 2022, Vol. 42
2022, Vol. 42文章信息
- 张喜亭, 张建宇, 肖路, 陈胜仙, 仲召亮, 高薇, 王文杰
- ZHANG Xiting, ZHANG Jianyu, XIAO Lu, CHEN Shengxian, ZHONG Zhaoliang, GAO Wei, WANG Wenjie
- 大兴安岭多布库尔国家级自然保护区植物多样性和群落结构特征
- Characteristics of plant diversity and community structure in Duobukuer Nature Reserve in Daxing'anling area
- 生态学报. 2022, 42(1): 176-185
- Acta Ecologica Sinica. 2022, 42(1): 176-185
- http://dx.doi.org/10.5846/stxb201812212775
-
文章历史
- 收稿日期: 2018-12-21
- 网络出版日期: 2021-08-16





2. 中国科学院东北地理与农业生态研究所, 长春 130102;
3. 大兴安岭行署资源管理局, 加格达奇 165000
2. Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun 130102, China;
3. Resource Management Bureau, Daxing'anling Administration Office, Jiagedaqi 165000, China
包括植物组成多样性在内的生物多样性是影响生态系统功能和服务发挥的关键因素之一, 对生态系统功能的可持续与生态服务功能的稳定性供应具有重要的影响, 也是森林生态系统管理与保护的重要组成部分[1]。有别于草地和农业生态系统, 群落特征及个体大小特征对评价森林生态功能具有更重要的作用, 合理的林分群落结构, 特别是垂直结构, 是森林发挥多种生态服务功能的基础[2]。我国已经把所有1.2×108 hm2天然林全部纳入了天然林保护工程[3], 探究天然林群落结构特征变化与植物多样性的关系, 为增强天然林保护的有效性采取管理措施提供合理建议具有重要的实践意义和科学意义。
大兴安岭是我国最北、面积最大的国有林区, 该地区具有独特的植物群落组成和生物多样性。大兴安岭地区是国家天然林保护工程的重点区域, 在维持生物多样性、涵养水源、固碳等方面起着关键作用, 具有极为重要的生态意义和经济价值[4]。大兴安岭曾为国家木材资源提供重要保障, 天保工程实施后, 大兴安岭天然林已由以往的砍伐优势木获取利益转变为林下资源产品的利用为辅、保护森林资源为主的管理经营方式, 加强森林经营、提升生态服务功能显得尤为重要。有关本区生物多样性研究已在多样性指数描述[5-6]、物种数量[7]、植物多样性与区系地理[8-9]、植物多样性与环境梯度的关系[10-12]等方面取得较多进展, 但森林群落结构和植物多样性关系的研究较少。选择典型天然林分(如国家级自然保护区), 通过树种大小、垂直结构与建群种多度、多样性、丰富度等多角度探究天然林林分结构特征及其与植物多样性关系, 能够作为天然林保护的范例与基准, 为天然林保护工程实施区域为森林生态系统植物多样性保护、森林结构与功能维持及后续生态服务服务功能持续稳定供给提供有益借鉴与参考[13-15]。
基于此, 本研究选取黑龙江多布库尔国家级自然保护区为研究对象, 综合多样性相关的各种群落结构特征指标体系, 在明确不同层次间群落结构特征与多样性分布规律的基础上, 探究群落结构特征与植物多样性的耦合关系, 并对比大兴安岭历史数据与区域其他研究结果, 研究发现将为保护区制定森林经营策略、提高林分组成多样性和复杂性提供数据支撑。
1 研究地区和方法 1.1 研究区及研究地点概况黑龙江多布库尔国家级自然保护区处于北纬50°19′— 50°43′, 东经124°18′— 125°04′之间, 面积约为12.9万hm2, 海拔在400—600 m之间[16]。该区域属寒温带大陆性气候, 其气候特点是冬季寒冷漫长, 春秋季不明显, 夏季较短。年温差较大, 全年平均气温-1.3—2℃, 最低气温-45.4℃, 最高气温37.3℃。全年平均降水量为500 mm, 无霜期80—130 d。保护区属中低山丘陵地貌, 山势平缓, 平均坡度10°左右。土壤类型主要有棕色针叶林土为主, 暗棕壤面积较小, 草甸土、河滩森林土与沼泽土镶嵌分布。植物种类组成较为丰富, 主要乔木为落叶松(Larix gmelinii)、蒙古栎(Quercus mongolica)、黑桦(Betula dahurica)等。主要灌木为越桔(Vaccinium vitis-idaea)、笃斯越桔(Vaccinium uliginosum)、榛(Corylus heterophylla)等。草本以莎草科(Cyperaceae)、菊科(Compositae)等为主。
本研究于2017年8月在黑龙江多布库尔国家级自然保护区进行调查, 在综合考虑功能区(核心区、缓冲区和实验区)、生境类型(干生、中生和湿生生境)及林型差异(落叶松林、白桦落叶松林、桦树林和杂木林)的前提下采用随机分层抽样的方法共设置42块30 m×30 m的样方。按照功能区划分, 核心区、缓冲区和实验区样方数量分别为13、15和14;按照生境类型划分, 干生生境、中生生境和湿生生境样方数分别为12、16和14;按照不同林型划分, 落叶松林、白桦落叶松林、桦树林和杂木林样方数量分别为14、11、10和7。在每个乔木层样方内随机设置5块2 m×2 m灌木样方调查灌木层, 在灌木样方内随机设置5块1 m×1 m小样方用于调查草本层。
