Ondoño S, Ruiz-Navarro A, Hernández T, Richnow H H, Starke R, García C, Jehmlich N. The active microbial diversity drives ecosystem multifunctionality and is physiologically related to carbon availability in Mediterranean semi-arid soils. Molecular Ecology, 2016, 25(18): 4660-4673. DOI:10.1111/mec.13783 |
| [23] |
Li J, Delgado-Baquerizo M, Wang J T, Hu H W, Cai Z J, Zhu Y N, Singh B K. Fungal richness contributes to multifunctionality in boreal forest soil. Soil Biology and Biochemistry, 2019, 136: 107526. DOI:10.1016/j.soilbio.2019.107526 |
| [24] |
Wagg C, Schlaeppi K, Banerjee S, Kuramae E E, van der Heijden M G A. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. Nature Communications, 2019, 10(1): 4841. DOI:10.1038/s41467-019-12798-y |
| [25] |
江小雷, 张卫国. 功能多样性及其研究方法. 生态学报, 2010, 30(10): 2766-2773. |
| [26] |
Tilman D. Functional diversity//Levin S A. Encyclopedia of Biodiversity. Amsterdam: Academic Press, 2001: 109-120.
|
| [27] |
Grime J P. Benefits of plant diversity to ecosystems: immediate, filter and founder effects. Journal of Ecology, 1998, 86(6): 902-910. DOI:10.1046/j.1365-2745.1998.00306.x |
| [28] |
Tilman D, Lehman C L, Thomson K T. Plant diversity and ecosystem productivity: theoretical considerations. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(5): 1857-1861. DOI:10.1073/pnas.94.5.1857 |
| [29] |
Garnier E, Cortez J, Billès G, Navas M L, Roumet C, Debussche M, Laurent G, Blanchard A, Aubry D, Bellmann A, Neill C, Toussaint J P. Plant functional markers capture ecosystem properties during secondary succession. Ecology, 2004, 85(9): 2630-2637. DOI:10.1890/03-0799 |
| [30] |
Mouillot D, Villéger S, Scherer-Lorenzen M, Mason N W H. Functional structure of biological communities predicts ecosystem multifunctionality. PLoS One, 2011, 6(3): e17476. DOI:10.1371/journal.pone.0017476 |
| [31] |
Lohbeck M, Bongers F, Martinez-Ramos M, Poorter L. The importance of biodiversity and dominance for multiple ecosystem functions in a human-modified tropical landscape. Ecology, 2016, 97(10): 2772-2779. DOI:10.1002/ecy.1499 |
| [32] |
Gross N, Le Bagousse-Pinguet Y, Liancourt P, Berdugo M, Gotelli N J, Maestre F T. Functional trait diversity maximizes ecosystem multifunctionality. Nature Ecology & Evolution, 2017, 1(5): 0132. |
| [33] |
Huang X B, Su J R, Li S F, Liu W D, Lang X D. Functional diversity drives ecosystem multifunctionality in a Pinus yunnanensis natural secondary forest. Scientific Reports, 2019, 9(1): 6979. DOI:10.1038/s41598-019-43475-1 |
| [34] |
Valencia E, Maestre F T, Le Bagousse-Pinguet Y, Quero J L, Tamme R, Börger L, García-Gómez M, Gross N. Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands. New Phytologist, 2015, 206(2): 660-671. DOI:10.1111/nph.13268 |
| [35] |
Mensah S, Salako K V, Assogbadjo A, Glèlè Kakaï R, Sinsin B, Seifert T. Functional trait diversity is a stronger predictor of multifunctionality than dominance: evidence from an Afromontane forest in South Africa. Ecological Indicators, 2020, 115: 106415. DOI:10.1016/j.ecolind.2020.106415 |
| [36] |
Fry E L, Savage J, Hall A L, Oakley S, Pritchard W J, Ostle N J, Pywell R F, Bullock J M, Bardgett R D. Soil multifunctionality and drought resistance are determined by plant structural traits in restoring grassland. Ecology, 2018, 99(10): 2260-2271. DOI:10.1002/ecy.2437 |
| [37] |
