 2022, Vol. 42
2022, Vol. 42文章信息
- 王乐, 杜灵通, 马龙龙, 丹杨
- WANG Le, DU Lingtong, MA Longlong, DAN Yang
- 人工灌丛化对荒漠草原生态系统碳储量的影响
- Effects of planted shrub encroachment on carbon storage of desert steppe ecosystem
- 生态学报. 2022, 42(1): 246-254
- Acta Ecologica Sinica. 2022, 42(1): 246-254
- http://dx.doi.org/10.5846/stxb202010112593
-
文章历史
- 收稿日期: 2020-10-11
- 网络出版日期: 2021-08-16





2. 宁夏大学西北退化生态系统恢复与重建教育部重点实验室, 银川 750021;
3. 河海大学水文水资源学院, 南京 210009
2. Key Laboratory for Restoration and Reconstruction of Degraded Ecosystem in Northwest China of Ministry of Education, Ningxia University, Yinchuan 750021, China;
3. College of Hydrology and Water Resources, Hohai University, Nanjing 210009, China
“千分之四倡议”科学技术委员会的专家在《Nature》发文, 呼吁各国采取措施增加土壤碳储量以应对全球气候变化, 并指出种植固碳植物是提高土壤碳储量以应对气候变化的重要手段之一[1]。植物作为大气与陆地间CO2循环的媒介, 大约会消耗掉1/3的CO2, 最终会将10%—15%的CO2截存在土壤中[2]。同时, 植被生物量的增加也可提升土壤的碳截存能力[3], 如1970—2000年间我国的大规模造林运动使得森林碳储量增加了40%[4]。卫星遥感监测表明, 全球植被结构的变化已显著增强了陆地生态系统的碳汇能力[5];中国和印度近些年因土地利用转变而导致“地球变绿”引起全球广泛关注[6], 其中一个主要贡献是植树造林, 但有关这一植被结构变化所引起的碳循环反馈机制尚需深入研究[7]。在中国西北干旱风沙区荒漠草原上, 大量人工种植灌木植被进行防沙治沙已成为重要的生态治理措施, 这一措施引起了草地的灌丛化, 改变了区域植被结构, 也势必对生态系统碳储量产生深远影响。
灌丛化是指草原生态系统中灌木或木本植物的密度、盖度和生物量增加的现象。这一现象在干旱及半干旱区广泛发生, 不仅会致使草地生态系统结构与功能发生转变[8], 还将最终改变生态系统的碳收入[9]。国内外有关灌丛化的报道多为气候变暖或过渡放牧干扰引起的草地退化结果[10]。然而, Meta分析表明, 全球草地广泛发生的灌丛化过程对表层土壤有机碳有积极影响[11];也有文献指出灌木的入侵会导致土壤有机碳减少[12-13];由此可见, 草地生态系统碳循环对灌木入侵过程存在不同的响应机制, 需要分地区、分类型厘清。在草地灌丛化现象中, 有一种近几十年来发生在我国西北干旱区的人工灌丛化现象值得关注, 不同于草地退化引起的灌丛化自然现象, 人工灌丛化是由人为介导的灌木入侵, 其目的是进行区域防沙治沙和生态恢复。目前已有学者关注到人工灌丛化会影响草地生态系统的碳水循环过程[14], 尤其是人工灌丛化改变了地表蒸散过程和土壤水文过程[15-16], 但有关人工灌丛化如何影响生态系统碳储量的认识尚显缺乏。虽然, 有文献报道了宁夏荒漠草原灌丛化过程中的土壤有机碳变化[17], 但由于人工灌丛化过程伴随着灌木植被的快速生长入侵, 其对草地系统碳储量的影响将伴随着灌木生长过程而发生非线性变化。故掌握人工灌丛化过程中长时间序列的生态系统碳储量动态变化, 对评估区域生态治理效果具有重要参考价值。为此, 本研究以宁夏盐池县荒漠草原的人工灌丛化过程为例, 利用Logistic模型模拟灌木生长过程, 结合Biome-BGC模型模拟荒漠草原人工灌丛化过程中的碳储量变化, 以期揭示生态系统碳储量对人工灌丛化的响应规律。