在乔木样方内对胸径≥2.5 cm的乔木进行每木检尺, 调查项目包括胸径、树高和枝下高, 并记录物种名。灌木层样方调查灌木的种类、株数、株高、冠幅、盖度及地径;胸径小于2.5 cm且高于50 cm的乔木树种在灌木层样方中同步记录其种类、地径和树高, 并记入更新层中。草本层样方记录草本植物种类、盖度、高度和相对多度。记录每个样地的坡位和坡向。
在植被调查的同时, 在每乔木层样方中对0—20 cm土层土壤进行五点(四角及中心)混合取样, 同一样地的土样混合成一个混合样, 放入土壤袋, 风干测定样地含水量, 过2 mm土壤筛, 去掉大土壤砂砾和植物碎屑后, 再用粉碎机研磨过0.25 mm土壤筛装瓶, 用于后期土壤养分(有机碳、全氮、全磷)测定。
1.2 土壤养分测定土壤有机碳用重铬酸钾油浴法, 全氮用半微量凯氏定氮法, 全磷采用氢氧化钠熔融—钼锑抗比色法测定。相关详细测定方法已经在本课题组以往研究中多次使用[17-19]。
1.3 植物多样性指数、丰富度指数和均匀度指数的计算分别计算每个样方乔木、灌木和草本的Simpson指数、Alatalo均匀度指数和物种丰富度。计算方法如下:
丰富度指数:
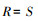
|
多样性指数[20]:
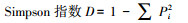
|
Alatalo均匀度指数[21]:
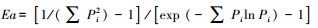
|
式中, Pi为样方内第i种的个体株数占总株数的比例;S为样方中物种总数。
在数据分析开始前, 本研究还计算了Shannon-Wiener指数、Pieolu均匀度指数, 这些指数与本文选择的Simpson多样性指数及Alatalo均匀度指数具有较高的相关性(R2>0.90), 为了避免所需指数的冗余, 因此本研究选择了以上3种指数来描述植物多样性特征。
1.4 个体大小特征和群落特征值的计算个体大小包括乔木层树高、枝下高、胸径, 灌木高度、地径和冠幅, 草本高度, 更新层树高, 更新层地径;群落特征指标包括乔木盖度、灌木盖度、草本盖度、乔木密度、灌木密度。所有特征指标均取平均值, 其中草本、更新和冠层层, 需要先计算每一样方的加权平均数, 以相对多度(或者树木、灌木株数)为权重, 再计算平行样方的平均值作为该样地的特征值。
1.5 数据分析与处理用Excel 2018饼状图方法表征乔灌草及更新层植物优势种相对多度;运用JMP 15.0频率分布分析, 确定平均值、中位数和分布范围等统计指标, 频率分布图相关图示可参考文献[22]。本文还用单峰型和多峰型表征频率分布图波动变化, 其中单峰型为正态分布或偏正态分布;多峰型为波动变化, 存在多个峰值。
采用Canoco 5.0软件分别进行群落特征、个体大小特征、土壤立地条件三者和乔木层、灌木层、草本层特征三者的方差分解分析。耦合关系采用RDA冗余排序分析, 具体分析过程中, 对15个林分特征因子和6个土壤立地条件因子(有机碳、全氮、全磷、含水量、坡度和坡向)进行前项选择, 每个因子重要性和显著性采用Monta-Carlo假设检验, 以P < 0.05作为显著性检验, 排除贡献较小的因子, 选择排名前10的解释变量进行分析。
2 结果与分析 2.1 乔、灌、草及更新层主要种相对多度多布库尔保护区乔木层中落叶松所占比例最大, 高达42%, 其次是白桦和黑桦。灌木层中, 榛所占比例高达60%, 占有绝对优势, 其次是胡枝子, 所占比例为18%。本研究中共记录到116种草本植物, 莎草科羊须草所占比例超过10%, 其次, 东方草莓(Fragaria orientalis)所占比例为6%, 地榆(Sanguisorba officinalis)、铃兰(Convallaria majalis)和小叶章(Deyeuxia purpurea)所占比例都为5%。更新层主要树种为蒙古栎, 所占比例高达54%;其次为白桦, 占更新层树种25%, 落叶松仅占6%(图 1)。

|
| 图 1 乔木层、灌木层、草本层和更新层主要种相对多度 Fig. 1 Relative abundance of main species in arbor layer, shrub layer, herb layer and regenerationlayer 图例按照物种占比由大到小顺序排列:落叶松Larix gmelinii白桦Betula platyphylla黑桦Betula dahurica蒙古栎Quercus mongolica山杨Populus davidiana稠李Padus racemosa兴安柳Salix hsinganica榛Corylus heterophylla胡枝子Lespedeza bicolor黑果茶藨Ribes nigrum刺玫蔷薇Rosa davurica毛榛Corylus mandshurica欧亚绣线菊Spiraea media羊须草Carex callitrichos东方草莓Fragaria orientalis地榆Sanguisorba officinalis铃兰Convallaria majalis小叶章Deyeuxia angustifolia北野豌豆Vicia ramuliflora兴安鹿蹄草Pyrola dahurica 关苍术Atractylodes japonica北方拉拉藤Galium boreale |