Srivastava D S, Cadotte M W, MacDonald A A M, Marushia R G, Mirotchnick N. Phylogenetic diversity and the functioning of ecosystems. Ecology Letters, 2012, 15(7): 637-648. DOI:10.1111/j.1461-0248.2012.01795.x |
| [38] |
Narwani A, Matthews B, Fox J, Venail P. Using phylogenetics in community assembly and ecosystem functioning research. Functional Ecology, 2015, 29(5): 589-591. DOI:10.1111/1365-2435.12431 |
| [39] |
Tucker C M, Cadotte M W, Carvalho S B, Davies T J, Ferrier S, Fritz S A, Grenyer R, Helmus M R, Jin L S, Mooers A O, Pavoine S, Purschke O, Redding D W, Rosauer D F, Winter M, Mazel F. A guide to phylogenetic metrics for conservation, community ecology and macroecology. Biological Reviews, 2017, 92(2): 698-715. DOI:10.1111/brv.12252 |
| [40] |
Luo G W, Rensing C, Chen H, Liu M Q, Wang M, Guo S W, Ling N, Shen Q R. Deciphering the associations between soil microbial diversity and ecosystem multifunctionality driven by long-term fertilization management. Functional Ecology, 2018, 32(4): 1103-1116. DOI:10.1111/1365-2435.13039 |
| [41] |
Li H, Huo D, Wang W B, Chen Y X, Cheng X L, Yu G L, Li R H. Multifunctionality of biocrusts is positively predicted by network topologies consistent with interspecies facilitation. Molecular Ecology, 2020, 29(8): 1560-1573. DOI:10.1111/mec.15424 |
| [42] |
Le Bagousse-Pinguet Y, Soliveres S, Gross N, Torices R, Berdugo M, Maestre F T. Phylogenetic, functional, and taxonomic richness have both positive and negative effects on ecosystem multifunctionality. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(17): 8419-8424. DOI:10.1073/pnas.1815727116 |
| [43] |
Yuan Z Q, Ali A, Ruiz-Benito P, Jucker T, Mori A S, Wang S P, Zhang X K, Li H, Hao Z Q, Wang X G, Loreau M, Allan E. Above-and below-ground biodiversity jointly regulate temperate forest multifunctionality along a local-scale environmental gradient. Journal of Ecology, 2020, 108(5): 2012-2024. DOI:10.1111/1365-2745.13378 |
| [44] |
Gamfeldt L, Roger F. Revisiting the biodiversity-ecosystem multifunctionality relationship. Nature Ecology & Evolution, 2017, 1(7): 0168. |
| [45] |
Fanin N, Gundale M J, Farrell M, Ciobanu M, Baldock J A, Nilsson M C, Kardol P, Wardle D A. Consistent effects of biodiversity loss on multifunctionality across contrasting ecosystems. Nature Ecology & Evolution, 2018, 2(2): 269-278. |
| [46] |
Jing X, Prager C M, Classen A T, Maestre F T, He J S, Sanders N J. Variation in the methods leads to variation in the interpretation of biodiversity-ecosystem multifunctionality relationships. Journal of Plant Ecology, 2020, 13(4): 431-441. DOI:10.1093/jpe/rtaa031 |
| [47] |
Chen Q L, Ding J, Zhu D, Hu H W, Delgado-Baquerizo M, Ma Y B, He J Z, Zhu Y G. Rare microbial taxa as the major drivers of ecosystem multifunctionality in long-term fertilized soils. Soil Biology and Biochemistry, 2020, 141: 107686. DOI:10.1016/j.soilbio.2019.107686 |
| [48] |
Liu C C, Li Y, Yan P, He N P. How to improve the predictions of plant functional traits on ecosystem functioning?. Frontiers in Plant Science, 2021, 12: 622260. DOI:10.3389/fpls.2021.622260 |
| [49] |
Naeem S, Hahn D R, Schuurman G. Producer-decomposer co-dependency influences biodiversity effects. Nature, 2000, 403(6771): 762-764. DOI:10.1038/35001568 |
| [50] |
Duffy J E, Cardinale B J, France K E, Mcintyre P B, Thébault E, Loreau M. The functional role of biodiversity in ecosystems: incorporating trophic complexity. Ecology Letters, 2007, 10(6): 522-538. DOI:10.1111/j.1461-0248.2007.01037.x |