1 材料与方法 1.1 研究区概况盐池县位于宁夏回族自治区东部, 37°04′—38°10′N和106°30′—107位于宁夏回之间, 总面积6769 km2, 海拔1295—1951 m。北邻毛乌素沙地, 东南接黄土高原, 地势南高北低。1958—2017年间的年平均气温为8.34 ℃, 年均降水为296.99 mm, 属于典型的温带大陆性气候。研究区土壤结构松散, 地带性土壤以灰钙土为主, 非地带性土壤主要有风沙土和草甸土等。主要物种有中间锦鸡儿(Caragana liouana)、短花针茅(Stipa breviflora)、牛枝子(Lespedeza potaninii)、猪毛蒿(Artemisia scoparia)、白草(Pennisetum centrasiaticum)等[18]。盐池属于典型的过渡地带, 地理单元从黄土丘陵向鄂尔多斯台地的过渡, 气候从半干旱区向干旱区过渡, 植被从干草原向荒漠草原过渡, 故其生态系统极其脆弱。由于草地沙化严重, 20世纪中后期开始大量种植中间锦鸡儿(俗称柠条)用以防风固沙和草原生态恢复, 这一措施造就了盐池半草原—半灌丛的特殊自然景观(图 1)。

|
| 图 1 研究区地理位置及荒漠草原上的人工灌丛景观 Fig. 1 Location of study area and landscape of planted shrub in desert steppe |
驱动Biome-BGC模型需要气象数据和其他描述性数据。气象数据包括盐池县1958—2017年的逐日最高温度、最低温度及降水量等, 从中国气象数据网(http://data.cma.cn/)获取;描述性数据包括观测站点属性、土壤属性、CO2浓度数据、植被类型及生理生态参数等(表 1)。根据Biome-BGC模型模拟需求, 分别构建了C3草本和灌木2种光合类型植被的44个参数生理生态参数, 包括叶片碳氮比、物候过程、根茎分配、冠层比叶面积、最大气孔导度等, 具体含义详见文献[19];大气CO2浓度数据来自青海省瓦里关大气本底站的观测。构建Logistic生长模型所需的柠条灌木连续生长观测数据来自参考文献[20]。
| 数据类型 Data type |
数据内容 Content of data set |
时间分辨率 Temporal resolution |
数据来源 Data sources |
| 站点属性 Physical characteristics of site |
高程、海拔和经纬度等 | — | 观测站实测 |
| 气象数据 Meteorological data |
日最高温度、最低温度和降水量 | 天 | 中国气象数据网 |
| 植物生理生态 Eco-physiological parameters of vegetation |
44个生理生态参数 | — | 本地化后的缺省值[15] |
本研究结合Logistic生长模型和Biome-BGC生态过程模型来模拟生态系统碳储量随人工灌丛化过程的变化。因荒漠草原的人工灌丛化交织着两种自然过程, 一是柠条种植后的灌木自然生长和草地退化过程, 这一过程至少需要15年;二是草本植物和柠条灌木各自的碳蓄积过程。为将这两种自然过程在模型中描述清楚, 首先利用已有的人工柠条生长观测数据, 基于灌木生长的Logistic通用方程, 确定生长模型参数, 假定以1970年种植的柠条为情景, 模拟灌木在草原上的生长过程, 律定每年单位面积内灌木与草本植物的冠层覆盖占比。然后, 结合Biome-BGC模拟的草本和灌木植被的碳蓄积过程, 换算出原始荒漠草原生态系统向灌草复合系统转变的碳储量动态, 并将其与原始荒漠草原生态系统的碳蓄积过程对比。
1.3.1 Logistic模型植物生长过程通常可利用数学模型进行描述, 其中Logistic模型是一种常用方法, 它通过“S”型曲线模拟植物生长过程, 又被称为自我抑制性方程[21]。本研究利用该模型模拟柠条灌木种植后由幼苗到成年的生长过程, 公式如下:
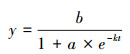
|
(1) |
式中, t为时间, y为植物生长指标, b表示一定时期内生长的上限, a是与曲线位置有关的参数, k表示内禀生长率。其中k, a, b为该方程参数, 对不同的植物类型, 需要用实测数据律定参数值。本文采用程杰等[20]发表的黄土高原区柠条逐年地径生长实测数据进行拟合, 由于本研究未区分不同的坡位状态, 故将其三种坡位的实测地径生长数据进行平均, 再采用Levenberg-Marquardt方法在Matlab软件中进行Logistics方程的拟合, 来模拟人工灌丛后柠条的生长过程, 确定地表单位面积上柠条灌木与草本的占比。