多布库尔保护区乔木丰富度均值为3.4, 最小50%集中区域丰富度为3—4;乔木层Simpson多样性指数频率分布也呈单峰型, 乔木层Simpson多样性平均值0.32, 但大部分样地的Simpson指数较小, 主要集中在0.2—0.3, 最小50%集中区域为0—0.32;该保护区的乔木层均匀度Alatalo指数平均值为0.6, 分布在0.5—0.6之间的样地最多(表 1)。
| 项目 Item |
平均值 Mean |
中位数 Median |
标准误差 Standard error |
25%中位数 25%Median |
75%中位数 75%Median |
最小50%区间 Minimum 50% range |
最大分布区间 Max interval |
分布区间 Distribution interval |
类型 Distribution type |
| 乔木丰富度Tree richness | 3.44 | 4 | 0.30 | 2 | 4 | 3—4 | 4—5 | 1—9 | 单峰 |
| 乔木Simpson指数Tree simpson | 0.32 | 0.31 | 0.05 | 0.05 | 0.53 | 0—0.32 | 0.2—0.3 | 0.07—0.76 | 单峰 |
| 乔木Alatalo指数Tree evenness | 0.60 | 0.56 | 0.04 | 0.46 | 0.73 | 0.52—0.74 | 0.5—0.6 | 0.31—0.94 | 单峰 |
| 灌木丰富度Shrub richness | 2.00 | 2 | 0.16 | 1 | 2.25 | 1—2 | 2—3 | 1—4 | 单峰 |
| 灌木Simpson指数Shrub simpson | 0.27 | 0.28 | 0.04 | 0.02 | 0.49 | 0—0.32 | 0—0.1 | 0—0.65 | 多峰 |
| 灌木Alatalo指数Shrub evenness | 0.75 | 0.72 | 0.04 | 0.58 | 0.93 | 0.53—0.78 | 0.7—0.8 | 0.40—1 | 多峰 |
| 草木丰富度Herb richness | 13.00 | 13 | 0.57 | 11 | 16 | 10—13 | 10—12.5 | 2.5—20 | 多峰 |
| 草木Simpson指数Herb simpson | 0.80 | 0.82 | 0.02 | 0.76 | 0.88 | 0.78—0.88 | 0.85—0.9 | 0.58—0.91 | 单峰 |
| 草木Alatalo指数Herb evenness | 0.65 | 0.66 | 0.02 | 0.56 | 0.73 | 0.62—0.73 | 0.65—0.7 | 0.41—0.88 | 单峰 |
| 更新丰富度Regenerated tree richness | 1.26 | 1 | 0.21 | 0 | 2 | 0—1 | 0—1 | 0—4 | 单峰 |
| 更新Simpson指数Regenerated simpson | 0.17 | 0 | 0.04 | 0 | 0.33 | — | 0—0.1 | 0—0.9 | 单峰 |
| 更新Alatalo指数Regenerated evenness | 0.50 | 0.56 | 0.07 | 0 | 1 | 0.58—1 | 0—0.2 | 0.45—1 | 多峰 |
| 乔木株高Tree height | 13.15 | 12.92 | 0.63 | 9.80 | 15.80 | 9.3—13.1 | 12—14 | 8.43—18.79 | 多峰 |
| 乔木胸径Tree diameter | 14.26 | 13.73 | 0.90 | 10.33 | 16.33 | 7.6—13.7 | 10—12.5 | 8.85—25.70 | 单峰 |
| 乔木枝下高Underbranch height | 7.50 | 6.51 | 0.67 | 4.54 | 9.99 | 3.3—7.2 | 4—6 | 2.78—14.42 | 单峰 |
| 灌木高度Shrub height | 1.06 | 1.10 | 0.08 | 0.66 | 1.50 | 1—1.70 | 1.50—1.75 | 0.29——1.91 | 多峰 |
| 灌木地径Shrub ground diameter | 1.09 | 1.04 | 0.05 | 0.92 | 1.27 | 0.80—1.08 | 0.75—1 | 0.63—1.97 | 多峰 |
| 灌木冠幅Shrub canopy width | 72.43 | 72.50 | 4.98 | 54.38 | 92.29 | 57.35—89.10 | 80—100 | 21.88—128.13 | 单峰 |
| 草本高度Herb height | 37.64 | 35.02 | 2.67 | 29.57 | 39.51 | 29.56—37.10 | 30—40 | 10.60—87.50 | 单峰 |
| 更新株高Regenerated tree height | 1.97 | 1.94 | 0.09 | 1.68 | 2.36 | 1.60—2.10 | 1.75—2 | 1.18—3.00 | 单峰 |