| [51] |
Soliveres S, van der Plas F, Manning P, Prati D, Gossner M M, Renner S C, Alt F, Arndt H, Baumgartner V, Binkenstein J, Birkhofer K, Blaser S, Blüthgen N, Boch S, Böhm S, Börschig C, Buscot F, Diekötter T, Heinze J, Hölzel N, Jung K, Klaus V H, Kleinebecker T, Klemmer S, Krauss J, Lange M, Morris E K, Müller J, Oelmann Y, Overmann J, Pašalić E, Rillig M C, Schaefer H M, Schloter M, Schmitt B, Schöning I, Schrumpf M, Sikorski J, Socher S A, Solly E F, Sonnemann I, Sorkau E, Steckel J, Steffan-Dewenter I, Stempfhuber B, Tschapka M, Türke M, Venter P C, Weiner C N, Weisser W W, Werner M, Westphal C, Wilcke W, Wolters V, Wubet T, Wurst S, Fischer M, Allan E. Biodiversity at multiple trophic levels is needed for ecosystem multifunctionality. Nature, 2016, 536(7617): 456-459. DOI:10.1038/nature19092 |
| [52] |
Eisenhauer N, Reich P B, Isbell F. Decomposer diversity and identity influence plant diversity effects on ecosystem functioning. Ecology, 2012, 93(10): 2227-2240. DOI:10.1890/11-2266.1 |
| [53] |
Jing X, Sanders N J, Shi Y, Chu H Y, Classen A T, Zhao K, Chen L T, Shi Y, Jiang Y X, He J S. The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate. Nature Communications, 2015, 6: 8159. DOI:10.1038/ncomms9159 |
| [54] |
Schuldt A, Assmann T, Brezzi M, Buscot F, Eichenberg D, Gutknecht J, Härdtle W, He J S, Klein A M, Kühn P, Liu X J, Ma K P, Niklaus P A, Pietsch K A, Purahong W, Scherer-Lorenzen M, Schmid B, Scholten T, Staab M, Tang Z Y, Trogisch S, von Oheimb G, Wirth C, Wubet T, Zhu C D, Bruelheide H. Biodiversity across trophic levels drives multifunctionality in highly diverse forests. Nature Communications, 2018, 9(1): 2989. DOI:10.1038/s41467-018-05421-z |
| [55] |
Lefcheck J S, Byrnes J E K, Isbell F, Gamfeldt L, Griffin J N, Eisenhauer N, Hensel M J S, Hector A, Cardinale B J, Duffy J E. Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats. Nature Communications, 2015, 6: 6936. DOI:10.1038/ncomms7936 |
| [56] |
Ye J S, Delgado-Baquerizo M, Soliveres S, Maestre F T. Multifunctionality debt in global drylands linked to past biome and climate. Global Change Biology, 2019, 25(6): 2152-2161. DOI:10.1111/gcb.14631 |
| [57] |
Lucas-Borja M E, Delgado-Baquerizo M. Plant diversity and soil stoichiometry regulates the changes in multifunctionality during pine temperate forest secondary succession. Science of the Total Environment, 2019, 697: 134204. DOI:10.1016/j.scitotenv.2019.134204 |
| [58] |
Pringle R M, Doak D F, Brody A K, Jocqué R, Palmer T M. Spatial pattern enhances ecosystem functioning in an african savanna. PLoS Biology, 2010, 8(5): e1000377. DOI:10.1371/journal.pbio.1000377 |
| [59] |
Seahra S E, Yurkonis K A, Newman J A. Species patch size at seeding affects diversity and productivity responses in establishing grasslands. Journal of Ecology, 2016, 104(2): 479-486. DOI:10.1111/1365-2745.12514 |
| [60] |
Pasari J R, Levi T, Zavaleta E S, Tilman D. Several scales of biodiversity affect ecosystem multifunctionality. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(25): 10219-10222. DOI:10.1073/pnas.1220333110 |
| [61] |
Grman E, Zirbel C R, Bassett T, Brudvig L A. Ecosystem multifunctionality increases with beta diversity in restored prairies. Oecologia, 2018, 188(3): 837-848. DOI:10.1007/s00442-018-4248-6 |