1.3.2 Biome-BGC模型Biome-BGC模型遵循物质与能量守恒定律, 即进入系统的物质和能量等于留在系统中的物质和能量加上离开系统的物质和能量[22]。该模型可模拟常绿针叶林、常绿阔叶林、落叶针叶林、落叶阔叶林、C3草本植物、C4草本植物和灌木林共7种植被类型的碳、氮、水的循环过程, 但缺少对于混合植被类型的生理生态参数描述。故本文基于BGC模型对C3草本和灌木2种植被类型的碳储量独立模拟, 结合Logistic生长模型对灌木与草本动态演变的描述, 定量计算出人工灌丛化过程中荒漠草原生态系统碳储量的变化过程。
2 结果与分析 2.1 荒漠草原人工灌丛化过程模拟 2.1.1 柠条的Logistic生长方程及生长曲线利用实测的23年柠条地径数据, 带入Logistic模型求出该地区柠条生长方程的对应参数, 并利用律定的柠条生长方程公式, 模拟出1970—2017年间盐池县柠条地径的标准生长曲线(图 2)。从图 2可以看出, 自盐池荒漠草原1970年种植柠条灌木起, 柠条地径从种植当年的0.17 cm开始以非线性增长速率快速生长, 直至1989年达到1.5 cm左右后, 其地径生长速率开始放缓, 并逐渐变得较为固定, 地径大小保持在1.5 cm上下, 即柠条进入了成年植株。模型误差分析结果表明, Logistic模型能够较好的模拟出柠条灌木的生长过程, 其模拟值与实测值之间的决定系数(R2)达到了0.96, 模拟的均方根误差(RMSE)为0.11 cm, 在柠条灌木成年后, 模拟误差明显小于地径实际测量值, 表明本研究所建立的柠条灌木Logistic生长模型具有良好的适用性。

|
| 图 2 柠条Logistic生长曲线 Fig. 2 Logistic growth curve of Caragana liouana |
基于柠条Logistic生长模型模拟出的柠条地径生长速率, 来换算其在草地上生长时的逐年冠幅面积占比, 由于柠条灌木的入侵, 草地中柠条灌丛冠幅占比逐渐增加, 而草本植物冠幅的占比随之下降, 这种变化过程为非线性变化, 与柠条生长曲线变化特征一致(图 3)。从计算出的灌木与草本占比可以看出, 以1970年开始人工种植的柠条灌木为情景, 柠条冠幅的面积占比在生长初中期快速增加。经过十几年的生长, 柠条灌木进入成年期, 其面积占比在1989年后增速放缓, 直至逐渐保持在9.0%左右的不变比例, 形成了较为稳定的人工灌草复合系统。这一结果也表明, 荒漠草原生态系统在人工灌丛化初中期, 植被结构发生快速转变, 并引起生态系统碳储量及碳循环特征由草地型向灌丛型转变, 随着柠条植株成年, 灌木和草本的结构比例稳定, 其生态系统的碳储量也将逐渐维持稳定。

|
| 图 3 灌木—草本面积占比随柠条种植年限增长的变化 Fig. 3 Changes in area percentage of shrub and grass with growth of Caragana liouana |
结合Logistic生长过程模型与Biome-BGC模型, 模拟出荒漠草原人工灌丛化过程中的逐年碳储量。本研究利用植被净初级生产力数据[23], 间接换算生态系统碳储量后验证上述方法模拟的碳储量精度。依据方精云等报道的灌丛碳密度与和草地碳密度的经验比值6.42[24], 本研究将宁夏草地植被净初级生产力数据乘以该系数转换为荒漠草原灌丛化后灌-草系统的植被净初级生产力, 再利用净初级生产力与地上植被碳储量间的转换系数0.45[24], 计算获取盐池荒漠草原人工灌丛化后的植被碳储量。然后依据该数据验证本研究模拟的植被碳储量。结果表明(图 4), 本研究模拟的植被碳储量与植被净初级生产力换算出的碳储量具有较高的相关性, 模型估算的均方根误差(RMSE)为0.15 kg/m2, 其精度可满足生态系统碳储量特征研究。

|
| 图 4 模拟植被碳储量验证 Fig. 4 Validation of simulated vegetation carbon storage NPP:净初级生产力, Net Primary Production |