| 更新地径Regenerated tree diameter | 1.34 | 1.30 | 0.08 | 1.15 | 1.50 | 0.10—1.10 | 1.25—1.50 | 0.71—2.40 | 单峰 |
| 乔木郁闭度Tree canopy density | 0.73 | 0.70 | 0.02 | 0.70 | 0.80 | — | 0.7—0.75 | 0.50—0.90 | 单峰 |
| 灌木盖度Shrub coverage | 23.23 | 19.67 | 3.64 | 6.33 | 32.25 | 2.2—19.67 | 0—10 | 0—80 | 单峰 |
| 草本盖度Herb coverage | 54.36 | 53.67 | 2.71 | 46.33 | 65.50 | 48.33—62.37 | 50—60 | 4.66—86.33 | 单峰 |
| 乔木密度Tree density | 1500 | 1560 | 110 | 1177 | 1911 | 1177—1860 | 1000—1250 | 277—2666 | 多峰 |
| 灌木密度Shrub density | 12583 | 9583 | 1758 | 5833 | 14583 | 4100—11800 | 5000—10000 | 1666—39166 | 单峰 |
| 更新密度Regenerated tree density | 1723 | 500 | 208 | 0 | 2650 | 0—500 | 0—2000 | 0—11600 | 单峰 |
灌木层丰富度平均值为2.0, 最小50%集中区域为1—2;灌木层Simpson多样性指数平均值为0.27, 最大值为0.7;灌木层均匀度Alatalo指数平均值为0.75, 分布范围为0.4—1, 最小50%集中区域为0.53—0.78(表 1)。
草本层丰富度平均值为13, 分布区间为2.5—20, 最小50%集中区域为10—13;该保护区草本Simpson多样性平均值为0.80, 主要集中在0.85—0.9, 最小50%集中区域为0.78—0.88;草本均匀度指数平均值为0.65, 分布范围为0.4—0.9, 最小50%集中区域为0.62—0.73(表 1)。
更新层丰富度平均值为1.26, 有40%样方丰富度指数 < 1, 也就是没有发现更新层树种。更新层多样性指数平均值为0.17, 主要集中在0—0.1;更新层均匀度指数平均值为0.5(表 1)。
2.3 林分个体大小特征分布保护区内乔木树高平均值为13.15m, 分布区间在8—20m, 最小50%集中区域为9.3—13.1m;乔木胸径平均值为14.26cm, 最大分布区间在10—12.5cm之间;枝下高平均值为7.5m, 频率分布呈单峰型变化趋势, 分布范围在2.8—14.4m之间(表 1)。
灌木高度平均值为1.10m, 最小50%集中区域为1—1.7m;灌木地径平均值为在1.09cm, 最大分布区间为0.75—1cm, 最小50%集中区域为0.8—1.1cm;灌木冠幅平均值为72.43cm, 频率分布呈单峰型, 最小50%集中区域为57.3—89.1cm。
草本层高度平均值为37.6cm, 草本高度最大分布区间在30—40cm之间, 最小50%集中区域为39.5—37.1cm。
该保护区更新层树高均值为1.97m;地径平均值为1.34cm, 最大分布区间在1.25—1.50cm, 最小50%集中区域为0.10—1.10m。
2.4 林分群落特征分布从盖度的角度分析, 乔木郁闭度平均值为0.73, 呈单峰型变化趋势, 最大分布区间为0.7—0.75;灌木盖度平均值为23.23%, 分布区间在0—80%, 最小50%集中区域为2%—20%;草本层盖度平均值为54.36%, 频率分布图呈单峰型, 草本覆盖最高达86%, 最小50%集中区域为45%—62%(表 1)。
从密度的角度分析, 乔木层密度平均值为1500株/hm2, 频率分布呈多峰型变化趋势, 最小50%集中区域为1177—1860株/hm2;灌木层密度平均值为12583株/hm2, 大部分样地的灌木密度处于较低水平, 最小50%集中区域为4100—11800株/hm2;更新层密度平均值为1723株/hm2。
2.5 植物多样性变化的方差分解分析方差分解分析结果表明, 分为个体大小特征、群落特征和土壤立地条件3组时, 个体大小特征对多样性变化解释能力最大, 解释量为50.9%, 群落特征的解释量是个体大小特征的39%, 土壤养分和立地条件的解释量为29.6。分为乔木层、灌木层和草本层特征3组时, 乔木层的特征单独作用对多样性变化贡献最大, 解释量达到55.8%, 是灌木层的2倍, 草本层特征对多样性贡献较小, 解释量仅为6.9% (图 2)。

|
| 图 2 区分个体大小、群落特征与土壤立地条件和乔、灌、草的方差分解分析 Fig. 2 Variation partition between individual sizes, community characteristics and soil site conditions and among arbor layer, shrub layer and herb layer 图中简写的中文名称, TH:树高Tree height;TDBH:乔木胸径Tree diameter;TCBH:乔木枝下高Tree underbranch height;SH:灌高Shrub height;SCD:灌木冠幅Shrub canopy width;SGD:灌木地径Shrub ground diameter;HH:草高Herb height;RH:更新层株高Regenerated tree heightt;RGD:更新层地径Regenerated tree diameter;TC:乔木郁闭度Tree canopy density;SC:灌木盖度Shrub coverage;HC:草木盖度Herb coverage;TD:乔木密度Tree density;SD:灌木密度Shrub density;RD:更新层密度Regenerated tree density;MC:含水量Moisture;SLP:坡度Slope;ASP:坡向Aspect |
