 2021, Vol. 41
2021, Vol. 41文章信息
- 李键, 刘鑫铭, 姚成硕, 宫欢欢, 吴承祯, 范海兰
- LI Jian, LIU Xinming, YAO Chengshuo, GONG Huanhuan, WU Chengzhen, FAN Hailan
- 武夷山国家公园不同林地土壤呼吸动态变化及其影响因素
- Dynamic change of soil respiration and its effecting factors in different forest lands of Wuyishan National Park
- 生态学报. 2021, 41(9): 3588-3602
- Acta Ecologica Sinica. 2021, 41(9): 3588-3602
- http://dx.doi.org/10.5846/stxb201905130981
-
文章历史
- 收稿日期: 2019-05-13
- 网络出版日期: 2021-03-08





2. 福建省高校森林生态系统经营与过程重点实验室, 福州 350002;
3. 武夷学院生态与资源工程学院, 南平 354300;
4. 福建省生态产业绿色技术重点实验室, 南平 354300
2. Key Laboratory of Forest Ecosystem and Management in Fujian Colleges, Fuzhou 350002, China;
3. College of Ecology and Resources Engineering, Wuyi College, Nanping 354300, China;
4. Key Laboratory for Eco-Industrial Green Technology of Fujian Provincial, Nanping 354300, China
土壤呼吸作为土壤碳的主要输出途径和大气CO2重要来源, 是陆地生态系统的第二碳通量, 在调控全球生态系统的碳循环和气候动态方面起着至关重要的作用[1-3]。近年来, 土壤呼吸及其影响因素已成为生态学研究热点[4], 气候变化、植被类型、土壤理化性质[5-6]、土壤微生物群落[7-8]等因素均对土壤呼吸产生影响[9]。温度[10]和水分[11]能够直接或通过影响土壤微生物和地表植物间接影响土壤呼吸。研究表明土壤呼吸速率与温度呈显著正相关[12-13], 温度是土壤碳排放变异的主要影响因素[14]。目前研究大多集中于温度和湿度对土壤呼吸速率的影响, 温度和水分对土壤呼吸的协同作用可通过线性关系或幂-指数模型[15]表示。而土壤呼吸与而温度、水分、土壤理化性质等因素的交互作用与土壤呼吸的关系较为复杂[16], 相关研究较少。
武夷山作为中国第二大丹霞地貌区, 地貌类型独特, 生态环境复杂。随着社会发展和人类活动的增加, 许多林地被重新开垦利用, 土地利用现状总体呈现出林地分布广, 茶园面积大的特点[17]。不同的林地类型会改变地表植被覆盖特征、土壤理化性质、土壤微生物数量及种类等, 从而改变土壤呼吸过程, 对全球生态系统的气候变化和碳循环变化产生影响[18]。2015年, 武夷山风景名胜区、自然保护区等地被国家发改委整合确定为我国首批国家公园体制试点之一, 建立以国家公园为主体的自然保护体系。由于风景区土地利用类型较为复杂, 对于自然保护地体系的生态平衡可能造成影响, 因此研究不同林地的土壤呼吸动态变化, 对于准确评价和预测区域碳收支具有重要的意义。目前, 许多学者对武夷山地区土壤呼吸的研究主要集中在不同海拔对土壤呼吸的影响[19-20], 对于不同林地利用方式下土壤呼吸差异的研究较少。本文通过野外定位研究和室内分析相结合的方法, 选取立地条件基本一致的裸地、茶园、锥栗(Castanea henryi (Skam) Rehd. et Wils.)林和马尾松(Pinus massoniana Lamb.)林为研究对象, 采用Li—8100开路式土壤碳通量测定系统, 测量这4种典型林地的土壤呼吸速率日动态变化和季节动态变化, 并同步测定林地内的近地面气温、土壤温度、土壤湿度、土壤养分和微生物碳、氮含量。旨在探索以下问题: (1)林地土壤呼吸的日动态和季节动态变化规律以及不同类型林地之间土壤呼吸的差异性;(2)不同类型林地的土壤呼吸与温度、湿度、土壤养分等因素的关系。以期为林地利用方式变化背景下武夷山国家公园生态系统碳循环研究和生态系统保护工作提供一定的数据支持和参考借鉴。
1 材料与方法 1.1 研究区概况武夷山国家公园位于福建省西北部, 地处东经117°37′22″—118°19′44″, 北纬27°27′31″—28°04′49″, 属于中亚热带季风气候(图 1)。春夏暖湿多雨, 秋冬干燥少雨, 年平均气温19.7℃, 年平均降雨量1960 mm。土壤质地为沙质砾土, 土质疏松、土层浅薄、有机质贫乏。植被类型为亚热带常绿阔叶林。不同林地基本概况及土壤基本理化性质均在2016—2017年测定, 结果见表 1。于研究区分别选取土地利用前林地条件基本一致的锥栗林(10 a)、茶园(10 a)、马尾松林(10 a)等3种林地利用方式, 裸地为对照。

|
| 图 1 研究区地图 Fig. 1 Research area map |
| 林地Forest land | 林龄Stand age/a | pH | TC/(g/kg) | TN/(g/kg) |
| 茶园Tea garden | 10 | 4.53±0.07 | 6.69±0.45 | 1.52±0.03 |
| 马尾松Pinus massoniana forest | 10 | 4.42±0.23 | 5.84±0.62 | 1.10±0.07 |
| 锥栗Castanea henryi forest | 10 | 4.49±0.14 | 8.34±0.74 | 1.64±0.07 |
| 裸地Bare land | — | 4.92±0.28 | 4.39±0.46 | 0.86±0.09 |
| TC: 全碳Total Carbon; TN: 全氮Total Nitrogen | ||||
裸地, 27°40′8″N, 117°54′48″E, 海拔238 m, 坡度22°, 东北坡, 坡中部。地表大部分土壤裸露, 仅零星丛生鬼针草(Bidens pilosa L.)、野艾(Artemisia lavandulaefolia)、牛筋草(Eleusine indica (L.) Gaertn.)等杂草。
锥栗林, 27°40′42″N, 117°54′54″E, 海拔221 m, 坡度20°, 东北坡, 坡中部, 平均树高8.5 m。平均冠幅1.09 m×0.76 m, 郁闭度0.6。林下植被稀少, 盖度约3%, 主要有菝葜(Smilax china L.)、小飞蓬(Conyza Canadensis (L.) Cronq.)、乌桕(Sapium sebiferum (L.) Roxb.)等。
茶园, 27°40′19″N, 117°55′18″E, 海拔214 m, 坡度16°, 东北坡, 平均树高0.58 m。林下植被稀少, 每年有定期的人工管理。2月下旬进行中耕(15 cm左右)、施肥和轻修剪, 5月下旬进行浅耕(10 cm左右)、施肥、除草和重修剪, 7月下旬进行浅耕(10 cm左右)和施肥, 8月进行除草, 11月上旬进行深耕(20 cm左右)、施肥、除草和轻修剪, 具体施肥情况见表 2。
| 时间Time | 形式Form | 种类Type | 用量Amount/(kg/667m2) |
| 11月上旬Early November | 基肥 | 有机肥 | 30 |
| 2月下旬Late February | 追肥 | 复合肥 | 10 |
| 5月下旬Late May | 追肥 | 复合肥、尿素 | 20、10 |
| 7月下旬Late July | 追肥 | 复合肥 | 10 |
