 2021, Vol. 41
2021, Vol. 41文章信息
- 刘领, 马宜林, 悦飞雪, 乔鑫鑫, 尹飞, 王艳芳
- LIU Ling, MA Yilin, YUE Feixue, QIAO Xinxin, YIN Fei, WANG Yanfang
- 生物炭对褐土旱地玉米季氮转化功能基因、丛枝菌根真菌及N2O释放的影响
- Effects of biochar on nitrogen transformation functional genes abundances, arbuscular mycorrhizal fungi and N2O emission of rainfed maize season in cinnamon soil
- 生态学报. 2021, 41(7): 2803-2815
- Acta Ecologica Sinica. 2021, 41(7): 2803-2815
- http://dx.doi.org/10.5846/stxb202006041438
-
文章历史
- 收稿日期: 2020-06-04
- 网络出版日期: 2021-01-27

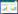




氧化亚氮(N2O)是除甲烷(CH4)和二氧化碳(CO2)之外的一种重要温室气体, 其增温潜势比CO2高约300倍, N2O可以破坏臭氧层, 造成气候变暖, 威胁人类健康[1]。农田土壤由于大量施入氮肥, 成为N2O的主要排放源, 大约占总排放量的80%[2]。N2O的产生主要来自于土壤的硝化作用和反硝化作用。在硝化过程中, N2O是羟胺氧化成N2O-N过程中产生的, 主要由含有氨单加氧酶(amoA)基因的氨氧化古菌(Ammonia-oxidizing archaea, AOA)和氨氧化细菌(Ammonia-oxidizing bacteria, AOB)参与完成, 并对土壤硝化起着限速作用[3-4]。在反硝化过程中, N2O则是由NO3-、NO2逐步还原成NO、N2O、N2过程中产生的。土壤N2O的产生主要受反硝化微生物活性的影响, 其中亚硝酸还原酶和氧化亚氮还原酶是土壤反硝化作用的关键酶和限速酶, 编码亚硝酸还原酶的nirS、nirK基因和编码氧化亚氮还原酶的nosZ基因代表反硝化微生物活性。在nirS、nirK 基因编码的亚硝酸还原酶的作用下NO2-还原为NO, 并生成N2O, 同时在由nosZ基因编码的氧化亚氮还原酶的作用下N2O还原为N2。
丛枝菌根(arbuscular mycorrhizal, AM)真菌是一类分布广泛(能与80%以上植物根系形成共生体)且数量丰富(占土壤总微生物生物量的30%以上)的土壤有益微生物, AM真菌对土壤N素循环起着重要作用, 可以显著影响N素的吸收与矿化、生物固N、硝化和反硝化, 以及N素的淋失等诸多土壤氮素循环过程[5]。已有研究表明, AM真菌影响农田N2O排放的潜在机理主要为:AM真菌可以通过改变作物-土壤系统的水分关系、影响土壤结构和土壤通气性能来调控N2O排放;AM真菌能够通过竞争吸收氮素来调控N2O排放;AM真菌可以通过改变根际沉积过程来调控N2O排放, 接种AM真菌能够改变植物根系分泌物的组成及数量, 从而导致根际及菌根际微生物群落发生变化, 包括硝化、反硝化细菌群落来调控N2O排放[6-8]。
生物炭是一些农作物秸秆、薪柴、杂草、粪便等生物质在缺氧或低氧条件下通过高温裂解后的富碳固体产物。近年来, 国内外学者对生物炭在改良土壤、提高作物产量、碳固存、温室气体减排等方面进行了研究[9-12], 特别是农田施用生物炭对N2O排放的影响机制得到了广泛的关注。一些研究表明, 农田施入生物炭可以通过影响硝化、反硝化功能基因来影响土壤N2O排放[7, 13]。另外一些研究表明, 生物炭对AM真菌具有特别的促进作用, 生物炭影响土壤AM真菌的机制主要表现在:生物炭改善土壤理化特性有利于AM真菌孢子的萌发和生长;生物炭调节植物-真菌信号物质影响AM真菌的萌发和菌丝的分枝;生物炭可能通过影响土壤中的解磷细菌活性, 从而间接影响AM真菌[14-16]。
基于N2O排放是一系列复杂土壤生态过程的综合反映, 并受到多种生物和非生物因素的交互影响。生物炭输入是如何调节土壤硝化、反硝化功能基因和AM真菌的, 以及他们之间的关系需要进一步了解。目前多数研究主要针对生物炭对AM真菌的影响[15, 17]或生物炭对N转化相关功能基因及N2O释放的影响[3, 13], 然而对施入生物炭对土壤AM真菌及N转化相关功能基因的影响以及它们之间的关系研究较少。