近60 a荒漠草原生态系统和人工灌丛化的灌-草复合系统的碳储量变化如图 5所示, 荒漠草原生态系统的总碳储量在60 a间呈缓慢的上升趋势, 在1.9799 kg/m2至2.0743 kg/m2之间波动;而人工灌丛化的荒漠草原总碳储量受灌木生长的影响明显, 从1970年的柠条种植初年开始, 总碳储量发生抬升, 随着柠条生长发育, 总碳储量在柠条种植后的20 a中发生显著抬升, 由1970年的2.0230 kg/m2快速增加至1990年的2.3518 kg/m2, 增幅达16.25%。当柠条灌木生长到成熟龄后, 灌木与草本植被的组分比例保持稳定, 灌-草复合系统的总碳储量也保持在了稳定状态, 其总碳储量在2017年为2.4055 kg/m2, 相比1990年, 仅增加了2.28%。荒漠草原人工灌丛化引起的总碳储量变化与植被群落的演替有关, 人工灌丛入侵荒漠草原后, 植被群落由草本为主转变为以灌木为主, 这导致生态系统的碳储量特征由草地型向灌木型转变。

|
| 图 5 人工灌丛化对荒漠草原生态系统总碳储量的影响 Fig. 5 Effect of planted shrub on total carbon storage of desert steppe ecosystem |
人工灌丛化也改变了荒漠草原生态系统各碳组分的比例(表 2)。荒漠草原生态系统的植被碳储量和枯落物碳储量分别占总碳储量的2.53%和3.51%, 即地上碳储量占总碳储量的比例仅有6.04%, 而地下土壤碳储量占比却高达93.96%, 对总碳储量的贡献非常大。当荒漠草原人工灌丛化后, 由于灌木的生物量比草本大, 故植被碳储量明显增加, 其占总碳储量的比例上升到13.07%;灌木生物量的增加带来了枯落物的增加, 枯落物碳储量占比也上升到5.31%;最终导致灌-草复合系统地上碳储量占比的升高。荒漠草原人工灌丛化后, 土壤碳储量由1.9435 kg/m2增加到了1.9633 kg/m2, 略有提高。由于生态系统将大气CO2循环至土壤中并储存下来是一个漫长的过程, 故荒漠草原人工灌丛化后的近几十年中土壤碳储量增幅并不大, 但由于植被碳和枯落物碳储量的快速增加, 导致灌-草复合系统的土壤碳储量占总碳储量比例下降。以上结果表明, 荒漠草原人工灌丛化不仅增加了总碳储量, 而且改变了生态系统的碳组分结构。
| 植被碳 Vegetation carbon storage/(kg/m 2) |
枯落物碳 Litter carbon storage/(kg/m 2) |
土壤碳 Soil carbon storage/(kg/m 2) |
总碳 Total carbon storage/(kg/m 2) |
|
| 荒漠草原生态系统 Desert steppe ecosystem |
0.0524 (2.53%) |
0.0727 (3.51%) |
1.9435 (93.96%) |
2.0686 (100.00%) |
| 灌-草复合系统 Composite system of shrub and grass |
0.3144 (13.07%) |
0.1278 (5.31%) |
1.9633 (81.62%) |
2.4055 (100.00%) |
从荒漠草原人工灌丛化前后植被和枯落物碳储量的年际变化来看(图 6), 人工灌丛化对荒漠草原的地上碳储量影响较大, 从灌木种植初年开始, 二者的碳储量曲线发生明显分异。在1958—2017年间, 荒漠草原生态系统的植被碳储量由0.0435 kg/m2增长至0.0524 kg/m2, 年均增长率为0.00015 kg m-2a-1;同期枯落物碳储量由0.0620 kg/m2增长为0.0727 kg/m2, 年均增长率为0.00018 kg m-2a-1。而荒漠草原人工灌丛化后, 植被和枯落物碳储量明显出现抬升的现象, 其中植被碳储量表现出先快速增长后转缓的特征, 而枯落物碳储量的整体抬升相对较缓。灌-草复合系统的植被和枯落物碳储量在2017年分别达到0.3144 kg/m2和0.1278 kg/m2, 近60年的年均增速分别为0.0045 kg m-2a-1和0.0011 kg m-2a-1, 明显高于荒漠草原。通过对比可见, 盐池荒漠草原在种植柠条灌木的37年后, 植被碳储量由荒漠草原生态系统的0.0524 kg/m2增加至灌-草复合系统的0.3144 kg/m2, 增加了6倍;枯落物碳储量由荒漠草原生态系统的0.0727 kg/m2增加至灌-草复合系统的0.1278 kg/m2, 增加约1.76倍。

|