马尾松林, 27°40′57″N, 117°55′32″E, 海拔230 m, 坡度17°, 东北坡, 平均树高6.11 m。平均冠幅1.02 m×0.53 m, 郁闭度0.4, 林下植被盖度约80%。主要灌木有檵木(Loropetalum chinense (R. Br.) Oliver)、桃金娘(Rhodomyrtus tomentosa)、乌饭(Vaccinium bracteatum Thunb.)、山矾(Symplocos sumuntia Buch.-Ham. ex D. Don)、红叶石楠(Photinia × fraseri Dress)、金樱子(Rose Laevigata Michx.)等, 另有少量盐肤木(Rhus chinensis Mill.)、菝葜(Smilax china L.)、山莓(Rubus corchorifolius L. f.)、苦竹(Pleioblastus amarus (Keng) keng)、桂花(Osmanthus fragrans (Thunb.) Lour.)、箬竹(Indocalamus barbatus (Munro) Keng f.)、山茶(Camellia japonica L.)、木荷(Schima superb Gardn. et Champ.), 在灌木层中还混生有栲属、青冈属等乔木树种的各级幼苗。草本以铁芒萁(Dicranopteris linearis)为主, 占90%, 另有少量乌蕨(Stenoloma chusanum (L.) J. Sm.)、油芒(Eccoilopus cotulifer (Thunb.) A. Camus)、五节芒(Miscanthus floridulu (Lab.) Warb. ex Schum. et Laut.)、胜红蓟(Ageratum conyzoides L.)。
1.2 样点布置与测定方法2016年9月, 在裸地、茶园(10 a)、锥栗林(10 a)、马尾松林(10 a)内分别随机选取面积为20 m×20 m的样地, 并在每个样地内随机设置5个采样点, 在每个采样点上布置PVC土壤环用于测量土壤呼吸。将一端削尖、直径16 cm、高11.5 cm的PVC土壤环压入土中, 使其插入地下5.5 cm, 地上5 cm。同时将土壤环内部的杂物清理干净, 外围的土踩实, 在整个过程中尽量不扰动周围环境。土壤环埋置完成后, 至少静置24 h后才能进行土壤呼吸测量, 并且每次测量前, 提前一天清除土壤环内的杂物。
采用Li—8100开路式土壤碳通量测定系统, 于2016年10月(秋季)、2017年1月(冬季)、2017年4月(春季)、2017年7月(夏季)选择相对晴好的天气, 分别对4个林地进行土壤呼吸速率(Soil respiration rate, Rs)测定。从6:00至18:00, 每隔2 h对每个样点进行逐一测定, 每个样点测定3个重复取平均值, 每次连续测定一个星期。以每个时间点所测量的林地内5个样点土壤呼吸速率的平均值作为该时间点下林地的土壤呼吸速率并形成日动态。以每个季节土壤呼吸速率日变化的数据取均值作为该季节土壤呼吸的季节均值。土壤温度(0—10 cm)和土壤湿度(0—10 cm)由Li—8100便携式土壤呼吸测定仪自带的温度和含水量传感器测定, 近地面(地上1.5 m处)气温用便携式温度计在原位测量土壤呼吸的同时测量, 下文所提的土壤湿度即土壤容积含水率。实际的具体测量时间为: 2016年10月11日—17日, 2017年1月5日—11日, 2017年4月2日—8日, 2017年7月16日—24日。
测量土壤呼吸的同时, 以“三点取样法”随机选择在三个土壤环附近设置样点进行土壤取样, 采集0—10 cm、10—20 cm、20—30 cm的土壤样品后将其密封于袋中带回, 一份于室内实验室自然风干后用于土壤养分测定(其中土壤有效养分的测定用风干后研磨过2 mm筛的土样, 土壤全量养分的测定用风干后研磨过0.149 mm筛的土样), 另一份放于4℃冰箱保存用于土壤微生物碳、氮测定。土壤全碳(Total carbon, TC)、全氮(Total nitrogen, TN)采用碳氮元素分析仪(Elementar, 德国)测定, 水解氮(Hydrolyzed nitrogen, HN)采用碱解扩散法, 全磷(Total phosphorus, TP)采用碱熔—钼锑抗比色法, 有效磷(Available phosphorus, AP)采用碳酸氢钠浸提—钼锑抗比色法, 全钾(Total potassium, TK)采用熔—火焰光度法, 速效钾(Available potassium, AK)采用乙酸铵浸提—火焰光度法[21]。参考相关方法[22-23]测定土壤微生物生物量碳、氮含量, 采用氯仿熏蒸法对样品进行处理, 采用硫酸—重铬酸钾外加热法测定微生物生物量碳(Microbial biomass carbon, MBC), 采用硫酸—半微量凯氏定氮法测定微生物生物量氮(Microbial biomass nitrogen, MBN)。
1.3 数据分析与模型选择运用Excel 2016对数据进行处理和作图, 运用SPSS 17.0软件对不同林地不同季节土壤呼吸速率的差异及不同林地不同土层土壤养分的差异进行方差分析(LSD, P<0.05), 对4种林地的土壤呼吸速率及水热因子(近地面气温、土壤温度、土壤湿度)进行多元回归分析, 并采用多元线性回归(Enter、Stepwise)、对数模型和幂-指数模型分别对4种林地的土壤呼吸和水热因子之间的关系进行模型拟合。以四季0—30 cm土层的土壤养分含量均值作为林地的土壤养分数据, 对不同林地土壤养分含量和土壤呼吸速率进行Pearson相关性分析, 并根据不同林地土壤呼吸速率和土壤养分含量的相关性, 利用逐步回归(Stepwise)法构造出最优多元回归模型。
对数模型[24]:
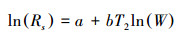
|
(1) |
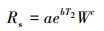
|
(2) |
式中, Rs为土壤呼吸速率(μmol CO2 m-2 s-1), T2为土壤温度(℃), W为土壤湿度(%), a、b、c均为待定系数。
2 结果与分析 2.1 不同林地土壤呼吸的动态变化茶园、马尾松、锥栗、裸地的土壤呼吸速率从6:00—18:00均呈现先增后减趋势, 在14:00达到峰值。锥栗、茶园和马尾松的土壤呼吸速率变化幅度较大, 而裸地变化幅度较小, 一天内土壤呼吸速率的极差为0.5 μmol m-2 s-1左右(图 2)。4种林地土壤呼吸速率的季节均值按大小排序为茶园(3.10 μmol m-2 s-1)>马尾松(2.96 μmol m-2 s-1)>锥栗(2.32 μmol m-2 s-1)>裸地(1.43 μmol m-2 s-1)。4种林地土壤呼吸速率的差异在夏季最大, 茶园的土壤呼吸速率(4.85 μmol m-2 s-1)是裸地(1.73 μmol m-2 s-1)的近3倍(图 3)。

|
| 图 2 不同林地土壤呼吸的日变化 Fig. 2 Daily variation of soil respiration in different forest land |

|
| 图 3 不同林地土壤呼吸的季节变化 Fig. 3 Seasonal variation of soil respiration in different forest land 不同大写字母表示相同林地不同季节差异显著(P<0.05), 不同小写字母表示同一季节不同林地差异显著(P<0.05) |
不同林地近地面气温、土壤温度和土壤湿度的日变化规律(图 4、5)与土壤呼吸速率(图 2)基本一致, 均呈现明显的单峰曲线规律。相同时间下同一林地的土壤温度比近地面气温略低, 夏季的差异最大。相同季节下, 14:00前后裸地的近地面气温和土壤温度高于其他三种林地。

|
| 图 4 不同林地近地面气温的日变化 Fig. 4 Daily variation of near surface air temperature in different forest land |

|
| 图 5 不同林地土壤温度和土壤湿度的日变化 Fig. 5 Daily variation of soil temperature and soil moisture in different forest land |
不同林地的近地面气温、土壤温度和土壤湿度均表现为夏季>春季>秋季>冬季。同一季节, 裸地的近地面气温和土壤温度在4种林地中最高。春、夏季各林地中锥栗林的近地面气温、土壤温度和土壤湿度均为最低, 而秋、冬季各林地的差异不大(图 6)。

|
| 图 6 不同林地近地面气温、土壤温度和土壤湿度的季节变化 Fig. 6 Seasonal variation of near surface air temperature, soil temperature and soil moisture in different forest land |
同一季节, 各林地不同土层之间的养分含量差异显著, 且随土层增加呈现不同的规律性。各林地的土壤TC、TN、HN、TP、AP含量均随土层的增加而减少, TK含量随土层的增加而增加, 茶园、锥栗林和马尾松林的AK含量随土层的增加而减少, 裸地则相反。各林地的TC含量在各土层之间的差异最大, 裸地的TN、HN、TP、AP和AK含量在各土层之间差异不明显。其中秋季茶园0—10 cm与20—30 cm的TC含量差异(9.43 g/kg)最大(表 3)。