河南省西部(豫西)位于黄土丘陵区, 属暖温带大陆性季风气候, 土壤类型为褐土, 旱地农业在该地区占据重要地位, 由于农田化肥使用量大, 成为N2O的重要排放源[9]。本研究以豫西地区旱地玉米农田为研究对象, 探讨生物炭施入后对土壤理化性质、N转化相关功能基因丰度、AM真菌及N2O释放的影响, 并探讨它们之间的相关性, 以期为旱地农田合理施用生物炭实现N2O减排提供理论依据。
1 材料与方法 1.1 试验地概况试验在河南省洛阳市河南科技大学开元校区农场(34°41′N, 112°27′E)进行。该地区年均气温13.7 ℃, 年均无霜期216 d, 年降水量约600—800 mm, 主要分布在6—8月, 作物种植方式为小麦-玉米轮作, 土壤类型为褐土, 土壤基本理化性质为:pH值为7.4, 有机质含量为15.1 g/kg, 总氮1.0 g/kg, 碱解氮78.6 mg/kg, 速效磷9.2 mg/kg, 速效钾116.5 mg/kg, 粘粒、粉粒、砂粒比例分别为20.34%、30.87%、48.79%。
1.2 试验材料供试的玉米品种为“郑单958”, 供试生物炭材料为小麦秸秆生物炭(河南商丘三利新能源有限公司, 热裂解炭化温度350—450℃), 生物炭基本理化性质为:pH值10.4, 比表面积为8.92 m2/g, 有机碳为52.2%, 全氮为5.9 g/kg, 全磷为0.89 g/kg, 全钾为23.2 g/kg。
1.3 试验设计试验于2019年6月5日开始, 9月10日结束。试验期内总降雨量为428.3 mm, 与常年相比, 属于平水年。生物炭用量共设3个处理:T0:对照, 不施用生物炭;T1:施用生物炭20 t/hm2;T2:施用生物炭40 t/hm2;每处理重复3次, 共9个小区, 小区面积32 m2(4 m×8 m)。2019年6月5日种植玉米, 行距为60 cm, 株距为25 cm, 种植密度为67000株/hm2, 各处理施氮量225 kg N/hm2, 以含氮量46%的尿素为氮源, 分2次施用, 其中基肥占60%, 大喇叭口期追肥占40%。各处理施磷肥40 kg P/hm2, 钾肥80 kg K/hm2, 分别以含P2O5 12%的过磷酸钙和含K2O 45%的硫酸钾作为肥源。生物炭、基施氮肥、磷肥和钾肥均在播前撒于地表后微耕机翻耕, 与10 cm左右土层混合均匀, 大喇叭口期在玉米垄一侧距离植株5—6 cm处采用点种器追施氮肥, 穴深6—8 cm, 施入氮肥后立即覆土。试验期间采用雨养方式, 不进行灌溉, 定期除草。
1.4 测定项目与方法 1.4.1 N2O排放通量测定本试验于追肥后第2天开始进行气体样品采集, 每2天采样1次, 连续采样4次恢复为5—7 d采样1次, 强降雨后第2天开始每2天采样1次, 连续采样3次恢复为5—7 d采样1次, 采气直至成熟期。采用密闭式静态箱法测定N2O排放通量, 箱体由有机玻璃制作而成, 包括底座和顶箱两部分。顶箱长、宽、高均为50 cm, 箱体外侧包有一层2.5 cm厚的聚苯乙烯泡沫板, 以减小采样时因太阳辐射所引起的箱内温度变化, 箱内安装风扇以将箱内气体混匀。底座长、宽、高分别为50 cm、50 cm、15 cm, 底座上缘有1.5 cm深的凹槽, 待玉米发芽后将底座随机安置在不同处理的小区, 插入土层5 cm深处, 整个生长季不再移动, 不同处理小区安置5个底座, 底座内种植两棵玉米, 玉米长至九叶期折断, 只留45 cm高度[17]。
采集气体时将顶箱套在底座凹型槽内, 将水注入凹槽加以密封。采样时间为08:00—10:00, 分别于罩箱后0、10、20、30 min用40 mL注射器采集箱内气体40 mL, 将采集的气体注射入40 mL的已抽为真空的集气瓶中, 带回试验室检测。采用岛津气相色谱仪(GC-2010)测定N2O气体浓度, 电子捕获检测器(ECD)检测N2O含量。
N2O排放通量计算公式为:
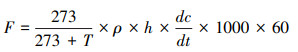
|
(1) |
式中, F为N2O排放通量(μg m-2 h-1);T为采样箱内的平均温度(℃);ρ为标准状态下N2O气体密度, 值为1.977 g/L;h为采气箱高度(m);dc/dt为采气箱内N2O浓度变化率(mL L-1 min-1)。
