 2022, Vol. 42
2022, Vol. 42文章信息
- 王思齐, 朱章明
- WANG Siqi, ZHU Zhangming
- 中国蔷薇属植物物种丰富度分布格局及其与环境因子的关系
- Relationships between species richness patterns of Rosa L. and environmental factors in China
- 生态学报. 2022, 42(1): 209-219
- Acta Ecologica Sinica. 2022, 42(1): 209-219
- http://dx.doi.org/10.5846/stxb202002280363
-
文章历史
- 收稿日期: 2020-02-28
- 网络出版日期: 2021-08-16

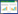




物种丰富度的大尺度地理格局及其成因是宏观生态学及生物地理学的核心问题[1-2], 是衡量区域多样性的基本尺度以及构建进化、生态模型和保护策略的基础[3-4]。物种丰富度是进化历史、生态过程、现代环境等因素共同驱动的结果[2, 5], 其中现代环境条件被认为是大尺度地理范围内物种丰富度的主要驱动因素[4, 6-7]。现代环境条件主要包括现代气候(热量、水分及其季节性变化等)和生境异质性, 并由此产生了能量假说、环境稳定性假说和生境异质性假说。其中能量假说认为, 一个地区的物种丰富度主要受能量控制, 能量越高的地方其物种丰富度就越高[6, 8-9], 经典的能量假说至少包括了生产力假说、水能-动态假说、寒冷忍耐假说及生态学代谢假说四种形式[6, 10-13];环境稳定性假说认为, 稳定的环境条件有利于物种的特化和适应, 使生态位趋于狭窄化, 有利于容纳更多的物种[14-15];生境异质性假说认为, 生境异质性高的地方能提供更多的生态位, 从而有利于多个物种共存[11]。这些假说基于不同的影响因子探讨了物种丰富度形成的主要因素。目前, 这些假说已在一些研究中被验证, 如中国裸子植物(Gymnospermae)[16]、中国种子植物(Spermatophyta)[17]、中国兰科(Orchidaceae)植物[18]等。类似研究多在科级以上水平展开, 而对属级水平的物种丰富度分布格局及其形成机制的探索仍较欠缺。
蔷薇属(Rosa L.)隶属于蔷薇科(Rosaceae), 全世界约有150—200种, 广泛分布于北半球的温带和亚热带地区。中国有蔷薇属植物95种, 其中65种为特有种, 是蔷薇属的现代分布中心[19]。蔷薇属植物花美芳香, 是重要的观赏植物和芳香油植物;果实富含维生素C, 具有很高的食用价值[20-21]。此外, 蔷薇属植物基因组中包含抗旱、抗寒、抗病等优良性状基因, 是引种驯化的重要遗传资源[22];许多蔷薇属物种抗逆性强, 能在恶劣环境下生存, 常作为生态修复、植被恢复的建群种[23]。目前蔷薇属植物的野生资源研究比较滞后, 部分物种处于濒危状态, 资源丢失严重[24-25], 而对国内蔷薇属植物的分布未有系统整理。深入研究蔷薇属分布格局及其与环境因子间的关系, 探索主要环境因子对蔷薇属物种丰富度的影响, 有助于我国蔷薇属植物资源的保护、引种、开发和利用, 对蔷薇属植物的分类、系统进化和遗传多样性保护研究也具有一定的意义。
本研究基于标本记录和文献资料, 利用GIS技术, 探讨蔷薇属植物在中国的丰富度分布格局, 并结合环境因子, 探讨形成这一格局的主要驱动因素及该属植物较适宜的生长环境。以期为蔷薇属植物的分类、进化、资源调查、收集、良种选育及物种保护等研究与应用提供参考。
1 数据与方法 1.1 数据来源和处理 1.1.1 物种分布数据中国蔷薇属植物分布数据来源于中国数字植物标本馆(CVH, http://www.cvh.ac.cn/), 并依据《中国植物志》[20]和《Flora of China》[19]进行校对。剔除人工栽培种, 将变种、变型等种下等级计为一个独立的分类单元, 并将分布信息统一至县级单元[26]。最终得到的数据集共包括中国蔷薇属的125个种及种下单元, 889条县区级数据, 15451条分布信息。
1.1.2 环境数据通过查阅相关文献, 本研究选取了植物物种丰富度研究中常用的11种气候(包括能量和水分因子)和生境异质性因子[27-29]。年平均气温(Mean annual temperature, MAT)、最热月平均气温(Mean temperature of the warmest month, MTWM)、最冷月平均气温(Mean temperature of the coldest month, MTCM)、气温年较差(Temperature annual range, ART)、年均降雨量(Annual precipitation, AP)和降水季节性(Precipitation seasonality, PSN)数据来源于WorldClim数据库(http://worldclim.org/version2)(1970—2000年), 原始分辨率为1km×1km;年实际蒸散量(Actual evapotranspiration, AET)来源于国际农业研究组织(http://www.cgiarcsi.org/)(1970—2000年), 原始分辨率为1km×1km;湿润指数(Moisture index, MI)来自中国科学院资源环境科学数据中心(http://www.resdc.cn/DOI/doi.aspx?DOIid=39), 原始分辨率为500m×500m;海拔变化范围(Range of elevation, ELER)根据各栅格内最高海拔与最低海拔之差计算, 海拔数据来自EarthEnv数据库(http://www.earthenv.org/topography), 原始分辨率为1km×1km;气温空间差异(Range of mean annual temperature, MATR)和降水空间差异(Range of annual precipitation, APR)分别为每个栅格内的年平均降水和年平均气温的最大值和最小值之差。