| 季节 Season |
林地类型 Forest type |
土层 Soil layer/cm |
全碳 TC/(g/kg) |
全氮 TN/(g/kg) |
水解氮 HN/(mg/kg) |
全磷 TP/(g/kg) |
有效磷 AP/(mg/kg) |
全钾 TK/(g/kg) |
速效钾 AK/(mg/kg) |
| 春季Spring | 茶园Tea garden | 0—10 | 9.47±2.23Ba | 2.00±0.10Ba | 76.49±2.13Aa | 0.81±0.04Aa | 60.41±2.99Aa | 1.63±0.08Ba | 50.16±2.86Aa |
| 10—20 | 6.40±1.47Bb | 1.87±0.06Bab | 44.32±8.07Ab | 0.69±0.05Ab | 37.49±3.91Bb | 1.63±0.02Ba | 46.70±4.49Aa | ||
| 20—30 | 3.97±0.46Bb | 1.69±0.15Bb | 34.05±6.99Ab | 0.60±0.05Ab | 31.63±3.93Ab | 1.71±0.21BCa | 45.83±2.37ABa | ||
| 马尾松 | 0—10 | 10.17±0.49Ba | 1.29±0.24Ca | 34.99±4.01Ba | 0.52±0.02Ba | 10.08±1.04Ca | 1.62±0.06Ba | 40.82±0.64Ba | |
| Pinus massoniana forest | 10—20 | 6.57±0.97Bb | 1.18±0.10Ca | 28.45±5.82Bab | 0.52±0.02Ba | 9.57±0.94Ba | 1.63±0.15Ba | 39.44±1.39Aa | |
| 20—30 | 3.40±1.22Bc | 1.13±0.06Ca | 23.32±4.04Bb | 0.50±0.01Ba | 9.02±0.3Ba | 1.56±0.07Ca | 35.53±1.32Cb | ||
| 锥栗 | 0—10 | 14.73±2.00Aa | 3.17±0.42Aa | 71.87±8.51Aa | 0.61±0.05Ba | 20.04±0.97Ba | 1.70±0.06Bb | 46.50±4.63Aa | |
| Castanea henryi forest | 10—20 | 11.67±1.75Aab | 2.61±0.42Aab | 46.17±2.79Ab | 0.56±0.09Ba | 12.66±1.24Ab | 1.78±0.05Ba | 42.14±4.51Aa | |
| 20—30 | 9.43±1.10Ab | 2.17±0.21Ab | 28.93±6.31ABc | 0.51±0.06Ba | 10.48±1.03Bb | 1.84±0.13Ba | 42.23±2.15Ba | ||
| 裸地Bare land | 0—10 | 9.4±1.48Ba | 1.16±0.21Ca | 29.86±8.43Ba | 0.52±0.06Ba | 10.34±0.5Ca | 2.35±0.34Ab | 40.57±3.96Bb | |
| 10—20 | 4.10±1.40Bb | 0.82±0.11Cb | 22.39±5.60Ba | 0.49±0.05Ba | 9.7±0.26Ba | 2.62±0.06Aa | 43.52±1.77Aab | ||
| 20—30 | 3.33±1.00Bb | 0.77±0.09Db | 20.53±3.53Ba | 0.44±0.02Ba | 9.52±0.42Ba | 2.68±0.09Aa | 47.93±3.23Aa | ||
| 夏季Summer | 茶园 | 0—10 | 9.47±2.04Aa | 1.30±0.20ABa | 62.97±8.38Ba | 0.73±0.07ABa | 50.39±3.72Aa | 1.46±0.15Ca | 55.27±3.82Aa |
| 10—20 | 8.23±1.63Aa | 1.22±0.11Aab | 48.51±5.65Ab | 0.56±0.14Aa | 36.98±2.26Ab | 1.59±0.10Ba | 49.19±3.78Ab | ||
| 20—30 | 3.60±0.75ABb | 0.90±0.17Ab | 28.45±3.76Ac | 0.50±0.15Aa | 21.62±2.96Ac | 1.61±0.04Ba | 48.49±3.02Bb | ||
| 马尾松 | 0—10 | 8.60±1.77ABa | 1.01±0.32Ba | 42.93±6.31Ca | 0.50±0.01Ba | 10.47±1.47Ca | 1.62±0.05BCa | 38.61±3.38Ca | |
| 10—20 | 5.97±1.68ABab | 0.90±0.10Ba | 34.06±4.65Bab | 0.49±0.01Aa | 9.99±0.63Ca | 1.68±0.05Ba | 36.08±4.19Ba | ||
| 20—30 | 4.43±0.78Ab | 0.76±0.06Aa | 25.66±3.52Ab | 0.42±0.09Aa | 9.58±0.26Ca | 1.68±0.02Ba | 34.88±3.12Ca | ||
| 锥栗 | 0—10 | 11.70±1.40Aa | 1.63±0.29Aa | 97.01±7.16Aa | 0.55±0.02Ba | 26.8±3+.03Ba | 1.67±0.07Ba | 49.76±1.60ABa | |
| 10—20 | 6.00±1.23ABb | 1.13±0.15ABb | 58.79±7.27Ab | 0.55±0.09Aa | 18.75±2.27Bb | 1.74±0.03Ba | 46.31±3.21Ab | ||
| 20—30 | 3.00±0.36Bc | 0.87±0.06Ab | 31.26±6.32Ac | 0.51±0.05Aa | 13.91±1.5Bb | 1.74±0.09Ba | 46.49±3.28Bab | ||
| 裸地 | 0—10 | 6.3±1.05Ba | 1.05±0.23Ba | 35.92±6.62Ca | 0.49±0.01Ba | 10.88±0.81Ca | 1.95±0.16Ab | 46.60±2.78Ba | |
| 10—20 | 4.03±0.84Bb | 0.93±0.15Ba | 27.53±5.83Ba | 0.45±0.01Ab | 10.06±0.65Cab | 2.48±0.36Aa | 50.03±4.25Aa | ||
| 20—30 | 3.10±0.53Bb | 0.92±0.10Aa | 14.32±1.52Bb | 0.43±0.03Ab | 9.61±0.38Cb | 2.47±0.22Aab | 53.06±3.27Aa | ||
| 秋季Autumn | 茶园 | 0—10 | 11.50±1.44Aa | 2.70±0.15Aa | 51.31±8.85Ba | 0.34±0.05Ba | 12.54±1.41Aa | 0.64±0.12Ba | 30.64±3.48Aa |
| 10—20 | 5.93±1.55ABb | 2.13±0.41Ab | 28.45±3.71Bb | 0.36±0.11Aa | 11.64±1.46Aa | 0.68±0.10Ba | 22.41±4.14Bb | ||
| 20—30 | 2.07±0.86Bc | 1.30±0.20Ac | 9.80±2.80Cc | 0.30±0.03Aa | 10.9±1.47Aa | 0.59±0.05Ca | 19.57±3.49BCb | ||
| 马尾松 | 0—10 | 8.63±1.04Ba | 1.73±0.51Ba | 37.78±3.51Ca | 0.39±0.04ABa | 11.44±0.66Aa | 0.64±0.15Ba | 22.78±0.85Ba | |
| 10—20 | 4.00±0.82ABb | 1.55±0.22Ba | 32.65±6.79Ba | 0.33±0.05Aa | 10.11±0.32Bab | 0.69±0.12Ba | 16.66±2.44Bb | ||