N2O累积排放量的计算公式如下
|
(2) |
式中, M为土壤N2O累积排放量(μg/m2);F为N2O排放通量(μg m-2 h-1);i为采样次数;ti+1-ti为采样间隔天数。
1.4.2 土壤DNA提取、土壤pH、含水量、容重、有机碳、土壤全氮、硝态氮和铵态氮测定玉米追肥后10 d, 处于玉米抽雄、开花期, 是由营养生长转向生殖生长的关键时期, 肥水需求量大, 生理比较活跃, 我们取该时期土壤测定土壤理化性质和提取土壤DNA。采用环刀法取0—20 cm土层测定土壤容重, 用抖根法[18]采集玉米根际土壤, 一个小区取3个土壤样品, 一部分土壤样品储存在-80℃冰箱用于土壤DNA提取, 一部分土壤用于测定土壤pH、含水量、土壤有机碳、土壤全氮、硝态氮、铵态氮、易提取球囊霉素相关土壤蛋白(EE-GRSP)和总球囊霉素相关土壤蛋白(T-GRSP)。土壤硝态氮和铵态氮采用连续流动分析仪(SAN++, Skalar, 荷兰)测定;土壤含水量采用烘干法进行测定;土壤pH值采用pH计测定(上海精密科学仪器有限公司, 雷磁pHS-3C), 水土比为5∶1。土壤有机碳(SOC)测定采用外加热、重铬酸钾(K)容量法[19];全氮(TN)采用凯氏定氮法(2300全自动定氮仪, Sweden)测定[20]。每个样品重复测定3次。
采用Fast DNASPIN Kit for Soil(MP Biomedicals, 美国)提取土壤DNA, 操作步骤按照试剂盒操作说明进行。用1%的琼脂糖凝胶电泳检测DNA的完整性, 用紫外分光光度计(NanoDrop2000, Thermo Fisher Scientific, 美国)测定DNA浓度及质量。检测质量合格的DNA样品交由上海美吉生物医药有限公司进行荧光定量PCR分析。
1.4.3 AM真菌侵染率和球囊霉素相关土壤蛋白的测定使用抖根法取好土壤样品后, 仔细收集玉米根系, 用清水清洗干净, 用于AM真菌侵染率的测定。AM真菌侵染率采用Bierman & Linderman(1981)的方法测定[21]。易提取球囊霉素相关土壤蛋白(EE-GRSP)、总球囊霉素相关土壤蛋白(T-GRSP)采用李少朋等[22]的方法测定。
1.4.4 实时荧光定量PCR分析本试验选取土壤中AM真菌、氨氧化古菌(AOA)和氨氧化细菌(AOB)的氨单加氧酶(amoA)基因、亚硝酸盐还原酶(nirK、nirS)基因和氧化亚氮还原酶(nosZ)基因进行定量分析, 在ABI7500型荧光定量PCR仪(Applied Biosystems, 美国)进行荧光绝对定量PCR分析。荧光定量PCR的反应体系为20 μL, 其中包含10 μL的ChamQ SYBR Color qPCR Master Mix (2X)(南京诺唯赞生物科技有限公司), 前后引物(5μmol L-1)各0.8 μL, 1 μL DNA模板, 7.4 μL超纯水。具体PCR引物和扩增条件见表 1, 每个样品重复3次。分别以含有AM真菌基因、氨氧化古菌amoA 基因、氨氧化细菌amoA 基因、亚硝酸盐还原酶(nirK、nirS)基因和一氧化二氮还原酶(nosZ)基因的重组PMD18-T载体作为标准质粒, 然后计算出标准质粒的拷贝数, 10倍梯度稀释构建好的各质粒, 90μL稀释液+10μL质粒, 一般做4—6个点, 通过预实验分别选取AOA标准品的10-2—10-6稀释液、AM、AOB、nirS、nosZ标准品的10-2—10-7稀释液、nirK标准品的10-2—10-5稀释液用于制备标准曲线, 根据标准曲线计算基因丰度。
| 目的基因 Targeted gene |
引物 Primer |
引物序列(5′→3′) Primer sequence |
定量PCR反应程序 Reaction conditions of quantitative amplification PCR |
扩增片段长度 Amplified fragment length/(bp) |
扩增效率 Amplified efficiency/% |
参考文献 References |
| AM真菌 | AMV4-5NF AMDGR |
AAGCTCGTAGTTGAATTTCG CCCAACTATCCCTATTAATCAT |
95 ℃预变性5 min, 95 ℃变性30 s, 58 ℃退火30 s, 72 ℃延伸1 min, 35个循环 | 258 | 95.81 | [23] |