| 图 6 人工灌丛化对荒漠草原生态系统植被和枯落物碳储量的影响 Fig. 6 Effect of planted shrub on vegetation and litter carbon storage of desert steppe ecosystem |
从人工灌丛化前后荒漠草原生态系统土壤碳储量的年际变化来看(图 7), 人工灌丛化对荒漠草原的土壤碳储量也产生了一定的影响。在荒漠草原生态系统中, 土壤碳储量在60 a间总体呈现波动上升趋势, 从1958年的1.8728 kg/m2增长至2017年的1.9436 kg/m2, 60 a间土壤碳储量的最高值出现在2015年, 达到了1.9489 kg/m2, 此后两年有微弱的回落, 这与气候变化导致的土壤碳储量年际间微弱波动有关。荒漠草原人工灌丛化后, 其土壤碳储量的增速加强, 到2017年增加到1.9633 kg/m2, 是同期原始荒漠草原生态系统土壤碳储量的1.01倍。以上结果可知, 荒漠草原人工灌丛化会增强生态系统向土壤的碳输入, 但对地下土壤碳储量的影响不及对地上植被和枯落物碳储量的影响大。这与土壤碳的累积输入过程有关, 土壤碳是通过生态系统的生物活动缓慢转化并最终累积于土壤之中, 尽管人工灌丛入侵荒漠草原后有较强的地上生物生产能力, 能快速产生较多的植被和枯落物碳储量, 但这些生物碳转化到土壤之中尚需更长的时间。

|
| 图 7 人工灌丛化对荒漠草原生态系统土壤碳储量的影响 Fig. 7 Effect of planted shrub on soil carbon storage of desert steppe ecosystem |
荒漠草原人工灌丛化是一个植被类型受人为干扰而快速转变的复杂过程, 本研究将柠条灌木和原始地被层草本分开成两种植被类型, 即分别看做一个独立的C3草地和一个独立的灌丛, 这两种植被类型在样地尺度上组合, 形成了灌-草复合系统。本研究首先利用Biome-BGC模型分别模拟C3草地和灌丛生态系统碳储量序列, 然后基于Logistic生长曲线模拟出荒漠草原柠条灌木的生长过程, 构建单位面积上的灌丛和草地动态变化比例, 最终按面积占比对模拟的C3草地和灌丛的碳储量序列进行重构, 得到60 a间人工灌丛化过程中荒漠草原生态系统碳储量的动态变化规律。这一方法不仅解决了Biome-BGC模型无法模拟植被类型转换过程中的碳储量问题, 也使得种植人工灌丛化的荒漠草原碳储量模拟更逼近现实情况, 为今后的相关研究提供了解决思路。但上述模拟方法只假设了两种类型植被在物理空间上的演变, 未考虑生物群落演替作用, 特别是草本物种组成上发生的变化, 而人工灌丛入侵引起的草本物种群落退化演替对草地生态系统碳蓄积的影响尚不明确。例如, 前人研究发现人工灌丛入侵荒漠草原会引起草地土壤有机碳的显著增加[17], 但灌丛化对典型草原的丛间草本群落地上生物量及枯落物并没有显著影响[25], 因此, 本研究忽略了人工灌丛入侵草地后对灌丛丛间草本植物生物量及碳储量的影响。此外, 本研究假定人工种植柠条灌木过程中一直未进行平茬和刈割处理, 而盐池县部分村镇存在平茬处理复壮柠条的现象。从灌木林经营的角度看, 平茬会加速灌木的生长速度, 提高生物产量, 增强固碳能力[26-27]。鉴于模型限制, 本研究未将柠条平茬与刈割因素考虑进去, 而考虑人工经营的碳储量模拟将是未来重要的研究方向。
3.2 荒漠草原人工灌丛化的碳固持作用欧妮尔等人实测出内蒙古东部兴安盟地区的柠条林的生物量是1.1647 kg/m2, 含碳率均值为0.4241[28], 根据生物量与碳储量的转换公式:生物量=碳储量×含碳率[29], 其植被碳储量为0.4939 kg/m2, 略高于本研究模拟的植被碳储量最高值(0.3143 kg/m2);而本研究模拟的植被碳储量又显著高于马文红等计算的中国北方草地地上生物碳储量平均值(0.0490 kg/m2)[30], 可见人工灌丛化导致盐池荒漠草原的碳储量显著升高, 其地上植被碳已具有灌丛生态系统的特征。从土壤碳储量来看, 本研究模拟的结果为1.8917—1.9681 kg/m2, 与丰思捷等人实测的内蒙古典型草原土壤碳储量((2.41±0.84) kg/m2)较为相近[31], 可见荒漠草原人工灌丛化后, 其土壤碳储量特征还与草地生态系统较为接近, 这与生态系统需要缓慢的过程才能将生物碳转化到土壤之中有关。以上分析可知, 本研究模拟的人工灌丛化荒漠草原的碳储量与大多数灌丛生态系统的碳储量接近, 且明显高于单一的草地生态系统, 即荒漠草原地区人工灌木的种植不仅提高了地上生物量, 还增加了生态系统的总碳固持。