| 20—30 | 3.37±0.61ABb | 1.46±0.10Aa | 17.73±2.83Bb | 0.31±0.04Aa | 9.63±0.51Ab | 0.76±0.10Ba | 15.95±2.75Cb | ||
| 锥栗 | 0—10 | 12.30±2.60Aa | 2.33±0.26Aa | 81.63±7.49Aa | 0.51±0.15Aa | 12.68±1.05Aa | 0.77±0.04ABb | 24.48±3.58Ba | |
| 10—20 | 7.20±2.86Ab | 1.60±0.20Bb | 64.85±9.34Ab | 0.36±0.04Aa | 11.56±0.12Ab | 0.67±0.11Bb | 21.32±2.46Ba | ||
| 20—30 | 4.20±0.95Ab | 1.13±0.14Ac | 34.99±1.40Ac | 0.32±0.02Aa | 10.51±0.5Ab | 0.83±0.06Ba | 23.28±2.24Ba | ||
| 裸地 | 0—10 | 5.20±1.05Ca | 0.88±0.19Ca | 18.66±5.30Da | 0.35±0.06Ba | 11.3±0.55Aa | 0.92±0.09Ab | 25.66±3.68ABa | |
| 10—20 | 3.17±1.32Bb | 0.80±0.17Ca | 13.53±3.52Ca | 0.31±0.04Aa | 10.86±0.45ABa | 1.24±0.09Aa | 29.52±4.81Aa | ||
| 20—30 | 2.43±0.31Bb | 0.76±0.21Ba | 11.66±2.08Ca | 0.29±0.04Aa | 10.94±0.88Aa | 1.31±0.06Aa | 31.09±1.95Aa | ||
| 冬Winter | 茶园 | 0—10 | 11.00±0.89Aa | 1.43±0.15Aa | 84.91±10.60Aa | 0.49±0.10Ba | 13.98±0.17Aa | 0.68±0.05ABa | 32.07±2.48Aa |
| 10—20 | 5.43±1.23Ab | 0.94±0.05Ab | 55.50±9.00Ab | 0.34±0.02Ab | 12.6±0.15Ab | 0.77±0.04Ba | 26.80±2.87Ab | ||
| 20—30 | 3.17±1.10 Ac | 0.70±0.10ABc | 32.19±4.72Ac | 0.31±0.01Ab | 11.72±0.32Ac | 0.75±0.06Ba | 24.52±3.06Bb | ||
| 马尾松 | 0—10 | 10.23±2.53Aa | 1.10±0.35ABa | 33.59±1.39Ca | 0.33±0.05Aa | 10.85±0.28Ba | 0.58±0.13Ba | 21.80±3.91Ba | |
| 10—20 | 2.70±0.72Bb | 0.63±0.25Aab | 12.60±1.40Cb | 0.28±0.03Aa | 9.96±0.54Bb | 0.59±0.04Ba | 16.57±1.78Bab | ||
| 20—30 | 2.03±0.84ABb | 0.47±0.16Bb | 9.80±1.06Bc | 0.26±0.04Aa | 9.44±0.43Bb | 0.66±0.08Ba | 13.67±2.81Cb | ||
| 锥栗 | 0—10 | 11.40±0.85Aa | 1.33±0.21ABa | 66.25±4.96Ba | 0.37±0.06Aa | 10.94±1.73Ba | 0.62±0.04Ba | 30.87±4.36Aa | |
| 10—20 | 5.63±1.68Ab | 0.93±0.24Aab | 35.92±0.81Bb | 0.32±0.05Aa | 8.93±0.68Ca | 0.57±0.07Ba | 23.37±1.22Aa | ||
| 20—30 | 2.80±0.53ABc | 0.73±0.15Ab | 30.79±2.81Ab | 0.30±0.03Aa | 8.47±0.35Ca | 0.68±0.09Ba | 25.61±4.12Ba | ||
| 裸地 | 0—10 | 6.80±1.31Ba | 1.01±0.20Ba | 25.65±4.27Ca | 0.31±0.02Aa | 9.08±0.35Ca | 0.77±0.13Ab | 22.86±3.21Bc | |
| 10—20 | 3.13±0.76Bb | 0.67±0.12Ab | 18.19±1.40Cb | 0.30±0.02Aab | 8.64±0.56Ca | 1.03±0.18Aab | 28.24±1.28Ab | ||
| 2—30 | 1.63±0.21Bb | 0.53±0.11ABb | 11.20±1.04Bc | 0.27±0.01Ab | 8.48±0.46BCa | 1.18±0.06Aa | 32.28±2.21Aa | ||
| 不同大写字母表示同一土层不同林地间差异显著(P<0.05); 不同小写字母表示同一林地不同土层间差异显著(P<0.05); HN: 水解氮Hydrolyzed nitrogen; TP:全磷Total phosphorus;AP:速效磷Available phosphorus;TK:全钾Total potassium;AK:速效钾Available potassium | |||||||||
4种林地的MBC含量与MBN含量均随土层增加而减少, 不同季节不同林地的MBC和MBN含量在不同土层存在差异。4种林地中, 裸地的MBC和MBN含量随土层减小的幅度最小, 各土层之间差异不明显。冬季锥栗林上、下土层的MBC含量差异最大, 0—10 cm的MBC含量是20—30 cm的1.95倍, 其他季节的差异为马尾松林最大。茶园上、下土层的MBN含量差异在4种林地中较大, 其中夏季0—10 cm与20—30 cm的差异为22.81 mg/kg(表 4)。
| 林地类型 Forest type |
土层 Soil layer/cm |
春季 Spring |
夏季 Summer |
秋季 Autumn |
冬季 Winte |
|
| MBC/(mg/kg) | 茶园 | 0—10 | 370.20±48.62Ba | 481.15±35.88Ba | 468.81±112.15Aa | 265.58±100.87ABa |
| 10—20 | 346.74±64.89Aa | 410.25±45.13Aab | 334.72±58.95Bab | 246.47±98.28Aa | ||
| 20—30 | 174.49±41.04BCb | 348.90±63.41Ab | 262.99±39.05ABb | 185.33±64.91ABa | ||
| 马尾松 | 0—10 | 586.50±74.84Aa | 609.56±61.30Aa | 537.10±90.01Aa | 376.40±82.04Aa | |
| Pinus massoniana forest | 10—20 | 463.82±36.12Aa | 481.26±51.08Aab | 520.30±85.92Aa | 288.51±63.07Aa | |
| 20—30 | 377.57±40.55Aa | 400.66±35.82Ab | 349.89±39.52Ab | 246.48±37.91Aa | ||
| 锥栗 | 0—10 | 399.55±36.27Ba | 450.45±84.11Ba | 416.57±87.17Aa | 431.78±108.18Aa | |
| Castanea henryi forest | 10—20 | 311.20±46.05Aab | 412.17±86.82Aa | 301.19±57.75Ba | 240.74±44.64Ab | |
| 20—30 | 252.73±42.61ABb | 247.31±21.54Bb | 224.93±96.80Ba | 221.65±35.44Ab | ||
| 裸地 | 0—10 | 47.01±17.31Ca | 61.35±19.56Ca | 47.53±0.83Ba | 45.35±11.77Ba | |
| 10—20 | 41.12±11.40Ba | 44.08±11.81Bab | 37.75±5.99Cab | 29.67±3.50Ba | ||
| 20—30 | 37.21±13.55Ca | 24.84±2.78Cb | 23.46±9.58Cb | 28.14±5.83Ba | ||