| AOA-amoA | Arch-amoAF Arch-amoAR |
STAATGGTCTGGCTTAGACG GCGGCCATCCATCTGTATGT |
95℃预变性5 min, 95℃变性30 s, 58℃退火30 s, 72℃延伸1 min, 35个循环 | 636 | 102.91 | [24] |
| AOB-amoA | AmoA-1F AmoA-2R |
GGGGTTTCTACTGGTGGT CCCCTCKGSAAAGCCTTCTTC |
95℃预变性5min, 95℃变性30 s, 60℃退火30 s, 72℃延伸1 min, 35个循环 | 492 | 100.38 | [25] |
| nirK | nirK1F R3Cu |
GGMATGGTKCCSTGGCA GCCTCGATCAGRTTRTGGTT |
95℃预变性5min, 95℃变性30 s, 55℃退火30 s, 72℃延伸1 min, 35个循环 | 503 | 92.86 | [26] |
| nirS | cd3AF R3cd |
GTSAACGTSAAGGARACSGG GASTTCGGRTGSGTCTTGA |
95℃预变性5min, 95℃变性30 s, 58℃退火30 s, 72℃延伸1 min, 35个循环 | 407 | 100.18 | [27] |
| nosZ | nosZ-f nosZ-1622R |
CGYTGTTCMTCGACAGCCAG CGSACCTTSTTGCCSTYGCG |
95℃预变性5min, 95℃变性30 s, 60℃退火30 s, 72℃延伸1 min, 35个循环 | 455 | 95.09 | [27] |
| AM真菌:丛枝菌根真菌Arbuscular mycorrhizal fungi;AOA-amoA:氨氧化古菌Ammonia oxidizing archaea;AOB-amoB:氨氧化细菌,Ammonia oxidizing bacteria | ||||||
采用Microsoft Office Excel 2007和SPSS 17.0对数据进行整理和分析, 采用单因素方差分析(One-way ANOVA)和LSD法比较不同处理间的差异显著性(P < 0.05), 采用Pearson相关分析对生物炭施用量、土壤pH值、含水量、容重、土壤硝态氮、铵态氮, 有机碳、土壤各基因丰度与N2O排放通量之间相关性和显著性进行分析, 差异极显著为P < 0.01, 差异显著为P < 0.05。采用Origin 10.0软件绘图。利用R 2.7.1软件里的“gbmplus”统计包, 进行聚类推进树(Aggregated boosted tree, ABT)分析土壤因子、硝化、反硝化功能基因和AM真菌丰度对N2O排放量的相对重要性。
2 结果与分析 2.1 土壤理化性质变化由表 2可知, 不同生物炭处理条件下, 土壤理化性质发生相应变化。随着生物炭施用量的增加, 土壤pH和土壤含水量呈增加趋势, 但均未达到显著水平(P>0.05)。施加生物炭显著降低土壤容重, T1和T2处理土壤容重分别较T0显著降低3.97%和8.78%(P < 0.05);施加生物炭显著提高土壤有机碳含量, T1和T2处理土壤有机碳含量分别较T0显著提高38.44%和71.01%(P < 0.05);与T0相比, T1处理土壤全氮含量未达到显著水平(P>0.05), T2处理显著提高土壤全氮含量, 较T0处理显著提高14.87%(P < 0.05);添加生物炭土壤硝态氮含量显著降低, 与T0相比, T1和T2处理土壤硝态氮含量分别减少10.57%和21.40%(P < 0.05), T1和T2两处理间未达到显著水平(P>0.05)。添加生物炭后土壤铵态氮含量显著高于不添加生物炭处理, T1和T2处理土壤铵态氮含量分别较T0显著增加15.89%和30.46%(P < 0.05)。
| 处理 Treatment |
pH | 土壤含水量 Soil water content/% |
土壤容重 Bulk density/ (g/cm3) |
有机碳 Soil organic carbon/(g/kg) |
全氮 Total N/ (g/kg) |
硝态氮 Nitrate N/ (mg/kg) |
铵态氮 Ammonium N/ (mg/kg) |
| T0 | 7.39±0.24a | 22.53±0.92ab | 1.18±0.13a | 11.84±0.61c | 1.21±0.04b | 27.52±1.53a | 1.51±0.15c |