| [62] |
van der Plas F, Manning P, Soliveres S, Allan E, Scherer-Lorenzen M, Verheyen K, Wirth C, Zavala M A, Ampoorter E, Baeten L, Barbaro L, Bauhus J, Benavides R, Benneter A, Bonal D, Bouriaud O, Bruelheide H, Bussotti F, Carnol M, Castagneyrol B, Charbonnier Y, Coomes D A, Coppi A, Bastias C C, Dawud S M, De Wandeler H, Domisch T, Finér L, Gessler A, Granier A, Grossiord C, Guyot V, Hättenschwiler S, Jactel H, Jaroszewicz B, Joly F X, Jucker T, Koricheva J, Milligan H, Mueller S, Muys B, Nguyen D, Pollastrini M, Ratcliffe S, Raulund-Rasmussen K, Selvi F, Stenlid J, Valladares F, Vesterdal L, Zielinski D, Fischer M. Biotic homogenization can decrease landscape-scale forest multifunctionality. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(13): 3557-3562. DOI:10.1073/pnas.1517903113 |
| [63] |
Hautier Y, Isbell F, Borer E T, Seabloom E W, Harpole W S, Lind E M, Macdougall A S, Stevens C J, Adler P B, Alberti J, Bakker J D, Brudvig L A, Buckley Y M, Cadotte M, Caldeira M C, Chaneton E J, Chu C J, Daleo P, Dickman C R, Dwyer J M, Eskelinen A, Fay P A, Firn J, Hagenah N, Hillebrand H, Iribarne O, Kirkman K P, Knops J M H, La Pierre K J, McCulley R L, Morgan J W, Pärtel M, Pascual J, Price J N, Prober S M, Risch A C, Sankaran M, Schuetz M, Standish R J, Virtanen R, Wardle G M, Yahdjian L, Hector A. Local loss and spatial homogenization of plant diversity reduce ecosystem multifunctionality. Nature Ecology & Evolution, 2018, 2(1): 50-56. |
| [64] |
Yan Y Z, Zhang Q, Buyantuev A, Liu Q F, Niu J M. Plant functional β diversity is an important mediator of effects of aridity on soil multifunctionality. Science of the Total Environment, 2020, 726: 138529. DOI:10.1016/j.scitotenv.2020.138529 |
| [65] |
Seidl R, Rammer W, Spies T A. Disturbance legacies increase the resilience of forest ecosystem structure, composition, and functioning. Ecological Applications, 2014, 24(8): 2063-2077. DOI:10.1890/14-0255.1 |
| [66] |
Ratcliffe S, Wirth C, Jucker T, van der Plas F, Scherer-Lorenzen M, Verheyen K, Allan E, Benavides R, Bruelheide H, Ohse B, Paquette A, Ampoorter E, Bastias C C, Bauhus J, Bonal D, Bouriaud O, Bussotti F, Carnol M, Castagneyrol B, Chećko E, Dawud S M, de Wandeler H, Domisch T, Finér L, Fischer M, Fotelli M, Gessler A, Granier A, Grossiord C, Guyot V, Haase J, Hättenschwiler S, Jactel H, Jaroszewicz B, Joly F X, Kambach S, Kolb S, Koricheva J, Liebersgesell M, Milligan H, Müller S, Muys B, Nguyen D, Nock C, Pollastrini M, Purschke O, Radoglou K, Raulund-Rasmussen K, Roger F, Ruiz-Benito P, Seidl R, Selvi F, Seiferling I, Stenlid J, Valladares F, Vesterdal L, Baeten L. Biodiversity and ecosystem functioning relations in European forests depend on environmental context. Ecology Letters, 2017, 20(11): 1414-1426. DOI:10.1111/ele.12849 |
| [67] |
徐满厚, 薛娴. 气候变暖对陆地植被-土壤生态系统的影响研究. 生命科学, 2012, 24(5): 492-500. |
| [68] |
Valencia E, Gross N, Quero J L, Carmona C P, Ochoa V, Gozalo B, Delgado-Baquerizo M, Dumack K, Hamonts K, Singh B K, Bonkowski M, Maestre F T. Cascading effects from plants to soil microorganisms explain how plant species richness and simulated climate change affect soil multifunctionality. Global Change Biology, 2018, 24(12): 5642-5654. DOI:10.1111/gcb.14440 |
| [69] |