| MBN/(mg/kg) | 茶园 | 0—10 | 31.80±7.83Aa | 98.86±13.02Aa | 51.71±5.30Aa | 31.25±7.44ABa |
| 10—20 | 20.05±7.63Aa | 85.04±13.73Aab | 42.45±6.68Aab | 30.42±1.96Aa | ||
| 20—30 | 17.41±7.63Ab | 76.05±13.13Ab | 35.16±2.24Ab | 27.38±5.54Aa | ||
| 马尾松 | 0—10 | 23.51±5.18Aa | 82.96±27.90Aa | 46.46±3.01Aa | 36.22±2.18Aa | |
| 10—20 | 17.28±6.84Aa | 70.56±16.34Aab | 41.07±2.12Aa | 25.99±4.80Aa | ||
| 20—30 | 15.21±5.17Aa | 63.61±13.81Ab | 33.32±3.39Ab | 20.74±7.38Aa | ||
| 锥栗 | 0—10 | 22.12±1.31Ba | 49.09±5.58Ba | 42.58±1.87Aa | 37.03±5.69Aa | |
| 10—20 | 19.08±1.78Aab | 45.31±4.89Aa | 39.27±2.15Ba | 29.63±5.04Ab | ||
| 20—30 | 12.73±1.67BCb | 29.04±6.12Bb | 33.46±4.23Ba | 24.33±4.88Ab | ||
| 裸地 | 0—10 | 12.03±3.43Ca | 30.43±3.91Ca | 21.15±7.66Ba | 14.52±1.69Ba | |
| 10—20 | 11.06±1.95Ba | 29.03±6.12Bab | 17.97±7.83Cab | 11.06±1.96Ba | ||
| 20—30 | 6.36±1.71Ca | 22.08±5.94Cb | 9.54±5.45Cb | 9.82±2.04Ba | ||
| 不同大写字母表示同一土层不同林地间差异显著(P<0.05); 不同小写字母表示同一林地不同土层间差异显著(P<0.05); MBC: 微生物生物量碳Microbial biomass carbon; MBN: 微生物生物量氮Microbial biomass Nitrogen | ||||||
各林地的土壤养分和微生物碳、氮含量呈现出不同的季节变化规律。茶园四季的TC含量无显著差异, 均在7 g/kg左右, HN含量冬季最大(57.53 mg/kg), 是秋季的近2倍。马尾松林四季的TC含量相差不大, 均在6 g/kg左右, HN含量冬季(18.66 mg/kg)显著低于其他季节, AP含量均在10 mg/kg左右, 无明显季节差异。锥栗林四季的TC含量春季(11.94 g/kg)显著高于其他季节, TN含量春(2.65 g/kg)>秋(1.69 g/kg)>夏(1.21 g/kg)>冬(1.00 g/kg), AP含量夏季最大(19.82 mg/kg), MBC含量差异不显著。裸地四季的TC含量春季最大, TN含量均在0.8 g/kg左右, AP含量(10 mg/kg左右)无显著差异。4种林地的MBN含量均为夏>秋>冬>春。
同一季节不同林地的土壤养分和微生物碳、氮含量存在差异。TC含量呈现出锥栗林>茶园>马尾松林>裸地的规律, TN、HN、TP、AP含量均表现为茶园、锥栗林>马尾松林>裸地。春、夏季茶园的AP含量远高于其他林地, 茶园春季43.18 mg/kg, 夏季36.33 mg/kg, 其他林地的AP含量则在10 mg/kg左右。裸地的TK含量显著高于其他林地, 马尾松林的AK含量在4种林地中最小。MBC含量表现出马尾松林>茶园、锥栗林>裸地的规律, 其中裸地四季的MBC含量维持在40 mg/kg左右, 远远低于其他林地。MBN含量茶园>马尾松>锥栗>裸地, 夏季4种林地的MBN含量差异最大, 最大的茶园(86.65 mg/kg)与最小的裸地(27.18 mg/kg)相差59.47 mg/kg(图 7)。

|
| 图 7 不同林地土壤养分和微生物碳、氮含量的季节变化 Fig. 7 Seasonal variation of soil nutrient, soil microbial biomass carbon and nitrogen content in different forest land 不同大写字母表示同一土层不同林地间差异显著(P<0.05); 不同小写字母表示同一林地不同土层间差异显著(P<0.05) |
分别对4种林地的土壤呼吸和水热因子(近地面气温、土壤温度、土壤湿度)进行单因子回归分析, 从指数模型、对数模型、线性模型、多项式模型和幂函数模型五种模型中筛选出拟合度最高的模型(图 8)。从图中可以明显看出, 茶园土壤呼吸受土壤温度和土壤湿度影响较大(R2分别为0.77和0.78), 而裸地的土壤呼吸受土壤湿度(R2=0.27)影响大于温度。各模型对茶园的拟合度最高, R2达到0.7以上, 而裸地的R2不超过0.3。近地面气温和土壤温度与土壤呼吸的关系模型中, 二次多项式模型对茶园、锥栗林和裸地的拟合度较高, 而马尾松林更适合指数模型(近地面气温)和幂函数模型(土壤温度)。土壤湿度与土壤呼吸的关系模型中, 4种林地均适用于多项式模型。

|
| 图 8 不同林地土壤呼吸速率与近地面气温、土壤温度和土壤湿度的关系 Fig. 8 Relationship between soil respiration rate and near surface air temperature, soil temperature and soil moisture in different forest lands |
4种模型中, 土壤温度、湿度共同建立的幂-指数函数模型对茶园的土壤呼吸拟合度较高, R2达到0.8, 远高于其他三种林地, 并且也高于温度、湿度的单因子模型。马尾松林的土壤呼吸较适用于土壤温度、湿度建立的对数函数模型。加入近地面气温进行三因子回归分析得到的线性模型对茶园和马尾松林的拟合度低于土壤温度、湿度双因子模型, 而三因子线性模型(进入回归法)对锥栗林和裸地的土壤呼吸的拟合度最优, R2分别为0.565和0.281(表 5)。
| 林地Forest land | 模型Model | 回归方程Regression equation | R2 | P |
| 茶园Tea garden | 线性函数(逐步回归) | Rs=-0.015+0.06W+0.078T2 | 0.788 | <0.05 |
| 线性函数(进入回归) | Rs=0.011+0.052T2+0.056W+0.028T1 | 0.795 | <0.05 | |
| 对数函数 | ln(Rs)=0.244+0.012T2ln(W) | 0.774 | <0.05 | |
| 幂-指数函数 | Rs=0.416e0.026T2W0.452 | 0.800 | <0.05 | |
| 马尾松林 | 线性函数(逐步回归) | Rs=0.279+0.13T2 | 0.448 | <0.05 |
| Pinus massoniana forest | 线性函数(进入回归) | Rs=0.05+0.146T2-0.017W+0.014T1 | 0.451 | <0.05 |
| 对数函数 | ln(Rs)=0.398+0.009T2ln(W) | 0.471 | <0.05 | |
| 幂-指数函数 | Rs=1.099e0.035T2W0.081 | 0.431 | <0.05 | |
| 锥栗林 | 线性函数(逐步回归) | Rs=0.218+0.101T1 | 0.552 | <0.05 |
| Castanea henryi forest | 线性函数(进入回归) | Rs=-0.068+0.064T2-0.008W+0.063T1 | 0.565 | <0.05 |
| 对数函数 | ln(Rs)=0.155+0.01T2ln(W) | 0.497 | <0.05 | |
| 幂-指数函数 | Rs=1.187e0.069T2W-0.24 | 0.559 | <0.05 | |
| 裸地Bare land | 线性函数(逐步回归) | Rs=1.231+0.066T1-0.06T2 | 0.266 | <0.05 |
| 线性函数(进入回归) | Rs=1.227-0.059T2+0.00052W+0.066T1 | 0.284 | <0.05 | |