将等面积网格与环境数据叠加, 计算所得各环境因子在每个网格内的平均值即为每个网格的环境数据。环境数据的提取和计算均利用ArcGIS 10.6完成。
1.2 数据分析 1.2.1 物种丰富度统计物种丰富度以物种数量表示。首先, 利用ArcGIS 10.6进行等面积圆锥投影(Albers equal area conic projection)的投影转换;其次, 为了减小不同县区的面积差异对丰富度的影响, 使分析时单位面积相同[30], 因此将中国政区图转化为100km×100km的等面积网格, 去除面积不足75%的网格, 共得到901个网格;然后, 提取各县区的中心坐标, 依据中心坐标点将各县区的物种丰富度与网格进行关联, 取各网格内所有县区拥有的总物种数(同一网格内不同县区的相同物种不重复统计)作为该网格的物种丰富度;最后, 利用ArcGIS中的Jenks分级功能, 将物种丰富度划分为五个等级。提取最大分级区间对应网格中的环境变量作为蔷薇属植物生长条件的适宜范围。
1.2.2 物种丰富度与环境因子相关性分析首先, 对不满足正态分布的数据进行的Y=log(Y+1)对数转换;其次, 采用回归分析考察物种丰富度与经纬度和各环境因子的关系;然后, 为了进一步探究能量、水分和生境异质性的影响程度, 对物种丰富度和上述环境因子分别进行了逐步回归分析, 同时增加一个综合模型(即分别进行逐步回归分析后筛选出的全部环境因子), 通过R2进而对能量、水分、生境异质性及综合模型的影响程度进行评估。同时为了消除各环境因子之间多重共线性的影响, 在逐步回归时选择方差膨胀因子(Variance inflation factor, VIF)小于10的模型进行分析。相关性分析均利用SPSS 22完成。
2 结果与分析 2.1 中国蔷薇属物种丰富度空间分布格局中国蔷薇属植物空间分布不均匀, 每个网格的物种丰富度在0—43种之间(图 1)。在省级尺度上, 蔷薇属物种丰富度最高的省份为四川省和云南省, 分别有67和62种蔷薇属植物分布, 共计82种, 占全国总物种数的65.6%。其次为甘肃、陕西、重庆、西藏、贵州、河南和湖北, 物种数均在20种以上。

|
| 图 1 中国蔷薇属物种丰富度格局 Fig. 1 Species richness pattern of Rosa in China |
蔷薇属植物在中国的水平分布范围为18.89°—53.04°N, 75.13°—133.56°E;垂直分布范围为3.08—5455.02m(表 1)。对蔷薇属物种丰富度的经纬度和海拔的回归分析结果(图 2)表明:(1)在水平方向上, 中国蔷薇属植物在26.19°—34.29°N区域内呈现出较高的物种丰富度, 向北随着纬度增加而逐渐下降;在99.10°—108.47°E区域内物种丰富度达到最大值, 在此高值范围的东西两侧, 随着经度的增大或减小均下降。(2)在垂直方向上, 中国蔷薇属物种丰富度随着海拔的升高呈现先增加后减少的单峰形式, 峰值在956.46—3518.60m范围内。
| 项目 Item |
最大值 Max |
平均值 Mean |
最小值 Min |
标准差 SD |
适宜范围Optimal range | |
| 下限 Lower limit |
上限 Upper limit |
|||||
| 物种丰富度Species richness | 43 | 2.89 | 0 | 5.27 | ||
| 地理因子Geographic factors | ||||||
| 北纬Latitude/(°) | 53.04 | 35.81 | 18.89 | 6.93 | 26.19 | 34.29 |
| 东经Longitude/(°) | 133.56 | 103.46 | 75.13 | 13.87 | 99.10 | 108.47 |
| 海拔Altitude/m | 5455.02 | 1848.20 | 3.08 | 1699.11 | 956.46 | 3518.60 |
| 气候因子Climatic factors | ||||||
| (1)能量因子Energy factors | ||||||
| 年均温Mean annual temperature/℃ | 23.88 | 6.53 | -11.21 | 8.10 | 1.90 | 14.91 |
| 最热月均温 Mean temperature of the warmest month/℃ |
29.41 | 19.28 | 1.86 | 7.73 | 11.62 | 24.44 |
| 最冷月均温 Mean temperature of the coldest month/℃ |
18.29 | -7.92 | -27.87 | 10.00 | -8.20 | 6.99 |
| 气温年较差 Temperature annual range/℃ |
46.57 | 27.20 | 9.29 | 8.18 | 11.95 | 23.50 |
| (2)水分因子Water factors | ||||||