冗余分析表明, 个体大小、群落特征和土壤立地条件对植物多样性的总解释量为42.0%, 第一轴解释了多样性变化的16.49%, 第二轴解释了9.70%。简单效应结果表明, 乔木树高TH、枝下高TCBH和坡度SLP是多样性变化主要因子, 解释量分别为8.9%、7.7%和5.7%, P < 0.05;条件效应结果表明, 树高TH依然是对多样性变化贡献最大的, 解释量与单独作用相等, P < 0.01;其次灌木密度RD和乔木枝下高TCBH, P < 0.05(表 2)。
| 简单效应Simple Term Effects | 条件效应Conditional Term Effects | |||||||
| 项目Item | 解释量Explains/% | F | P | 项目Item | 解释量Explains/% | F | P | |
| TH | 8.9 | 3.4 | 0.004 | TH | 8.9 | 3.4 | 0.004 | |
| TCBH | 7.7 | 2.9 | 0.010 | RD | 5.3 | 2.1 | 0.046 | |
| SLP | 5.7 | 2.1 | 0.034 | SD | 5.3 | 2.2 | 0.046 | |
| SD | 4.9 | 1.8 | 0.102 | TCBH | 5.3 | 2.2 | 0.054 | |
| RD | 4.7 | 1.7 | 0.092 | TC | 4.1 | 1.8 | 0.118 | |
| SCD | 4.6 | 1.7 | 0.120 | SH | 3.5 | 1.6 | 0.124 | |
| SH | 4.4 | 1.6 | 0.128 | SCD | 3.3 | 1.5 | 0.148 | |
| HH | 4.2 | 1.6 | 0.146 | SGD | 2.2 | 1.0 | 0.418 | |
| SGD | 4.0 | 1.5 | 0.216 | HH | 2.2 | 1.0 | 0.432 | |
| TC | 3.5 | 1.3 | 0.266 | SLP | 2.1 | 1.0 | 0.384 | |
| 表中简写中文名称, TH:树高Tree height;TCBH:乔木枝下高Tree underbranch height;SH:灌高Shrub height;SCD:灌木冠幅Shrub canopy width;SGD:灌木地径Shrub ground diameter;HH:草高Herb height;TC:乔木郁闭度Tree canopy density;SD:灌木密度Shrub density;RD:更新层密度Regenerated tree density;SLP:坡度Slope | ||||||||
从不同林型样地分异散点图来看, 在第一轴上不同样地分化明显, 主要表现在落叶松林分布趋于第一轴的左边, 而杂木林则趋向于第一轴的右边方向上。落叶松林具有更高的树高、更大的直径, 而杂木林则具有更高的乔木树种多样性与丰富度, 但是个体多较小(图 3)。

|
| 图 3 群落结构特征与植物多样性的RDA排序、不同林型样地分布图和不同生境样地图 Fig. 3 RDA ordinations between species diversity and various factors, non-differentiation of sampling sites at 4 different forest types and non-differentiation of sampling sites at 3 different habitat 图中简写中文名称, T-R:乔木丰富度Tree richness;T-D:乔木Simpson指数Tree simpson;T-Ea:乔木均匀度Tree evenness;S-R:灌木丰富度Shrub richness;S-D:灌木Simpson指数Shrub simpson;S-Ea:灌木均匀度Shrub evenness;H-R:草本丰富度Herb richness;H-D:草本Simpson指数Herb simpson;H-Ea:草本均匀度Herb evenness;R-R:更新层丰富度Regenerated tree richness;R-D:更新层Simpson指数Regenerated tree simpson;R-Ea:更新层均匀度Regenerated tree evenness;B:白桦林Birch forest;L:落叶松林Larch forest;BL:白桦落叶松林Birch-Larch forest;S:杂木林Shaw;D:干生生境Dry habitat;M:中生生境Medium habitat;W:湿生生境Wet habitat |