| 对数函数 | ln(Rs)=0.152+0.003T2ln(W) | 0.111 | <0.05 | |
| 幂-指数函数 | Rs=1.016e0.11T2W0.31 | 0.145 | <0.05 | |
| Rs: 土壤呼吸Soil respiration; T1: 近地面气温Near surface air temperature; T2: 土壤温度Soil temperature; W: 土壤湿度Soil moisture | ||||
茶园的土壤呼吸与TP、AP、TK、AK含量存在极显著(P<0.01)正相关, 马尾松林的土壤呼吸受TP、TK、AK含量的影响显著(P<0.05), 锥栗林的土壤呼吸与TK、AK、MBN含量呈现正相关, 裸地的土壤呼吸受MBN含量影响较为显著(表 6)。4种林地土壤呼吸速率与土壤养分含量的线性模型拟合程度均较高, 除去水热因子外, 茶园土壤呼吸主要受P元素影响, 马尾松林地土壤呼吸受K元素影响较多(表 7)。
| 林地 Forest land |
茶园 Tea garden |
马尾松 Pinus massoniana forest |
锥栗 Castanea henryi forest |
裸地 Bare land |
| 全碳TC/(g/kg) | -0.100 | -0.096 | -0.241 | -0.365* |
| 全氮TN/(g/kg) | -0.297 | -0.081 | -0.232 | 0.098 |
| 水解氮HN/(mg/kg) | -0.096 | 0.213 | -0.053 | -0.163 |
| 全磷TP/(g/kg) | 0.542** | 0.491** | 0.227 | 0.014 |
| 有效磷AP/(mg/kg) | 0.530** | -0.303 | 0.217 | 0.164 |
| 全钾TK/(g/kg) | 0.719** | 0.648** | 0.374* | 0.120 |
| 速效钾AK/(mg/kg) | 0.736** | 0.502** | 0.362* | 0.346* |
| 微生物生物量碳MBC/(mg/kg) | 0.503 | 0.576* | 0.514 | 0.169 |
| 微生物生物量氮MBN/(mg/kg) | 0.614* | 0.329 | 0.595* | 0.753** |
| *P<0.05; **P<0.01 | ||||
| 林地Forest land | 回归方程Regression equation | R2 | P |
| 茶园Tea garden | Rs=2.887-17.981 X(TP)+0.122X(AP)+0.161X(AK) | 0.998 | <0.01 |
| 马尾松Pinus massoniana forest | Rs=0.606+6.984 X(TP)+3.759 X(TK)-0.172 X(AK) | 0.999 | <0.01 |
| 锥栗Castanea henryi forest | Rs=-0.355+1.222 X(TK)-0.014 X(AK)+0.053 X(MBN) | 0.999 | <0.01 |
| 裸地Bare land | Rs=-1.645+1.183 X(TC)-0.123 X(AK)+0.155 X(MBN) | 0.987 | <0.01 |
土壤呼吸是森林生态系统碳循环的重要一环, 易受到各种生物和环境因素影响。本文实验结果表明, 武夷山4种林地土壤呼吸速率的日变化趋势与近地面气温和土壤温度相同;在季节动态上, 土壤呼吸速率与近地面气温、土壤温度和土壤湿度一样, 表现出明显的季节变化(夏>春>秋>冬)。土壤呼吸的动态变化结果与傅勇等[26]、孙海燕[27]等的研究结果一致, 温度和湿度对土壤呼吸的影响很大, 有研究认为土壤温度和湿度是土壤呼吸速率季节变化的主导因子[25, 28-29]。在对温度(近地面气温和土壤温度)与土壤呼吸之间的关系模拟中, 本文尝试使用了线性函数、指数函数、幂函数、对数函数和多项式, 其中一元二次多项式模型对茶园、锥栗林和裸地的土壤呼吸拟合较好。许多学者在研究温度与土壤呼吸的关系时选用指数函数[15, 25-26, 30], 但本文指数函数仅对马尾松林近地面气温与土壤呼吸的关系拟合较好。目前研究中使用较为广泛的土壤湿度与土壤呼吸的单因子模型为线性模型[15]和一元二次模型[2, 15, 26], 本文4种林地的土壤呼吸与土壤湿度的关系用一元二次模型拟合度较高。对茶园、锥栗林和裸地三种林地来说, 水热因子的复合模型比单因子模型的拟合度更高, 说明温度和湿度对土壤呼吸的交互作用超过两者的单独作用, 更能够解释土壤呼吸的变异。无论水热单因子模型还是复合因子模型, 对茶园土壤呼吸变化的拟合程度(74.5%—80%)远高于其他林地, 说明茶园土壤呼吸对温度、湿度的响应较为敏感[31]。通常, 在多元回归过程中, 因子数量的增加同时导致R2增大[32]。马尾松林土壤温度单因子对土壤呼吸的拟合度为0.51, 加入近地面温度和土壤湿度后的模型拟合度反而降低, 说明马尾松林土壤呼吸主要受土壤温度调节, 这与巫志龙等[31]对杉阔人工林的研究结果相似。裸地土壤呼吸与水热因子的关系模型R2值仅为0.2左右, 可能是因为裸地地表无植被, 缺乏植物根系, 且土壤有机质含量低, 降低了土壤呼吸速率对温度、湿度的响应。土壤呼吸速率与温度之间呈现一元二次函数关系, 在林地四季的正常温度范围内为正相关。这是因为温度升高能够促进植物根系生长和微生物活动, 提高土壤有机质的分解速率, 因此土壤呼吸速率也随之提高[33-34]。土壤湿度与土壤呼吸的关系(除裸地外)则表现为湿度较低时呈现正相关, 超过一定阈值时呈现负相关。当土壤水分过低成为限制因子时, 增加湿度能够促进植物生长和微生物呼吸;当水分含量过高则会导致土壤通透性变差, 土壤空隙中的氧气含量减少, 因此对微生物活动产生抑制, 土壤呼吸速率随之降低[35]。
在本文研究的4种林地中, 茶园的土壤呼吸在4种林地中最高(季节均值为3.10 μmol m-2 s-1), 范利超[36]的研究结果也表明茶园的土壤呼吸高于锥栗、马尾松等林地。土壤呼吸包括根系呼吸和土壤动物、微生物参与的有机物氧化等基础呼吸[15], 其影响因素众多且交互性强。植被类型对土壤呼吸的影响较为复杂[15], 树种组成、群落结构的不同使得林内水热环境产生差异, 加上林地生产方式的差异性, 不同林地的土壤微生物、土壤碳库和植物碳分配受到影响, 进而使土壤呼吸产生差异[3]。锥栗林郁闭度较高, 树木较高, 林下植被较少, 林内整体形成较为阴凉通风的环境, 温度和土壤湿度与郁闭度低、受阳光照射较多的茶园和马尾松林相比较低, 因此其土壤呼吸速率也在三种林地中最低。由于茶树低矮, 地表其他灌草植物少, 每垄植株之间裸露的土壤也较多, 茶园的近地面气温和土壤温度高于马尾松林和锥栗林, 这可能是茶园土壤呼吸高的原因。加上茶园通常会进行定期翻耕、除草等活动, 土壤较为疏松, 有利于植物根系生长及土壤上层的微生物活动。锥栗作为阔叶落叶树种, 每年产生的凋落物将大量的C、N等元素带到了土壤中, 使得土壤C、N含量高于针叶林[37]。植被类型对土壤微生物群落存在极大影响[38], 土壤MBC、MBN含量会影响土壤的有机质转化和部分氮元素的形成, 而土壤有机碳是土壤微生物呼吸的碳源, 氮元素则促进植物生长, 为土壤呼吸提供更多的底物[39], 很多研究[25, 40-41]表明土壤有机碳和微生物碳含量与土壤呼吸速率相关性显著。但本文研究发现土壤呼吸速率与TC、TN等相关性不显著, 反而与P、K元素以及MBN相关性较强, 因此林地土壤有机质含量对土壤呼吸变异的贡献不大。这与张涛等[42]对马尾松林土壤呼吸的研究结果相似, 土壤呼吸与TC、TN无显著相关关系, 杨明臻等[43]的研究也发现磷元素对土壤呼吸产生影响。南方土壤缺磷比较严重[44], 研究区位于福建省武夷山国家公园, 植物生长需要P、K元素, 当土壤中P、K成为限制因子时, 其含量的细微变化就会影响土壤呼吸速率。与裸地相比, 其他三种林地由于受到人工施肥、凋落物补给等, 土壤C、N含量较高, 土壤呼吸受C、N元素的影响低于P、K元素;而裸地本身较为贫瘠, 缺乏有机质, 其土壤呼吸速率与TC、MBN含量相关性显著。