| 年均降水Annual precipitation/mm | 2599.81 | 591.43 | 16.19 | 496.20 | 696.38 | 1339.14 |
| 实际蒸散量Actual evapotranspiration/mm | 1169.90 | 467.77 | 20.11 | 337.58 | 607.76 | 942.91 |
| 湿润指数Moisture index | 11956.74 | -646.92 | -10885.27 | 4628.45 | 925.06 | 10790.28 |
| 降水季节性Precipitation seasonality | 141.82 | 89.06 | 34.76 | 22.25 | 62.60 | 92.20 |
| 生境异质性因子Habitat heterogeneity | ||||||
| 气温空间差异Range of mean annual temperature/℃ | 44.00 | 9.77 | 0.00 | 7.75 | 9.00 | 39.00 |
| 降水空间差异Range of annual precipitation/mm | 3240.00 | 239.08 | 6.00 | 272.55 | 183.00 | 984.00 |
| 海拔变化范围Range of elevation/m | 6677.88 | 1611.82 | 11.69 | 1129.03 | 1961.06 | 5354.88 |

|
| 图 2 中国蔷薇属物种丰富度与经度、纬度和海拔的关系 Fig. 2 The relationships between the richness of Rosa in China and longitude, latitude and elevation |
蔷薇属植物在中国分布区的年均温度范围为-11.21—23.88℃, 最适范围为1.90—14.91℃;年均降水范围为16.19—2599.81mm, 最适范围为696.38—1339.14mm(表 1)。相比于冷杉属(Abies Mill.)、云杉属(Picea Dietr.)、落叶松属(Larix Mill.)[31]、闽楠(Phoebe bournei (Hemsl.) Yang)[32]及黄连木(Pistacia chinensis Bunge)[33]等其他植物类群来说, 蔷薇属分布区内各地理与气候因子的变幅和标准差更大, 数据更离散, 可见中国蔷薇属物种的地理分布格局广, 对环境的生态适应能力强。
对中国蔷薇属植物物种丰富度和各环境因子的一元线性回归分析结果表明, 该属物种丰富度与各环境因子均有极显著相关关系(P < 0.001)。在能量因子中, 物种丰富度与年均温、最热月均温和最冷月均温均呈正相关, 与气温年较差呈负相关, 说明在热量条件更好、气温年变化较小的地方其丰富度更高;在水分因子中, 物种丰富度与年均降水量、实际蒸散量和湿润指数均呈正相关, 而与降水季节性呈负相关, 说明其在水分条件更好、降水季节性变化较小的地方其丰富度更高;在生境异质性因子中, 物种丰富度与气温空间差异、降水空间差异和海拔变化范围均呈正相关, 说明在生境异质性强的地方其丰富度更高。当单独考虑环境因子时, 实际蒸散量和湿润指数对蔷薇属物种丰富度格局的影响最大, 方差解释率分别为30%和27.7%(图 3)。

|
| 图 3 中国蔷薇属物种丰富度与环境因子的关系 Fig. 3 The relationships between species richness of Rosa in China and environmental factors |
为了进一步探究能量、水分、生境异质性以及全部环境因子对中国蔷薇属物种丰富度的影响程度, 并消除变量间多重共线性的影响, 以蔷薇属物种丰富度为因变量, 以上述四种模型为自变量分别进行的逐步回归分析(表 2)结果表明:在四种模型中, 综合模型对蔷薇属物种丰富度的解释率最高, 达45.9%;其次为水分模型, 解释率达34.6%;能量模型和生境异质性模型的解释率分别为29.1%和24.9%。中国蔷薇属物种丰富度分布格局受到能量、水分和生境异质性等各种环境因子的综合影响, 其中水分模型起到主导作用。
| 模型 Model |
环境因子 Environmental factors |
R2/% | P |
| 能量Energy | 最冷月均温(MTCM)、年均温(MAT) | 29.1 | < 0.001 |
| 水分Water | 实际蒸散量(AET)、降水季节性(PSN)、湿润指数(MI) | 34.6 | < 0.001 |
| 生境异质性Habitat heterogeneity | 降水空间差异(APR)、海拔变化范围(ELER)、气温空间差异(MATR) | 24.9 | < 0.001 |
| 综合Mixed model | 湿润指数(MI)、海拔变化范围(ELER)、年均温(MAT)、气温空间差异(MATR)、年均降水(AP) | 45.9 | < 0.001 |
植物物种丰富度由赤道向两极递减是地球表面最显著的生态格局之一[10, 17, 34-37]。在经度方向上, 中国植物类群物种丰富度普遍呈单峰关系, 且峰值多于95—110°E范围内[27, 29, 38], 因此, 中国植物物种丰富度最高的经纬度交汇区域为西南横断山脉地区[7, 39-42]。而在垂直方向上, 植物物种丰富度随着海拔升高呈现出先增加后减少, 并在中海拔地区出现峰值的物种垂直分布格局[43-44], 如中国横断山区的典型类群如乌头属(Aconitum L.)、无心菜属(Arenaria L.)、嵩草属(Kobresia Willd.)等[45], Aberdare山区的菊科(Asteraceae)、禾本科(Poaceae)、豆科(Fabaceae)等[46]及中国亚麻酸资源植物如唇形科(Lamiaceae)、大戟科(Euphorbiaceae)等[43]。蔷薇属植物在水平方向上的最适分布区主要为我国西南横断山区及四川盆地周边的山脉, 这与中国植物物种水平分布的普遍规律相适应;在垂直方向上, 中国蔷薇属植物物种丰富度呈现单峰形式, 峰值在956.46—3518.60m范围内, 这与物种随海拔变化的普遍规律相符合。