从不同生境样地分异散点图来看, 本研究发现物种构成是连续变化的, 无法找到特定的界限明显的生境类型。在两个轴的方向上, 多布库尔保护区不同生境间并没有发生明显的分异。说明这三种生境类型具有相类似的多样性特征, 在耦合关系方面并没有产生差异(图 3)。
3 讨论 3.1 多布库尔自然保护区的植物多样性多布库尔自然保护区植物种类相对较为丰富, 大兴安岭地区代表性植物种类在本区均有分布[16]。多布库尔保护区湿地面积占保护区总面积的22.59%, 保护区内存在典型的沼泽化湿地, 这也是多布库尔保护区植物种类较为丰富的主要原因。2019年本课题组对多布库尔保护区植物本底调查记录了269种植物(未发表), 而本次调查中, 乔木记录了11种, 灌木11种, 草本总共记录116种, 超过保护区植物总数的50%。同时, 调查中发现保护区分布国家重点保护植物8种, 其中Ⅰ级保护植物3种, 为大花杓兰(Cypripedium macranthum)、杓兰(Cypripedium calceolus)、紫点杓兰(Cypripedium guttatum);国家Ⅱ级保护植物5种, 为钻天柳(Chosenia arbutifolia)、手参(Gymnadenia conopsea)、蒙古黄芪(Astragalus membranaceus)、草苁蓉(Boschniakia rossica)和野大豆(Glycine soja)。这也使多布库尔保护区的植物多样性的保护具有重要意义。
与位处大兴安岭核心区域的呼中国家级自然保护区相比, 多布库尔保护区乔木层丰富度(3.44)、草本层丰富度(13.00)分别是呼中保护区的1.5倍和1.6倍, 灌木层丰富度(2.00)相当于呼中保护区的40%[15]。所调查保护区样方共记录了11种灌木, 主要以喜温或耐旱的植物榛和胡枝子为主, 这体现了本保护区温带向寒温带过度的特点。目前针对保护区所在加格达奇林业局的研究主要集中在经济植物资源调查分析, 本研究对灌木的数据显示丰富度较低, 科学利用的同时也要注重多样性的保护。
本次多布库尔保护区更新层树种调查中, 蒙古栎占比高达为54%, 而落叶松仅占6%, 这反映出该保护区落叶松更新较差。蒙古栎一般分布在较为干旱的生境内, 更新层幼树以蒙古栎为主说明有向蒙古栎林演替的发展趋势。且灌层主要物种以耐旱植物为主, 应该是保护区旱生化特征明显, 特别是有杂木林的存在, 可能经历了多次采伐, 所以难能恢复到地带性顶极植被落叶松林, 而是向次生顶极蒙古栎林演变。
3.2 多布库尔保护区的群落结构特征多布库尔保护区乔木层平均树高13.2m, 平均枝下高为7.5m, 平均胸径为14.3cm。历史上长期对主要树种落叶松的采伐生产, 导致个体大的落叶松大量减少[23], 多布库尔乔木大小的数据也体现了这一点。多布库尔保护区乔木层树高较大兴安岭北部根河地区原始林乔木树高(10.52m)高了25.5%, 胸径较根河地区(13.39cm)高了6.80%[13]。历史资料表明, 1978年大兴安岭地区落叶松林树高均值为23.6m, 平均胸径为26.3cm[24], 这表明近四十年来森林资源的质量下降了45%, 平均每年降低1.1%, 森林生态服务功能退化堪忧。多布库尔保护区所在的加格达奇林业局在1988年、2002年的林业资源调查报告对比显示, 阔叶林面积比重逐年增大, 而且存在大量蒙古栎树种[25], 这与本研究调查结果一致。
多布库尔保护区林分郁闭度(0.73)高于整个加格达奇林业局的郁闭度:2002年和2012年加格达奇林业局二类调查郁闭度均值分别为0.55和0.58[26]。从本研究调查结果来看, 保护区尽管存在一些典型原始落叶松林, 但树木个体依然较小, 幼龄林是林分主体。天然林保护工程实施二十多年来, 大兴安岭地区森林资源保护方面取得了重大成果, 乱砍滥伐被有效遏制, 森林资源得以休养生息。从本研究结果来看, 多布库尔保护区郁闭度已经有所恢复。此外, 本研究在调查中发现保护区内存在部分火烧迹地, 这也为火烧迹地恢复及更新情况提供了数据支撑。
3.3 植物多样性影响途径分析与群落结构和土壤立地条件比较来说, 多布库尔保护区林分个体大小对多样性变化贡献最大, 个体大小影响效应是群落结构的2.5倍。本研究突出个体大小在保护植物种类多样性方面的重要性, 乔木、灌木的个体大小的调整往往伴随着植物多样性与均匀性的同步变化。保护区乔木层树高是影响多样性变化的最显著因子。关于乔木层特征对植物多样性的影响研究很多, 孔令伟等[27]在落叶松林中发现, 乔木层郁闭度、胸径、树高等生长指标对植物多样性的影响程度逐步降低;较低密度林分有助林下植物多样性的提高与恢复[28-29]。这些研究突出林分郁闭度和密度等通过光照对多样性的影响。与前人研究的区别在于, 多布库尔保护区人工林面积较大, 平均林龄偏小, 这可能导致树高对于多样性变化的贡献明显。
林分的天然更新直接影响未来森林植物多样性、群落结构以及生态功能[30]。更新层的密度和草本植物多样性具有负相关, 草本层多样性过高, 伴随着更新层密度变低。促进乔木的天然更新, 需要管理草本高度和种类。本研究通过探究群落结构与植物多样性的耦合关系, 能明确有效提高物种多样性的途径。与周围区域相比, 国家级自然保护区受人为影响最小, 是一个区域的植物资源本底值。植物多样性与群落结构关系的揭示, 明确了有效提高物种多样性的途径。