4 结论利用Logistic生长曲线方程, 结合Biome-BGC模型模拟了盐池荒漠草原人工灌丛种植后的生长过程及该过程对生态系统碳储量的影响, 得出的主要结果如下:(1)人工灌丛的种植不仅会提高盐池荒漠草原碳储量, 也会改变各个碳储量的组分比例, 其中土壤碳的增涨幅度明显小于地上碳储量的增幅。(2)人工灌丛的种植会加速各个碳储量的累积速率, 但对不同碳组分的影响程度不同, 其中地上植被和枯落物碳储量累积增速最快, 导致灌丛化后生态系统的植被和枯落物碳分别增加了6倍和1.76倍, 而地下土壤碳储量仅增加了1.01倍。总之, 荒漠草原地区人工种植灌木不仅提高了地上生物量, 改善局地植被生态环境, 还能增强生态系统的碳固持能力, 为碳中和提供可行路径。
| [1] |
Rumpel C, Amiraslani F, Koutika L S, Smith P, Whitehead D, Wollenberg E. Put more carbon in soils to meet Paris climate pledges. Nature, 2018, 564(7734): 32-34. DOI:10.1038/d41586-018-07587-4 |
| [2] |
Fang J Y, Yu G R, Liu L L, Hu S J, Chapin III F S. Climate change, human impacts, and carbon sequestration in China. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(16): 4015-4020. DOI:10.1073/pnas.1700304115 |
| [3] |
Tang Z Y, Xu W T, Zhou G Y, Bai Y F, Li J X, Tang X L, Chen D M, Liu Q, Ma W H, Xiong G M, He H L, He N P, Guo Y P, Guo Q, Zhu J L, Han W X, Hu H X, Fang J Y, Xie Z Q. Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China's terrestrial ecosystems. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(16): 4033-4038. DOI:10.1073/pnas.1700295114 |
| [4] |
Fang J Y, Guo Z D, Hu H F, Kato T, Muraoka H, Son Y. Forest biomass carbon sinks in East Asia, with special reference to the relative contributions of forest expansion and forest growth. Global Change Biology, 2014, 20(6): 2019-2030. DOI:10.1111/gcb.12512 |
| [5] |
Chen J M, Ju W M, Ciais P, Viovy N, Liu R G, Liu Y, Lu X H. Vegetation structural change since 1981 significantly enhanced the terrestrial carbon sink. Nature communications, 2019, 10(1): 4259. DOI:10.1038/s41467-019-12257-8 |
| [6] |
Chen C, Park T, Wang X H, Piao S L, Xu B D, Chaturvedi R K, Fuchs R, Brovkin V, Ciais P, Fensholt R, Tømmervik H, Bala G, Zhu Z C, Nemani R R, Myneni R B. China and India lead in greening of the world through land-use management. Nature Sustainability, 2019, 2(2): 122-129. DOI:10.1038/s41893-019-0220-7 |