中国蔷薇属植物在水分和热量条件好、气候季节性变化小且生境异质性程度高的地方, 有更高的物种丰富度(图 3)。中国西南的横断山区及四川盆地周边的山脉都具有降水丰富、年均温适中、地形复杂、生境异质性高等特点[16, 42-43], 有利于蔷薇属物种的生长, 因此是蔷薇属植物分布的最适区域。除此之外, 在新疆北部以及东北长白山周边地区也有零星区域呈现出较高的物种丰富度。蔷薇属内的一些物种如宽刺蔷薇(R. platyacantha Schrenk)、疏花蔷薇(R. laxa Retz.)、刺蔷薇(R. acicularis Lindl.)、腺齿蔷薇(R. albertii Regel)、弯刺蔷薇(R. beggeriana Schrenk)等[20], 它们有着特殊的生态适应特征和形态特征[47], 既抗寒耐热、又抗旱耐淹, 能够在新疆生存[48-49];且新疆北部地区与南部相比, 年平均降水相对较多, 年平均气温相对较低[50-51], 而蔷薇属植物分布主要受水分的影响, 因此蔷薇属在新疆北部地区形成了局部聚集区域。同样, 刺蔷薇(R. acicularis Lindl.)、山刺玫(R. davurica Pall.)等物种也依靠其本身的适应能力, 在东北长白山周边等纬度较高、气候相对恶劣的地区形成了局部聚集区。因此, 蔷薇属植物分布区内的各环境因子数据离散性大, 蔷薇属植物的生态适应性广。
总之, 我国低纬地区相对于高纬地区有着更好的水热条件, 且根据Rapoport法则, 低纬地区的气候稳定性更强[17, 52];山区相对于平原地区的地形更复杂, 具有更高的生境异质性。因此, 中国蔷薇属植物呈现出在低纬地区比高纬地区、海拔落差大的山区比平原地区具有更高的物种丰富度的分布特征。部分蔷薇属物种在纬度较高、气候相对恶劣地区形成小范围聚集区域。
3.2 蔷薇属物种的丰富度格局支持多种环境假说中国蔷薇属物种丰富度格局与多个环境因子显著相关, 其中实际蒸散量的解释率最大, 根据生产力假说, 能量和水分的增加会提高一个地区净初级生产力, 从而对应更高的物种丰富度[10, 53-55], 而实际蒸散量表示从地表向大气输送的实际水量, 能代表太阳能和水分的双重影响, 因此实际蒸散量被认为是表征生产力假说的最好的单变量气候指标[10, 13], 蔷薇属植物物种丰富度随实际蒸散量的增加而增加, 符合生产力假说的预测。气温年较差反映了日均温在一年内的波动程度, 可作为环境稳定性的衡量指标[1, 8, 26, 56], 中国蔷薇属植物物种丰富度随气温年较差的增大而显著降低, 反映了该属物种适宜分布在气候相对稳定的地区, 这可能是因为稳定的环境条件有利于物种的特化和适应, 使生态位趋于狭窄化, 从而有利于容纳更多的物种[14-15]。在第四纪冰期和间冰期时气温变化较大, 而复杂地形可以提供多种生境, 从而为不同植物物种在极端气候条件下的生存提供避难所[57-59];并且在相互隔离的生境中由于奠基者效应、遗传漂变等机制, 促进了物种的分化过程, 有利于物种形成[60-61], 因此生境异质性高的地方有更高的物种丰富度。而本研究中海拔变化范围和气温空间差异、降水空间差异[16, 42, 62]均与蔷薇属丰富度呈正相关, 且蔷薇属植物的适宜分布区集中在横断山区及四川盆地周边的山脉, 该分布区地形复杂, 单位面积内拥有更多的生境类型, 从而有利于多个物种共存, 这些都较好的支持了生境异质性假说[11]。
3.3 水分是影响蔷薇属物种丰富度格局的主要因素物种丰富度分布格局与环境因子的复杂关系, 通常与该类群的进化历史和生理适应有关[63]。本研究结果表明综合模型对蔷薇属丰富度格局的贡献最大, 说明中国蔷薇属丰富度分布格局同时受到能量、水分和生境异质性等多种环境因子的共同影响, 没有任何单一的环境因子或假说能对物种的丰富度格局做出充分解释[2]。除综合模型外, 水分是影响蔷薇属丰富度的最主要因子, 这与蔷薇科植物多样性格局与环境因子相关性的研究结果一致[42]。从进化历史来看, 蔷薇属植物起源于东亚地区, 为典型的温带气候区[64], 且其生态位在第四纪冰期时大范围扩张, 在间冰期温度回升时向高海拔地区收缩, 为典型的冷适应性类群[59], 因此相对于水分因子来说, 与温度有关的各环境因子的限制性较弱。从生理上来看, 干旱胁迫会降低植物的光合速率[65], 从而破坏植物细胞的稳态, 同时关键生长阶段的水分缺乏会降低植物果实的大小和结实率, 该现象在桃(Prunus persica (L.) Batsch.)[66]、苹果(Malus pumila Mill. cv. Braeburn)[67]、覆盆子(Rubus idaeus L.)[68]等蔷薇科植物中均有报道。并且从分布格局上看, 蔷薇属物种多分布于南方地区, 这些地方一般热量条件较好, 水分便成为更重要的限制条件[69-70]。因此, 选择蔷薇属植物异地保护或园艺栽培地点时首先应考虑水分条件, 其次是热量条件。本研究获得的蔷薇属植物生长的环境条件适宜区间可为蔷薇属植物的保护和开发利用提供依据。