4 结论本文从乔灌草和更新层等角度分析了大兴安岭多布库尔国家级自然保护区的植物多样性和群落结构特征分布情况。研究结果表明, 多布库尔保护区平均树高为13.2m, 平均胸径为14.3cm, 胸径和树高仅相当于20世纪70年代的45%左右, 尚需要更长的保护时间来恢复森林资源。植物多样性受个体大小和群落特征影响不同, 个体大小特征的独立效应对多样性变化贡献最大, 解释力达到50.9%;而乔、灌、草三层比较来看, 乔木层解释力最大(55.8%)。RDA排序分析得出乔木层树高是引起多样性变化的显著因子。相关研究结果为保护区生物多样性保护以及周边天然林保护工程实施区域的进一步科学管理提供了数据支撑。
| [1] |
Hooper D U, ChapinIII F S, Ewel J J, Hector A, Inchausti P, Lavorel S, Lawton J H, Lodge D M, Loreau M, Naeem S, Schmid B, Setälä H, Symstad A J, Vandermeer J, Wardle D A. Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecological Monographs, 2005, 75(1): 3-35. DOI:10.1890/04-0922 |
| [2] |
张建国, 段爱国, 童书振. 林分直径结构模拟与预测研究概述. 林业科学研究, 2004, 17(6): 787-795. DOI:10.3321/j.issn:1001-1498.2004.06.017 |
| [3] |
杜红居, 张建宇, 仲召亮, 王文杰. 凉水国家自然保护区及毗邻区大型真菌资源调查. 安徽农业科学, 2017, 45(4): 1-5. DOI:10.3969/j.issn.0517-6611.2017.04.001 |
| [4] |
Wang C K, Gower S T, Wang Y H, Zhao H X, Yan P, Bond-Lamberty B P. The influence of fire on carbon distribution and net primary production of boreal Larix gmelinii forests in north-eastern China. Global Change Biology, 2001, 7(6): 719-730. DOI:10.1046/j.1354-1013.2001.00441.x |
| [5] |
任丽娟, 李钢铁, 孙新杰, 于世平, 郑宪武, 岳永杰. 内蒙古大兴安岭绰尔兴安落叶松群落结构与物种多样性研究. 林业资源管理, 2013(3): 83-87, 118-118. DOI:10.3969/j.issn.1002-6622.2013.03.019 |
| [6] |
王飞, 张秋良, 马秀枝, 郭旭. 不同发育阶段兴安落叶松林下植被多样性特征. 沈阳农业大学学报, 2013, 44(6): 771-775. DOI:10.3969/j.issn.1000-1700.2013.06.010 |
| [7] |
周以良, 倪红伟, 周瑞昌. 大兴安岭森林植物多样性特征. 国土与自然资源研究, 1998(3): 66-68. DOI:10.3969/j.issn.1003-7853.1998.03.016 |
| [8] |
萨如拉, 白学良, 毕庚辰, 福英. 大兴安岭南部山地苔藓植物区系多样性研究. 西北植物学报, 2013, 33(6): 1224-1233. DOI:10.3969/j.issn.1000-4025.2013.06.022 |
| [9] |
梁存柱, 王炜, 刘钟龄, 刘书润. 大兴安岭南部山地植物区系多样性研究. 内蒙古大学学报(自然科学版), 1997, 28(4): 553-562. |
| [10] |
孙菊, 李秀珍, 王宪伟, 吕久俊, 李宗梅, 胡远满. 大兴安岭冻土湿地植物群落结构的环境梯度分析. 植物生态学报, 2010, 34(10): 1165-1173. DOI:10.3773/j.issn.1005-264x.2010.10.005 |
| [11] |
孙菊, 李秀珍, 胡远满, 王宪伟, 吕久俊, 李宗梅, 陈宏伟. 大兴安岭沟谷冻土湿地植物群落分类、物种多样性和物种分布梯度. 应用生态学报, 2009, 20(9): 2049-2056. |
| [12] |
赵淑清, 方精云, 朴世龙, 宗占江, 吴晓莆, 古陶. 大兴安岭呼中地区白卡鲁山植物群落结构及其多样性研究. 生物多样性, 2004, 12(1): 182-189. DOI:10.3321/j.issn:1005-0094.2004.01.022 |
| [13] |
崔佳佳, 铁牛. 大兴安岭北部森林群落结构及植物多样性特征研究. 西北林学院学报, 2021, 36(02): 24-30. DOI:10.3969/j.issn.1001-7461.2021.02.04 |
| [14] |
Maltamo M, Kangas A, Uuttera J, Torniainen T, Saramäki J. Comparison of percentile based prediction methods and the weibull distribution in describing the diameter distribution of heterogeneous scots pine stands. Forest Ecology and Management, 2000, 133(3): 263-274. DOI:10.1016/S0378-1127(99)00239-X |
| [15] |