| [7] |
Piao S L, Wang X H, Park T, Chen C, Lian X, He Y, Bjerke J W, Chen A P, Ciais P, Tømmervik H, Nemani R R, Myneni R B. Characteristics, drivers and feedbacks of global greening. Nature Reviews Earth & Environment, 2020, 1(1): 14-27. |
| [8] |
Eldridge D J, Bowker M A, Maestre F T, Roger E, Reynolds J F, Whitford W G. Impacts of shrub encroachment on ecosystem structure and functioning: towards a global synthesis. Ecology Letters, 2011, 14(7): 709-722. DOI:10.1111/j.1461-0248.2011.01630.x |
| [9] |
Knapp A K, Briggs J M, Collins S L, Archer S R, Bret-Harte S, Ewers B E, Peters D P, Young D R, Shaver G R, Pendall E, Cleary M B. Shrub encroachment in North American grasslands: shifts in growth form dominance rapidly alters control of ecosystem carbon inputs. Global Change Biology, 2008, 14(3): 615-623. DOI:10.1111/j.1365-2486.2007.01512.x |
| [10] |
魏楠, 赵凌平, 谭世图, 赵芙蓉. 草地灌丛化研究进展. 生态科学, 2019, 38(6): 208-216. |
| [11] |
Li H, Shen H H, Chen L Y, Liu T Y, Hu H F, Zhao X, Zhou L H, Zhang P J, Fang J Y. Effects of shrub encroachment on soil organic carbon in global grasslands. Scientific Reports, 2016, 6: 28974. DOI:10.1038/srep28974 |
| [12] |
Jackson R B, Banner J L, Jobbágy E G, Pockman W T, Wall D H. Ecosystem carbon loss with woody plant invasion of grasslands. Nature, 2002, 418(6898): 623-626. DOI:10.1038/nature00910 |
| [13] |
Coetsee C, Gray E F, Wakeling J, Wigley B J, Bond W J. Low gains in ecosystem carbon with woody plant encroachment in a South African savanna. Journal of Tropical Ecology, 2013, 29(1): 49-60. DOI:10.1017/S0266467412000697 |
| [14] |
Du L T, Zeng Y J, Ma L L, Qiao C L, Wu H Y, Su Z B, Bao G. Effects of anthropogenic revegetation on the water and carbon cycles of a desert steppe ecosystem. Agricultural and Forest Meteorology, 2021, 300: 108339. DOI:10.1016/j.agrformet.2021.108339 |