综合模型对于中国蔷薇属的物种丰富度有较高的适合性, 但其无法解释的部分可能与过去的环境条件和进化历史、地形、土壤理化性质、人类活动及其他因素有关[5, 71-75]。
4 结论本研究利用蔷薇属植物在中国的分布数据, 结合相应的环境因子, 探讨了中国蔷薇属植物的丰富度分布格局及其影响因素, 发现中国蔷薇属植物分布的最适区域为中国西南横断山区和四川盆地周边的山脉, 新疆北部以及东北长白山周边地区形成局部聚集区域, 并且在垂直方向上集中分布于中高海拔区域;水热条件好、气候季节性变化小且生境异质性程度高的地方, 有更高的物种丰富度, 且水分是影响中国蔷薇属物种丰富度最重要的环境因子。
| [1] |
林鑫, 王志恒, 唐志尧, 赵淑清, 方精云. 中国陆栖哺乳动物物种丰富度的地理格局及其与环境因子的关系. 生物多样性, 2009, 17(6): 652-663. |
| [2] |
Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220-227. DOI:10.1038/35012228 |
| [3] |
Gotelli N J, Colwell R K. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecology Letters, 2001, 4(4): 379-391. DOI:10.1046/j.1461-0248.2001.00230.x |
| [4] |
D'Antraccoli M, Roma-Marzio F, Carta A, Landi S, Bedini G, Chiarucci A, Peruzzi L. Drivers of floristic richness in the Mediterranean: a case study from Tuscany. Biodiversity and Conservation, 2019, 28(6): 1411-1429. DOI:10.1007/s10531-019-01730-x |
| [5] |
Rao M D, Steinbauer M J, Xiang X G, Zhang M G, Mi X C, Zhang J T, Ma K P, Svenning J C. Environmental and evolutionary drivers of diversity patterns in the tea family (Theaceae s.s.) across China. Ecology and Evolution, 2018, 8(23): 11663-11676. DOI:10.1002/ece3.4619 |
| [6] |
Hawkins B A, Field R, Cornell H V, Currie D J, Guégan J F, Kaufman D M, Kerr J T, Mittelbach G G, Oberdorff T, O'Brien E M, Porter E E, Turner J R G. Energy, water, and broad-scale geographic patterns of species richness. Ecology, 2003, 84(12): 3105-3117. DOI:10.1890/03-8006 |
| [7] |
Wang Z H, Fang J Y, Tang Z Y, Lin X. Relative role of contemporary environment versus history in shaping diversity patterns of China's woody plants. Ecography, 2012, 35(12): 1124-1133. DOI:10.1111/j.1600-0587.2011.06781.x |
| [8] |
刘澈, 郑成洋, 张腾, 曾发旭, 王逸然. 中国鸟类物种丰富度的地理格局及其与环境因子的关系. 北京大学学报: 自然科学版, 2014, 50(3): 429-438. DOI:10.3969/j.issn.1671-167X.2014.03.017 |
| [9] |
吴建国, 周巧富. 中国嵩草属植物丰富度与气候要素的关系. 应用生态学报, 2012, 23(4): 1003-1017. |
| [10] |
王志恒, 唐志尧, 方精云. 物种多样性地理格局的能量假说. 生物多样性, 2009, 17(6): 613-624. |
| [11] |
Tews J, Brose U, Grimm V, Tielbörger K, Wichmann M C, Schwager M, Jeltsch F. Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. Journal of Biogeography, 2004, 31(1): 79-92. DOI:10.1046/j.0305-0270.2003.00994.x |
| [12] |
Stevens G C. The latitudinal gradient in geographical range: how so many species coexist in the tropics. The American Naturalist, 1989, 133(2): 240-256. DOI:10.1086/284913 |
| [13] |
谢婷. 中国木本植物物种丰富度格局的统计学机制研究[D]. 北京: 华北电力大学(北京), 2019.