张建宇, 王文杰, 杜红居, 仲召亮, 肖路, 周伟, 张波, 王洪元. 大兴安岭呼中地区3种林分的群落特征、物种多样性差异及其耦合关系. 生态学报, 2018, 38(13): 4684-4693. |
| [16] |
纪鹰翔. 大兴安岭林下资源现状及开发利用分析——以黑龙江多布库尔自然保护区为例. 林业科技情报, 2016, 48(1): 4-6. |
| [17] |
Wang W J, Wang Q, Zhou W, Xiao L, Wang H M, He X Y. Glomalin changes in urban-rural gradients and their possible associations with forest characteristics and soil properties in Harbin City, Northeastern China. Journal of Environmental Management, 2018, 224: 225-234. |
| [18] |
Wang Q, Wang W J, He X Y, Zhou W, Zhai C, Wang P J, Tang Z, Wei C H, Zhang B, Xiao L, Wang H Y. Urbanization-induced glomalin changes and their associations with land-use configuration, forest characteristics, and soil properties in Changchun, Northeast China. Journal of Soils and Sediments, 2019, 19(5): 2433-2444. DOI:10.1007/s11368-019-02266-x |
| [19] |
Wang Q, Zhang D, Zhou W, He X Y, Wang W J. Urbanization led to a decline in glomalin-soil-carbon sequestration and responsible factors examination in Changchun, Northeastern China. Urban Forestry & Urban Greening, 2020, 48: 126506. |
| [20] |
张涵丹, 康希睿, 邵文豪, 杨旭, 张建锋, 刘学全, 陈光才. 不同类型杉木人工林林下草本植物多样性特征. 生态学报, 2021, 41(06): 2118-2128. |
| [21] |
Hurlbert S H. The nonconcept of species diversity: a critique and alternative parameters. Ecology, 1971, 52(4): 577-586. DOI:10.2307/1934145 |
| [22] |
张波, 王文杰, 周伟, 肖路, 吕海亮, 魏晨辉. 哈尔滨城市森林树木生长状况及各生长指标的相关性分析. 安徽农业科学, 2016, 44(26): 127-128, 154-154. DOI:10.3969/j.issn.0517-6611.2016.26.044 |
| [23] |
Guo Y, Lu J M, Franklin S B, Wang Q G, Xu Y Z, Zhang K H, Bao D C, QiaoX J, Huang H D, Lu Z J. Spatial distribution of tree species in a species-rich subtropical mountain forest in central China. Canadian Journal of Forest Research, 2013, 43(9): 826-835. DOI:10.1139/cjfr-2013-0084 |
| [24] |
李永武, 陈新华, 胡沐钦. 关于大兴安岭林区落叶松成过熟林生长和枯损的初步研究. 林业资源管理, 1979(1): 38-43. |
| [25] |
周宏, 唐志强, 刘福. 加格达奇林业局森林资源消长变化分析. 内蒙古林业调查设计, 2004, 27(1): 39-41, 46-46. |
| [26] |
康文智, 于顺龙, 栾长江. 加格达奇林业局森林资源动态变化分析. 内蒙古林业调查设计, 2015, 38(2): 14-16. |
| [27] |
孔令伟, 陈祥伟, 鲁绍伟, 李少宁, 陈波, 高琛, 石媛, 杨小燕. 华北落叶松林木生长、草本植物多样性及地形因子之间的关系. 水土保持通报, 2014, 34(5): 60-66. |
| [28] |
张勇强, 李智超, 厚凌宇. 林分密度对杉木人工林下物种多样性和土壤养分的影响. 土壤学报, 2020, 57(1): 239-250. |
| [29] |
王媚臻, 毕浩杰, 金锁, 刘佳, 刘宇航, 王宇, 齐锦秋, 郝建锋. 林分密度对云顶山柏木人工林林下物种多样性和土壤理化性质的影响. 生态学报, 2019, 39(3): 981-988. |
| [30] |
金红喜, 杨占彪, 袁彩霞, 何小琴, 王刚. 六盘山4种类型森林群落天然更新初探. 西北林学院学报, 2009, 24(01): 93-97, 135-135. |