| T1 | 7.42±016a | 23.24±0.59ab | 1.13±0.08b | 16.39±0.44b | 1.28±0.02ab | 24.61±0.90bc | 1.75±0.09b |
| T2 | 7.48±025a | 23.98±0.50a | 1.07±0.10c | 20.24±0.31a | 1.39±0.03a | 21.63±0.71c | 1.97±0.09a |
| 同一列中不同字母表示不同处理间差异显著(P < 0.05); T0:不施用生物炭No biochar application;T1:施用生物炭20t/hm2 Biochar application rate of 20 t/hm2;T2:施用生物炭40t/hm2 Biochar application rate of 40 t/hm2 | |||||||
由图 1可知, 各处理土壤AOA-amoA基因拷贝数显著高于AOB-amoA。随着生物炭施用量的增加, AOA-amoA基因丰度显著降低(P < 0.05), 表现为T0>T1>T2。T0、T1和T2处理下AOA-amoA基因丰度分别为7.57×109、4.92×109和2.50×109拷贝/g, T1和T2处理分别较T0处理显著降低35.07%和66.96%(P < 0.05)。与T0相比, TI和T2处理的AOB-amoA基因丰度显著下降(P < 0.05), 但T1和T2处理未达到显著水平(P>0.05)。T0、T1和T2处理下AOB-amoA基因丰度分别为3.60×106、2.02×106、2.36×106拷贝/g, T1和T2分别较T0处理显著降低43.83%和34.58%(P < 0.05)。说明生物炭对旱地玉米土壤AOA和AOB生长具有抑制作用。

|
| 图 1 不同生物炭处理下氨氧化细菌、氨氧化古菌基因丰度 Fig. 1 The gene abundance of ammonia-oxidizing bacteria and archaea under different biochar treatments 不同字母表示不同处理间差异显著(P < 0.05); T0:不施用生物炭No biochar application;T1:施用生物炭20t/hm2 Biochar application rate of 20 t/hm2;T2:施用生物炭40t/hm2 Biochar application rate of 40 t/hm2;AOA-amoA:氨氧化古菌Ammonia oxidizing archaea;AOB-amoB:氨氧化细菌Ammonia oxidizing bacteria |
由图 2可知, 随着生物炭施用量的增加, nirK基因丰度显著降低(P < 0.05), 表现为T0>T1>T2。T0、T1和T2处理下nirK基因丰度分别为1.08×1010、7.33×109、6.16×109拷贝/g, T1和T2分别较T0处理显著降低31.90%和42.78%(P < 0.05), T1和T2两处理间无显著差异(P>0.05)。随着生物炭施用量的增加nirS基因丰度降低, 表现为T0>T1>T2, T0、T1和T2处理下nirS基因丰度分别为3.57×107、3.12×107、1.90×107拷贝/g, T1和T0两处理间无显著差异(P>0.05), T2分别较T0和T1处理显著降低46.69%和38.91%(P < 0.05), 说明生物炭能够抑制反硝化反应进程。施加生物炭nosZ基因丰度增加, 表现为T1>T2>T0。T0、T1和T2处理下nosZ基因丰度分别为2.11×107、3.74×107、2.92×107拷贝/g, 以T1处理nosZ基因丰度最高, T1和T2处理分别较T0显著提高77.28%和38.33%(P < 0.05), 说明施用生物炭能够促进nosZ基因表达, 加快了N2O还原进程。(nirK+nirS)/nosZ的比值越大表明排放N2O的能力越强。由图 2可知, 生物炭处理显著降低(nirK+nirS)/nosZ的比值(P < 0.05)。与T0处理相比, T1和T2处理显著降低(nirK+nirS)/nosZ的比值(P < 0.05), 但T1和T2处理之间无显著差异(P>0.05)。

|