| [15] |
丹杨, 杜灵通, 王乐, 马龙龙, 乔成龙, 吴宏玥, 孟晨. 荒漠草原人工灌丛化对蒸散发及其组分的影响——以盐池县为例. 生态学报, 2020, 40(16): 5638-5648. |
| [16] |
赵亚楠, 周玉蓉, 王红梅. 宁夏东部荒漠草原灌丛引入下土壤水分空间异质性. 应用生态学报, 2018, 29(11): 3577-3586. |
| [17] |
赵亚楠, 杜艳艳, 马彦平, 赵延兵, 周玉蓉, 王红梅. 宁夏东部荒漠草原灌丛引入过程中土壤有机碳变化及其空间格局预测. 应用生态学报, 2019, 30(6): 1927-1935. |
| [18] |
马龙龙, 杜灵通, 丹杨, 王乐, 乔成龙, 吴宏玥. 基于茎流-蒸渗仪法的荒漠草原带人工灌丛群落蒸散特征. 植物生态学报, 2020, 44(8): 807-818. |
| [19] |
康满春, 朱丽平, 许行, 查同刚, 张志强. 基于Biome-BGC模型的北方杨树人工林碳水通量对气候变化的响应研究. 生态学报, 2019, 39(7): 2378-2390. |
| [20] |
程杰, 王吉斌, 程积民, 罗宗宽. 黄土高原柠条锦鸡儿灌木林生长的时空变异特征. 林业科学, 2013, 49(1): 14-20. DOI:10.3969/j.issn.1006-1126.2013.01.003 |
| [21] |
赵龙, 王振凤, 郭忠升, 郭满才, 袁志发. 黄土丘陵半干旱区柠条林株高生长过程新模型. 生态学报, 2013, 33(7): 2093-2103. |
| [22] |
Thornton P E, Law B E, Gholz H L, Clark K L, Falge E, Ellsworth D S, Goldstein A H, Monson R K, Hollinger D, Falk M, Chen J, Sparks J P. Modeling and measuring the effects of disturbance history and climate on carbon and water budgets in evergreen needleleaf forests. Agricultural and Forest Meteorology, 2002, 113(1/4): 185-222. |
| [23] |
朱玉果, 杜灵通, 谢应忠, 刘可, 宫菲, 丹杨, 王乐, 郑琪琪. 2000-2015年宁夏草地净初级生产力时空特征及其气候响应. 生态学报, 2019, 39(2): 518-529. |
| [24] |
方精云, 郭兆迪, 朴世龙, 陈安平. 1981-2000年中国陆地植被碳汇的估算. 中国科学D辑: 地球科学, 2007, 37(6): 804-812. |
| [25] |
图雅, 刘艳书, 朱媛君, 杨晓晖, 张克斌. 锡林郭勒草原灌丛化对灌丛间地草本群落物种多样性和生物量的影响. 北京林业大学学报, 2019, 41(10): 57-67. |
| [26] |
郑士光, 贾黎明, 庞琪伟, 李锐. 平茬对柠条林地根系数量和分布的影响. 北京林业大学学报, 2010, 32(3): 64-69. |
| [27] |
马占英. 平茬对黄土高原人工柠条林土壤碳输入的影响[D]. 杨陵: 西北农林科技大学, 2020.
|
| [28] |
欧妮尔. 内蒙古东部山杏与柠条灌木林地上碳储量模型研究[D]. 呼和浩特: 内蒙古农业大学, 2017.
|
| [29] |
孙玉军, 张俊, 韩爱惠, 王雪军, 王新杰. 兴安落叶松(Larix gmelini)幼中龄林的生物量与碳汇功能. 生态学报, 2007, 27(5): 1756-1762. DOI:10.3321/j.issn:1000-0933.2007.05.011 |
| [30] |
马文红, 方精云, 杨元合, 安尼瓦尔·买买提. 中国北方草地生物量动态及其与气候因子的关系. 中国科学: 生命科学, 2010, 40(7): 632-641. |
| [31] |
丰思捷, 赵艳云, 李元恒, 闫勇智, 韩芳. 内蒙古典型草原表层土壤有机碳储量差异及影响因素. 中国草地学报, 2019, 41(2): 116-120. |