|
| [14] |
Klopfer P H, MacArthur R H. Niche size and faunal diversity. The American Naturalist, 1960, 94(877): 293-300. DOI:10.1086/282130 |
| [15] |
林晓倩, 黄勇, 左智力, 陈跃英. 不同尺度中国大陆灵长目物种丰富度的地理格局及其与环境因子和人类活动的关系. 应用与环境生物学报, 2012, 18(6): 954-963. |
| [16] |
吕丽莎, 蔡宏宇, 杨永, 王志恒, 曾辉. 中国裸子植物的物种多样性格局及其影响因子. 生物多样性, 2018, 26(11): 1133-1146. DOI:10.17520/biods.2018098 |
| [17] |
冯建孟, 徐成东. 中国种子植物物种丰富度的大尺度分布格局及其与地理因子的关系. 生态环境学报, 2009, 18(1): 249-254. DOI:10.3969/j.issn.1674-5906.2009.01.047 |
| [18] |
张晓龙. 中国野生兰科植物地理分布格局研究[D]. 太原: 山西大学, 2014.
|
| [19] |
Ku T C, Robertson K R. Rosa Linnaeus//Wu C Y, Raven P H, eds. Flora of China. Beijing/St. Louis: Science Press/Missouri Botanical Garden Press, 2003, Vol. 9: 339-381.
|
| [20] |
俞德浚, 陆玲娣, 谷粹芝, 关克俭, 李朝銮. 蔷薇科//中国科学院中国植物志编辑委员会. 中国植物志. 北京: 科学出版社, 1985, Vol. 37: 360-455.
|
| [21] |
章银柯, 江燕, 朱炜. 我国蔷薇属植物资源及其园林应用前景. 种子, 2009, 28(8): 68-70. DOI:10.3969/j.issn.1001-4705.2009.08.021 |
| [22] |
冯立国. 玫瑰野生种质资源评价及其与栽培种质亲缘关系的研究[D]. 济南: 山东农业大学, 2007.
|
| [23] |
金晶, 金平, 吴洪娥, 董万鹏, 杨成华, 周洪英. 贵州蔷薇属植物资源调查与应用研究. 种子, 2020, 39(8): 61-65, 69-69. |
| [24] |
周玉泉, 苏群, 张颢, 李树发, 唐开学, 蹇洪英. 极危植物中甸刺玫的分布及种群数量动态. 植物遗传资源学报, 2016, 17(4): 649-654, 662-662. |
| [25] |
覃海宁, 杨永, 董仕勇, 何强, 贾渝, 赵莉娜, 于胜祥, 刘慧圆, 刘博, 严岳鸿, 向建英, 夏念和, 彭华, 李振宇, 张志翔, 何兴金, 尹林克, 林余霖, 刘全儒, 侯元同, 刘演, 刘启新, 曹伟, 李建强, 陈世龙, 金效华, 高天刚, 陈文俐, 马海英, 耿玉英, 金孝锋, 常朝阳, 蒋宏, 蔡蕾, 臧春鑫, 武建勇, 叶建飞, 赖阳均, 刘冰, 林秦文, 薛纳新. 中国高等植物受威胁物种名录. 生物多样性, 2017, 25(7): 696-744. |
| [26] |
刘路平. 山东省鸟类物种丰富度的地理格局及其与环境因子的关系[D]. 济南: 山东师范大学, 2017.
|
| [27] |
王芳, 熊喆, 延晓冬, 戴新刚, 李亚飞, 王磊斌. 区域气候与中国柳属物种多样性格局的关系研究. 气候与环境研究, 2019, 24(2): 262-276. |
| [28] |
王寅, 王键铭, 崔盼杰, 钟悦鸣, 李景文, 褚建民. 科尔沁沙地植物物种丰富度格局及其与环境的关系. 植物科学学报, 2018, 36(6): 794-803. |
| [29] |
Chen S B, Jiang G M, Ouyang Z Y, Xu W H, Xiao Y. Relative importance of water, energy, and heterogeneity in determining regional pteridophyte and seed plant richness in China. Journal of Systematics and Evolution, 2011, 49(2): 95-107. DOI:10.1111/j.1759-6831.2011.00120.x |
| [30] |
Whittaker R J, Willis K J, Field R. Scale and species richness: towards a general, hierarchical theory of species diversity. Journal of Biogeography, 2001, 28(4): 453-470. DOI:10.1046/j.1365-2699.2001.00563.x |
| [31] |
刘增力, 方精云, 朴世龙. 中国冷杉、云杉和落叶松属植物的地理分布. 地理学报, 2002, 57(5): 577-586. DOI:10.3321/j.issn:0375-5444.2002.05.010 |
| [32] |
葛永金, 王军峰, 方伟, 叶荣华. 闽楠地理分布格局及其气候特征研究. 江西农业大学学报, 2012, 34(4): 749-753, 761-761. DOI:10.3969/j.issn.1000-2286.2012.04.022 |
| [33] |
符瑜, 潘学标, 高浩. 中国黄连木的地理分布与生境气候特征分析. 中国农业气象, 2009, 30(3): 318-322. DOI:10.3969/j.issn.1000-6362.2009.03.008 |
| [34] |
王健铭, 崔盼杰, 钟悦鸣, 李景文, 褚建民. 阿拉善高原植物区域物种丰富度格局及其环境解释. 北京林业大学学报, 2019, 41(3): 14-23. |
| [35] |
李果, 沈泽昊, 应俊生, 方精云. 中国裸子植物物种丰富度空间格局与多样性中心. 生物多样性, 2009, 17(3): 272-279. |
| [36] |
王宇卓. 中国山茶科物种多样性的地理格局研究[D]. 太原: 山西大学, 2017.