| 图 2 不同生物炭处理下反硝化基因丰度 Fig. 2 The gene abundance of denitrifying bacteria under different biochar treatments 不同字母表示不同处理间差异显著(P < 0.05) |
由表 3可知, 随着生物炭施用量的增加, 菌根侵染率随之增加, 与T0相比, T1和T2处理均显著增加菌根侵染率(P < 0.05), 但T1和T2处理差异不显著(P>0.05)。随着生物炭施用量的增加AM真菌基因丰度显著增加(P < 0.05), 表现为T2>T1>T0。T1和T2分别较T0处理显著提高71.88%和115.88%(P < 0.05), 说明生物炭对AM真菌生长具有明显的促进作用。施加生物炭显著提高土壤总球囊霉素相关土壤蛋白和易提取球囊霉素相关土壤蛋白含量, T1和T2处理土壤总球囊霉素相关土壤蛋白含量分别较T0显著提高20.67%和34.36%(P < 0.05), T1和T2处理土壤易提取球囊霉素相关土壤蛋白含量分别较T0显著提高17.40%和43.50%(P < 0.05)。
| 处理 Treatment |
菌根侵染率 Mycorrhizal colonization/% |
AM真菌拷贝数 AM fungi copy numbers/ (106拷贝/g) |
总球囊霉素相关 土壤蛋白含量 T-GRSP content/(mg/g) |
易提取球囊霉素相关 土壤蛋白含量 EE-GRSP content/(mg/g) |
| T0 | 20.39±1.04b | 4.93±0.75c | 1.86 c | 0.47c |
| T1 | 23.42±2.06a | 8.48±0.63b | 2.24 b | 0.55b |
| T2 | 24.48±1.75a | 10.71±0.97a | 2.49 a | 0.67a |
| 同一列中不同字母表示不同处理间差异显著(P < 0.05); AM真菌:丛枝菌根真菌Arbuscular mycorrhizal fungi;EE-GRSP:易提取球囊霉素相关土壤蛋白含量Easily extractable glomalin-related soil protein; T-GRSP:总球囊霉素相关土壤蛋白,Total glomalin-related soil protein | ||||
玉米大喇叭口期追肥后N2O排放通量和累积排放量动态变化如图 3所示。在不同时期, 施加生物炭处理的N2O排放通量均低于不施生物炭处理, N2O排放通量具体表现为:T0>T1>T2。追肥(7月24日)后, 不同处理N2O排放通量的变化趋势基本一致, 均表现为先升高后下降的趋势。追肥(7月24日)后第1天N2O排放量升高, 追肥后第3天出现排放峰最高值, T0、T1、T2处理下N2O排放通量分别为522.14、409.35和382.35 μg m-2 h-1, T1、T2处理分别较T0处理降低21.60%和26.77%。强降雨(8月6日)后第2天, 出现第二次排放峰, T0、T1、T2处理N2O排放通量分别为210.59、158.04、139.63 μg m-2 h-1, T1、T2处理分别较T0处理降低24.95%和33.70%。8月14日和8月21日降雨后又出现两次排放峰。总的来说, 施用生物炭降低旱地玉米土壤N2O排放, 且施肥和降雨后会引起N2O排放量增加。

|
| 图 3 生物炭对旱地玉米农田土壤N2O排放通量的影响 Fig. 3 Effect of biochar on soil N2O emission fluxes of rainfed maize 不同字母表示不同处理间差异显著(P < 0.05) |
由图 3可知, 随着生物炭施用量的增加N2O累积排放量呈减少趋势, 表现为T0>T1>T2。施加生物炭显著降低N2O累积排放量(P < 0.05), T0、T1和T2处理下的N2O累积排放量分别为162.97、136.38、120.56 mg/m2, T1和T2分别较T0处理显著降低16.31%和26.02%(P < 0.05), 说明施用生物炭对降低旱地玉米土壤N2O排放具有积极作用。
2.5 生物炭施用量与AM真菌及N转化相关功能基因丰度的相关性分析如表 4所示, 为生物炭施用量与AM真菌及N转化相关功能基因丰度的相关分析, AM真菌基因丰度与生物炭施用量呈显著正相关关系(P < 0.05);AOA、AOB、nirK、nirS基因丰度与生物炭施用量呈极显著负相关关系(P < 0.01);nosZ与生物炭施用量未达到显著水平(P>0.05)。其中, nirK基因丰度与生物炭施用量的相关性系数最大, 为-0.763, 说明nirK基因丰度受生物炭施用量影响较大。