Liu Y R, Delgado-Baquerizo M, Trivedi P, He J Z, Wang J T, Singh B K. Identity of biocrust species and microbial communities drive the response of soil multifunctionality to simulated global change. Soil Biology and Biochemistry, 2017, 107: 208-217. DOI:10.1016/j.soilbio.2016.12.003 |
| [70] |
Eldridge D J, Bowker M A, Maestre F T, Roger E, Reynolds J F, Whitford W G. Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecology Letters, 2011, 14(7): 709-722. DOI:10.1111/j.1461-0248.2011.01630.x |
| [71] |
Quero J L, Maestre F T, Ochoa V, García-Gómez M, Delgado-Baquerizo M. On the importance of shrub encroachment by sprouters, climate, species richness and anthropic factors for ecosystem multifunctionality in semi-arid Mediterranean ecosystems. Ecosystems, 2013, 16(7): 1248-1261. DOI:10.1007/s10021-013-9683-y |
| [72] |
Li J P, Zheng Z R, Xie H T, Zhao N X, Gao Y B. Heterogeneous microcommunities and ecosystem multifunctionality in seminatural grasslands under three management modes. Ecology and Evolution, 2017, 7(1): 14-25. DOI:10.1002/ece3.2604 |
| [73] |
Peco B, Navarro E, Carmona C P, Medina N G, Marques M J. Effects of grazing abandonment on soil multifunctionality: the role of plant functional traits. Agriculture, Ecosystems & Environment, 2017, 249: 215-225. |
| [74] |
Karl T R, Trenberth K E. Modern global climate change. Science, 2003, 302(5651): 1719-1723. DOI:10.1126/science.1090228 |
| [75] |
徐炜, 井新, 马志远, 贺金生. 生态系统多功能性的测度方法. 生物多样性, 2016, 24(1): 72-84. |
| [76] |
Duffy J E, Richardson J P, Canuel E A. Grazer diversity effects on ecosystem functioning in seagrass beds. Ecology Letters, 2003, 6(7): 637-645. DOI:10.1046/j.1461-0248.2003.00474.x |
| [77] |
Hooper D U, Vitousek P M. Effects of plant composition and diversity on nutrient cycling. Ecological Monographs, 1998, 68(1): 121-149. DOI:10.1890/0012-9615(1998)068[0121:EOPCAD]2.0.CO;2 |
| [78] |
Gamfeldt L, Hillebrand H, Jonsson P R. Multiple functions increase the importance of biodiversity for overall ecosystem functioning. Ecology, 2008, 89(5): 1223-1231. DOI:10.1890/06-2091.1 |
| [79] |
Byrnes J E K, Gamfeldt L, Isbell F, Lefcheck J S, Griffin J N, Hector A, Cardinale B J, Hooper D U, Dee L E, Duffy J E. Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions. Methods in Ecology and Evolution, 2014, 5(2): 111-124. DOI:10.1111/2041-210X.12143 |
| [80] |
Dooley Á, Isbell F, Kirwan L, Connolly J, Finn J A, Brophy C, Buckley Y. Testing the effects of diversity on ecosystem multifunctionality using a multivariate model. Ecology Letters, 2015, 18(11): 1242-1251. DOI:10.1111/ele.12504 |
| [81] |
Manning P, van der Plas F, Soliveres S, Allan E, Maestre F T, Mace G, Whittingham M J, Fischer M. Redefining ecosystem multifunctionality. Nature Ecology & Evolution, 2018, 2(3): 427-436. |
| [82] |
Meyer S T, Ptacnik R, Hillebrand H, Bessler H, Buchmann N, Ebeling A, Eisenhauer N, Engels C, Fischer M, Halle S, Klein A M, Oelmann Y, Roscher C, Rottstock T, Scherber C, Scheu S, Schmid B, Schulze E D, Temperton V M, Tscharntke T, Voigt W, Weigelt A, Wilcke W, Weisser W W. Biodiversity-multifunctionality relationships depend on identity and number of measured functions. Nature Ecology & Evolution, 2018, 2(1): 44-49. |
| [83] |
Li S F, Liu W D, Lang X D, Huang X B, Su J R. Species richness, not abundance, drives ecosystem multifunctionality in a subtropical coniferous forest. Ecological Indicators, 2021, 120: 106911. DOI:10.1016/j.ecolind.2020.106911 |