|
| [37] |
Willig M R, Kaufman D M, Stevens R D. Latitudinal gradients of biodiversity: pattern, process, scale, and synthesis. Annual Review of Ecology, Evolution, and Systematics, 2003, 34: 273-309. DOI:10.1146/annurev.ecolsys.34.012103.144032 |
| [38] |
郭子良, 王龙飞. 中国兰科植物沿经纬度的水平分布格局. 生物学杂志, 2013, 30(5): 49-53. DOI:10.3969/j.issn.2095-1736.2013.05.049 |
| [39] |
应俊生. 中国种子植物物种多样性及其分布格局. 生物多样性, 2001, 9(4): 393-398. DOI:10.3321/j.issn:1005-0094.2001.04.011 |
| [40] |
Wang Z H, Fang J Y, Tang Z Y, Lin X. Patterns, determinants and models of woody plant diversity in China. Proceedings of the Royal Society B: Biological Sciences, 2011, 278(1715): 2122-2132. DOI:10.1098/rspb.2010.1897 |
| [41] |
Zhang Y B, Du H D, Jin X H, Ma K P. Species diversity and geographic distribution of wild Orchidaceae in China. Chinese Science Bulletin (Chinese Version), 2015, 60(02): 179-188+171-116. |
| [42] |
邹东廷, 王庆刚, 罗奥, 王志恒. 中国蔷薇科植物多样性格局及其资源植物保护现状. 植物生态学报, 2019, 43(1): 1-15. |
| [43] |
肖佳伟, 孙林, 陈功锡, 张梦华. 中国亚麻酸资源植物分布格局及其与气候和地理因子的关系. 西北植物学报, 2016, 36(3): 606-617. |
| [44] |
Rahbek C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecology Letters, 2005, 8(2): 224-239. |
| [45] |
Zhang D C, Zhang Y H, Boufford D E, Sun H. Elevational patterns of species richness and endemism for some important taxa in the Hengduan Mountains, southwestern China. Biodiversity and Conservation, 2009, 18(3): 699-716. DOI:10.1007/s10531-008-9534-x |
| [46] |
Kipkoech S, Melly D K, Mwema B W, Mwachala G, Musili P M, Hu G W, Wang Q F. Conservation priorities and distribution patterns of vascular plant species along environmental gradients in Aberdare ranges forest. PhytoKeys, 2019, 131: 91-113. DOI:10.3897/phytokeys.131.38124 |
| [47] |
冯久莹, 蔡蕾, 贺海洋, 沈树祥, 张树玲, 孟永禄, 戴凡炜, 许正, 高俊平. 新疆14种野生蔷薇属植物生境调查. 林业科学, 2014, 50(11): 44-51. |
| [48] |
马燕, 陈俊愉. 我国西北的蔷薇属种质资源. 中国园林, 1990, 6(1): 50-51. |
| [49] |
刘彤. 新疆6种蔷薇属植物对干旱胁迫的生理响应及抗旱性评价[D]. 乌鲁木齐: 新疆农业大学, 2017.
|
| [50] |
普宗朝, 张山清, 王胜兰, 周晓丽, 冯志敏. 近48a新疆干湿气候时空变化特征. 中国沙漠, 2011, 31(6): 1563-1572. |
| [51] |
范丽红. 新疆天山山区与南、北疆气候变化及其影响的比较研究[D]. 乌鲁木齐: 新疆大学, 2006.
|
| [52] |
Rapoport E H. Areography: Geographical Strategies of Species. Oxford: Pergamon Press, 1982.
|
| [53] |
Brown J H. Two decades of homage to Santa Rosalia: toward a general theory of diversity. American Zoologist, 1981, 21(4): 877-888. DOI:10.1093/icb/21.4.877 |
| [54] |
Rosenzweig M L. Species diversity in space and time. Cambridge: Cambridge University Press, 1995, 971-971. |
| [55] |
Clarke A, Gaston K J. Climate, energy and diversity. Proceedings of the Royal Society B: Biological Sciences, 2006, 273(1599): 2257-2266. DOI:10.1098/rspb.2006.3545 |
| [56] |
冯建孟. 中国种子植物物种多样性的大尺度分布格局及其气候解释. 生物多样性, 2008, 16(5): 470-476. DOI:10.3321/j.issn:1005-0094.2008.05.007 |
| [57] |
沈浪, 陈小勇, 李媛媛. 生物冰期避难所与冰期后的重新扩散. 生态学报, 2002, 22(11): 1983-1990. DOI:10.3321/j.issn:1000-0933.2002.11.026 |
| [58] |
陈冬梅, 康宏樟, 刘春江. 中国大陆第四纪冰期潜在植物避难所研究进展. 植物研究, 2011, 31(5): 623-632. |
| [59] |
高云东, 张羽, 高信芬, 朱章明. 绢毛蔷薇复合体的谱系地理研究//生态文明建设中的植物学: 现在与未来——中国植物学会第十五届会员代表大会暨八十周年学术年会论文集——第1分会场: 系统与进化植物学. 南昌: 中国植物学会, 2013.