| AM真菌 AM fungi/ (106 拷贝/g) |
AOA / (拷贝/g) |
AOB / (拷贝/g) |
nirK
/ (拷贝/g) |
nirS
/ (拷贝/g) |
nosZ
/ (拷贝/g) |
|
| 相关系数 Correlation coefficient |
0.368* | -0.587** | -0.480** | -0.763** | -0.626** | 0.285 |
| **表示相关性达0.01极显著水平;*表示相关性达0.05显著水平 | ||||||
由表 5可知, AM真菌、nosZ基因丰度和N2O排放通量呈极显著负相关(P < 0.01), AOA、nirK、nirS和N2O排放通量呈极显著正相关(P < 0.01), AOB与N2O排放通量未达到显著水平(P>0.05)。说明AM真菌、AOA、nosZ、nirK和nirS是影响N2O排放的主要因子。
| AM真菌 AM fungi/ (106 拷贝/g) |
AOA / (拷贝/g) |
AOB / (拷贝/g) |
nirK
/ (拷贝/g) |
nirS
/ (拷贝/g) |
nosZ
/ (拷贝/g) |
|
| 相关系数 Correlation coefficient |
-0.865** | 0.819** | 0.207 | 0.623** | 0.799** | -0.629** |
| **表示相关性达0.01极显著水平;*表示相关性达0.05显著水平 | ||||||
N2O排放通量与土壤各因子的相关性分析如表 6所示。由表 6可知, 土壤含水量与N2O排放通量呈显著正相关关系(P < 0.05), N2O排放峰出现在降雨后, 土壤水分是影响N2O排放的主要因子。土壤容重、pH和总球囊霉素相关土壤蛋白含量与N2O排放通量未达到显著水平(P>0.05);土壤有机碳含量与N2O排放通量呈显著负相关关系(P < 0.05);易提取球囊霉素相关土壤蛋白含量、铵态氮含量与N2O排放通量呈极显著负相关关系(P < 0.01);土壤硝态氮含量与N2O排放通量呈极显著正相关关系(P < 0.01)。其中, 易提取球囊霉素相关土壤蛋白、硝态氮、铵态氮含量与N2O排放通量的相关系数分别为-0.887、0.738、-0.909、, 说明易提取球囊霉素相关土壤蛋白、硝态氮、铵态氮含量是影响N2O排放的主要因子。
| 土壤含水量 Soil water content/% |
土壤容重 Bulk density /(g/cm3) |
pH | 有机碳 Soil organic carbon/ (g/kg) |
硝态氮 Nitrate N /(mg/kg) |
铵态氮 Ammonium N /(mg/kg) |
T-GRSP /(mg/g) |
EE-GRSP /(mg/g) |
|
| 相关系数 Correlation coefficient |
0.503* | 0.387 | -0.355 | -0.565* | 0.738** | -0.909** | 0.311 | -0.887** |
| **表示相关性达0.01极显著水平;*表示相关性达0.05显著水平。T-GRSP:总球囊霉素相关土壤蛋白;EE-GRSP:易提取球囊霉素相关土壤蛋白 | ||||||||
硝化、反硝化功能基因与AM真菌的相关性分析如表 7所示。从表 7中可得知, AM真菌与AOA、AOB、nirK、nirS基因丰度呈极显著负相关(P < 0.01), 与nosZ呈极显著正相关(P < 0.01);AOA与AOB、nirK、nirS基因丰度呈极显著正相关(P < 0.01), 与nosZ呈极显著负相关(P < 0.01);AOB与nirK基因丰度呈极显著正相关(P < 0.01), 与nosZ呈极显著负相关(P < 0.01);nirK与nirS基因丰度呈极显著正相关(P < 0.01), 与nosZ呈极显著负相关(P < 0.01);nirS与nosZ基因丰度呈显著负相关(P < 0.05)。
| AM真菌 AM fungi/ (106 拷贝/g) |
AOA / (拷贝/g) |
AOB / (拷贝/g) |
nirK