土壤呼吸作为生态系统碳循环重要的一部分, 对生态系统碳平衡和气候动态方面起着十分重要的作用。与裸地相比, 锥栗、茶园、马尾松等林地的地表植被覆盖率上升、对温度和水分的调控能力提高、土壤呼吸的能力增强, 在一定程度上能够提高土壤质量、改善生态环境, 有利于森林生态系统的平衡和林地的可持续发展。茶园土壤呼吸受水热因子的影响较大, 且与其他林地相比, 土壤呼吸速率有所增加。武夷山作为中国著名的茶叶产区, 茶园面积广大, 若今后大力发展茶产业, 是否会导致茶园的土壤碳排放增加, 还需要进一步研究。土壤呼吸的影响因素众多而复杂, 对于影响武夷山地区土壤呼吸的众多因子本文未能全面探讨。为了更准确和系统地发现土壤呼吸的变化规律和影响因素, 在今后的研究中还应考虑到凋落物、土壤微生物群落、人类活动等因子对土壤呼吸的影响。并且对于茶园, 尽管武夷山茶园施肥量与其他茶园相比相对较低, 对土壤呼吸影响可能较小, 但施肥量、肥料种类等因素在今后的研究中也是不可忽略的部分。土壤呼吸是一个动态变化过程, 要深入精确地研究土壤呼吸的变化规律, 还需要更长期的定点定位动态监测, 不断加强与完善土壤呼吸的研究系统。
| [1] |
Raich J W, Potter C S. Global patterns of carbon dioxide emissions from soils. Global Biogeochemical Cycles, 1995, 9(1): 23-36. DOI:10.1029/94GB02723 |
| [2] |
Luo Y Q, Wan S Q, Hui D F, Wallace L L. Acclimatization of soil respiration to warming in a tall grass prairie. Nature, 2001, 413(6856): 622-625. DOI:10.1038/35098065 |
| [3] |
史宝库, 金光泽, 汪兆洋. 小兴安岭5种林型土壤呼吸时空变异. 生态学报, 2012, 32(17): 5416-5428. |
| [4] |
Davidson E A, Janssens I A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature, 2006, 440(7081): 165-173. DOI:10.1038/nature04514 |
| [5] |
Zhang Q, Lei H M, Yang D W. Seasonal variations in soil respiration, heterotrophic respiration and autotrophic respiration of a wheat and maize rotation cropland in the North China Plain. Agricultural and Forest Meteorology, 2013, 180: 34-43. DOI:10.1016/j.agrformet.2013.04.028 |
| [6] |
Zhang Y J, Guo S L, Zhao M, Du L L, Li R J, Jiang J S, Wang R, Li N N. Soil moisture influence on the interannual variation in temperature sensitivity of soil organic carbon mineralization in the Loess Plateau. Biogeosciences, 2015, 12(11): 3655-3664. DOI:10.5194/bg-12-3655-2015 |
| [7] |
Karhu K, Fritze H, Hämäläinen K, Vanhala P, Jungner H, Oinonen M, Sonninen E, Tuomi M, Spetz P, Kitunen V, Liski J. Temperature sensitivity of soil carbon fractions in boreal forest soil. Ecology, 2010, 91(2): 370-376. DOI:10.1890/09-0478.1 |
| [8] |
Vanhala P, Karhu K, Tuomi M, Björklöf K, Fritze H, Hyvärinen H, Liski J. Transplantation of organic surface horizons of boreal soils into warmer regions alters microbiology but not the temperature sensitivity of decomposition. Global Change Biology, 2011, 17(1): 538-550. DOI:10.1111/j.1365-2486.2009.02154.x |
| [9] |
Fu S L, Cheng W X. Rhizosphere priming effects on the decomposition of soil organic matter in C4 and C3 grassland soils. Plant and Soil, 2002, 238(2): 289-294. DOI:10.1023/A:1014488128054 |
| [10] |
Chang S X, Shi Z, Thomas B R. Soil respiration and its temperature sensitivity in agricultural and afforested poplar plantation systems in northern Alberta. Biology and Fertility of Soils, 2016, 52(5): 629-641. DOI:10.1007/s00374-016-1104-x |
| [11] |
Yang S H, Liu X J, Liu X Y, Xu J Z. Effect of water management on soil respiration and NEE of paddy fields in Southeast China. Paddy and Water Environment, 2017, 15(4): 787-796. DOI:10.1007/s10333-017-0591-1 |
| [12] |
段北星, 蔡体久, 宋浩, 肖瑞晗. 寒温带兴安落叶松林凋落物层对土壤呼吸的影响. 生态学报, 2020, 40(4): 1357-1366. |
| [13] |
李岩, 干珠扎布, 胡国铮, 万运帆, 李玉娥, 旦久罗布, 何世丞, 谢文栋, 高清竹. 增温对青藏高原高寒草甸呼吸作用的影响. 生态学报, 2020, 40(1): 266-273. |
| [14] |
张俞, 熊康宁, 喻阳华, 许敏, 程雯, 谭代军. 中国南方喀斯特石漠化地区3种经济林土壤呼吸日动态特征. 中南林业科技大学学报, 2019, 39(1): 92-99. |
| [15] |
郑鹏飞, 余新晓, 贾国栋, 李红娟, 王渝淞, 朱栩辉. 北京山区不同植被类型的土壤呼吸特征及其温度敏感性. 应用生态学报, 2019, 30(5): 1726-1734. |
| [16] |
Fender A C, Gansert D, Jungkunst H F, Fiedler S, Beyer F, Schützenmeister K, Thiele B, Valtanen K, Polle A, Leuschner C. Root-induced tree species effects on the source/sink strength for greenhouse gases (CH4, N2O and CO2) of a temperate deciduous forest soil. Soil Biology and Biochemistry, 2013, 57: 587-597. DOI:10.1016/j.soilbio.2012.08.004 |
| [17] |
陈晓璐, 潘文斌. 武夷新区土地利用变化及景观格局研究. 环境科学与管理, 2013, 38(12): 63-68. DOI:10.3969/j.issn.1673-1212.2013.12.015 |
| [18] |
牛莉平, 许嘉巍, 靳英华, 尹海霞, 王嫒林, 刘羽霞, 张英洁. 温带森林不同演替阶段下的土壤CO2排放通量昼间变化. 生态学报, 2017, 37(8): 2673-2683. |
| [19] |
任寅榜. 沿武夷山海拔梯度土壤置换对异养呼吸的影响[D]. 福州: 福建师范大学, 2018.