|
| [60] |
Wang L Y, Abbott R J, Zheng W, Chen P, Wang Y J, Liu J Q. History and evolution of alpine plants endemic to the Qinghai-Tibetan Plateau: Aconitum gymnandrum (Ranunculaceae). Molecular Ecology, 2009, 18(4): 709-721. DOI:10.1111/j.1365-294X.2008.04055.x |
| [61] |
Hughes C, Eastwood R. Island radiation on a continental scale: exceptional rates of plant diversification after uplift of the Andes. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(27): 10334-10339. DOI:10.1073/pnas.0601928103 |
| [62] |
沈梦伟, 陈圣宾, 毕孟杰, 陈文德, 周可新. 中国蚂蚁丰富度地理分布格局及其与环境因子的关系. 生态学报, 2016, 36(23): 7732-7739. |
| [63] |
王波, 黄勇, 李家堂, 戴强, 王跃招, 杨道德. 西南喀斯特地貌区两栖动物丰富度分布格局与环境因子的关系. 生物多样性, 2018, 26(9): 941-950. |
| [64] |
Li M, Ohi-Toma T, Gao Y D, Xu B, Zhu Z M, Ju W B, Gao X F. Molecular phylogenetics and historical biogeography of Sorbus sensu stricto (Rosaceae). Molecular Phylogenetics and Evolution, 2017, 111: 76-86. DOI:10.1016/j.ympev.2017.03.018 |
| [65] |
Bosabalidis A M, Kofidis G. Comparative effects of drought stress on leaf anatomy of two olive cultivars. Plant Science, 2002, 163(2): 375-379. DOI:10.1016/S0168-9452(02)00135-8 |
| [66] |
Sun M X, Peng F T, Xiao Y S, Yu W, Zhang Y F, Gao H F. Exogenous phosphatidylcholine treatment alleviates drought stress and maintains the integrity of root cell membranes in peach. Scientia Horticulturae, 2020, 259: 108821. DOI:10.1016/j.scienta.2019.108821 |
| [67] |
Mpelasoka B S, Behboudian M H, Green S R. Effects of irrigation regime and crop load on water use and yield of 'Braeburn' apple grown in lysimeters//Proceedings of the third international symposium on irrigation of horticultural crops, Vols 1 and 2. Estoril, Portugal: 1999: 741-748.
|
| [68] |
Morales C G, Pino M T, del Pozo A. Phenological and physiological responses to drought stress and subsequent rehydration cycles in two raspberry cultivars. Scientia Horticulturae, 2013, 162: 234-241. DOI:10.1016/j.scienta.2013.07.025 |
| [69] |
Hawkins B A, Porter E E, Diniz-Filho J A F. Productivity and history as predictors of the latitudinal diversity gradient of terrestrial birds. Ecology, 2003, 84(6): 1608-1623. DOI:10.1890/0012-9658(2003)084[1608:PAHAPO]2.0.CO;2 |
| [70] |
Huang Y, Dai Q, Chen Y Y, Wan H F, Li J T, Wang Y Z. Lizard species richness patterns in China and its environmental associations. Biodiversity and Conservation, 2011, 20(7): 1399-1414. DOI:10.1007/s10531-011-0033-0 |
| [71] |
郝文芳, 杜峰, 陈小燕, 梁宗锁. 黄土丘陵区天然群落的植物组成、植物多样性及其与环境因子的关系. 草地学报, 2012, 20(4): 609-615. |
| [72] |
毕伟娜. 哈尔滨市蔷薇科植物景观与生态因子相关性的研究[D]. 哈尔滨: 东北林业大学, 2001.
|
| [73] |
陈宝瑞, 朱玉霞, 张宏斌, 周磊, 辛晓平. 呼伦贝尔草甸草原植被的数量分类和排序研究. 武汉植物学研究, 2008, 26(5): 476-481. DOI:10.3969/j.issn.2095-0837.2008.05.009 |
| [74] |
Keith M, Warren M. Can anthropogenic variables be used as threat proxies for south African plant richness?. Bothalia, 2007, 37(1): 79-88. DOI:10.4102/abc.v37i1.305 |
| [75] |
Andrew S M, Totland Ø, Moe S R. Spatial variation in plant species richness and diversity along human disturbance and environmental gradients in a tropical wetland. Wetlands Ecology and Management, 2015, 23(3): 395-404. DOI:10.1007/s11273-014-9390-2 |