|
| [20] |
施政, 汪家社, 何容, 王国兵, 方燕鸿, 徐自坤, 张增信, 阮宏华. 武夷山不同海拔植被土壤呼吸季节变化及对温度的敏感性. 应用生态学报, 2008, 19(11): 2357-2363. |
| [21] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 1999.
|
| [22] |
林先贵. 土壤微生物研究原理与方法. 北京: 高等教育出版社, 2010.
|
| [23] |
吴金水. 土壤微生物生物量测定方法及其应用. 北京: 气象出版社, 2006.
|
| [24] |
张文丽. 不同耕作下土壤呼吸及其影响因子研究[D]. 长春: 中国科学院大学(中国科学院东北地理与农业生态研究所), 2019.
|
| [25] |
刘宝, 王民煌, 余再鹏, 林思祖, 林开敏. 中亚热带天然林改造成人工林后土壤呼吸的变化特征. 林业科学, 2019, 55(4): 1-12. |
| [26] |
傅勇, 王淘, 杨志平, 周伟, 刘琦, 任万军, 陈勇. 川西平原灌区不同水旱轮作模式周年土壤呼吸特征. 生态学报, 2019, 39(18): 6701-6709. |
| [27] |
孙海燕, 赵俊平, 肖艳玲, 冯建伟, 张占贵. 模拟氮沉降对武夷山亚热带常绿阔叶林土壤呼吸的影响. 生态环境学报, 2018, 27(9): 1632-1638. |
| [28] |
王楠, 潘小承, 白尚斌. 模拟酸雨对我国亚热带毛竹林土壤呼吸及微生物多样性的影响. 生态学报, 2020, 40(10): 3420-3430. |
| [29] |
温超, 单玉梅, 晔薷罕, 张璞进, 木兰, 常虹, 任婷婷, 陈世苹, 白永飞, 黄建辉, 孙海莲. 氮和水分添加对内蒙古荒漠草原放牧生态系统土壤呼吸的影响. 植物生态学报, 2020, 44(1): 80-92. |
| [30] |
Chen S T, Huang Y, Zou J W, Shen Q R, Hu A H, Qin Y M, Chen H S, Pan G X. Modeling interannual variability of global soil respiration from climate and soil properties. Agricultural and Forest Meteorology, 2010, 150(4): 590-605. DOI:10.1016/j.agrformet.2010.02.004 |
| [31] |
巫志龙, 周成军, 周新年, 刘富万, 朱奇雄, 黄金湧, 陈文. 不同强度采伐5年后杉阔混交人工林土壤呼吸速率差异. 林业科学, 2019, 55(6): 142-149. |
| [32] |
张录, 张芳, 熊黑钢, 段鹏程, 李荣荣, 张南. 不同季节强碱土土壤呼吸影响因子分析与模型预测. 干旱地区农业研究, 2017, 35(1): 71-78. |
| [33] |
吴建召, 严思维, 崔羽, 罗清虎, 林勇明, 王道杰, 吴承祯. 地震灾区2种气候生态治理区恢复初期土壤呼吸动态及其影响因子. 北京林业大学学报, 2019, 41(3): 93-104. |
| [34] |
陈毅, 闫文德, 郑威, 廖菊阳, 盘昱良, 梁小翠, 杨坤. 模拟氮沉降凋落物管理对樟树人工林土壤呼吸的影响. 生态学报, 2018, 38(21): 7830-7839. |
| [35] |
付若仙, 余景松, 张云彬, 俞元春, 陶晓. 氮添加下城市森林土壤呼吸动态变化及其影响因素. 应用生态学报, 2020, 31(3): 744-752. |
| [36] |
范利超. 茶园土壤呼吸及其组分研究[D]. 北京: 中国农业科学院, 2015.
|
| [37] |
万晓华, 黄志群, 何宗明, 胡振宏, 余再鹏, 王民煌, 杨玉盛, 范少辉. 杉木采伐迹地造林树种转变对土壤可溶性有机质的影响. 应用生态学报, 2014, 25(1): 12-18. |
| [38] |
Monson R K, Lipson D L, Burns S P, Turnipseed A A, Delany A C, Williams M W, Schmidt S K. Winter forest soil respiration controlled by climate and microbial community composition. Nature, 2006, 439(7077): 711-714. DOI:10.1038/nature04555 |
| [39] |
Luo Y Q, Zhou X H. Soil respiration and the environment[M]. Academic Press, 2006.
|
| [40] |
Song X Z, Kimberley M O, Zhou G M, Wang H L. Soil carbon dynamics in successional and plantation forests in subtropical China. Journal of Soils and Sediments, 2017, 17(9): 2250-2256. DOI:10.1007/s11368-016-1421-6 |
| [41] |
王泽西, 陈倩妹, 黄尤优, 邓慧妮, 谌贤, 唐实玉, 张健, 刘洋. 川西亚高山森林土壤呼吸和微生物生物量碳氮对施氮的响应. 生态学报, 2019, 39(19): 7197-7207. |
| [42] |
张涛, 罗雷, 雷杰, 宋秀丽, 王鹏程. 三峡库区马尾松林土壤呼吸的空间变异特征. 华中农业大学学报, 2017, 36(1): 28-34. |
| [43] |
杨明臻, 林衣东, 韩文炎. 施氮对茶园土壤基础呼吸的影响. 土壤通报, 2012, 43(6): 1355-1360. |
| [44] |
兰忠明, 林新坚, 张伟光, 张辉, 吴一群. 缺磷对紫云英根系分泌物产生及难溶性磷活化的影响. 中国农业科学, 2012, 45(8): 1521-1531. DOI:10.3864/j.issn.0578-1752.2012.08.008 |