/ (拷贝/g) |
nirS
/ (拷贝/g) |
nosZ
/ (拷贝/g) |
|
| AM真菌 AM fungi/(106 拷贝/g) |
1 | |||||
| AOA/(拷贝/g) | -0.715** | 1 | ||||
| AOB/(拷贝/g) | -0.554** | 0.605** | 1 | |||
| nirK/(拷贝/g) | -0.727** | 0.708** | 0.667** | 1 | ||
| nirS/(拷贝/g) | -0.743** | 0.803** | 0.398* | 0.772** | 1 | |
| nosZ/(拷贝/g) | 0.607** | -0.572** | -0.503** | -0.616** | -0. 323* | 1 |
将土壤因子、硝化、反硝化功能基因和AM真菌丰度分别加入ABT模型进行分析(图 4), 结果表明, AOA对N2O排放的影响最大(21.1%), 其次是AM真菌(19.2%)和nirS(11.8%), 其余因子贡献率均不足10%;影响N2O排放量的相对贡献率由大到小依次为:AOA、AM真菌、nirS、nosZ、AOB、铵态氮、土壤含水量、有机碳、硝态氮、nirK、土壤容重、pH。

|
| 图 4 土壤因子、硝化、反硝化功能基因和AM真菌丰度对N2O排放的相对影响的ABT分析 Fig. 4 Relative importance of soil factors, nitrification and denitrification functional gene abundances and AM fungi for N2O emission by the aggregated boosted tree (ABT) model |
许多研究表明, 农田土壤施入生物炭后, 土壤理化性质如pH、含水量、容重、有机碳、氮等发生相应变化[17, 28]。本研究表明, 施入生物炭后, 土壤pH值和土壤含水量呈增加趋势, 但均未达到显著水平, 主要是因为生物炭自身呈碱性, 施入土壤后能够提高土壤pH值, 但本试验地土壤为弱碱性, 具有一定的酸碱缓冲能力, 因此各处理间土壤pH值差异不显著(表 2), 另外生物炭具有较好的亲水性和持水能力, 增加了水分的渗流模式和路径, 减缓了土壤水分的蒸发损失, 引起土壤含水量增加。这和李培培等[28]的研究结果一致。施入生物炭后, 土壤容重呈减少趋势, 这与Baiamonte等[29]的研究结果一致, 主要是由于生物炭具有多孔疏松的结构、大的比表面积, 自身密度较小, 施入土壤之后对土壤具有“稀释效应”[30]。施入生物炭后土壤有机碳的增加主要是由于以下几方面原因:首先是由生物质热解形成的生物炭中的碳主要以惰性的芳香环状结构存在, 本身碳含量非常高, 在土壤中加入生物炭可以提高土壤有机碳的含量;其次是生物炭具有较强的吸附性, 能够吸附土壤有机小分子, 促进有机小分子聚合形成土壤有机质[31], 另外生物炭的多孔结构为微生物的生长繁殖提供附着位点, 为微生物提供了良好的栖息环境, 土壤微生物量增加, 加速了土壤有机态养分的分解释放, 从而增加土壤有机碳含量[17]。本研究表明, 施入生物炭后, 土壤全氮、铵态氮含量增加。这与Nguyen等[32]通过荟萃分析得出的结果不同, Nguyen等研究发现生物炭的高C/N特性及带入的活性物质引起微生物对土壤矿质氮的固定, 因此降低了氮素有效性。与宋大利等[33]和刘遵奇等[34]的研究结果相一致。主要是由于生物炭本身含有一定量的氮素;另外施用生物炭可降低氮素淋失和改善土壤通气状况抑制了微生物的反硝化作用, 从而减少了N2O的形成和排放, 进而使得土壤中全氮含量增加。
生物炭孔隙由于能够贮存水分和养分, 为微生物栖息生活提供了优越的天然微环境, 生物炭施入土壤能够显著增加微生物数量及活性。本研究表明, 施入生物炭能够显著提高AM真菌侵染率和AM真菌基因丰度, 与Hammer等[17]的研究结果一致。主要原因可能为:(1)生物炭改善土壤理化特性有利于AM真菌孢子的萌发和生长, 生物炭多孔结构为AM真菌提供栖息地[15];(2)生物炭调节植物-真菌信号物质影响AM真菌的萌发和菌丝的分枝[35];(3)生物炭可能通过影响土壤中的解磷细菌活性, 从而间接影响AM真菌[36]。AM真菌在氮素吸收利用和土壤氮素循环过程中有着重要的作用, 一方面AM真菌能够通过竞争吸收氮素来调控N2O排放。AM真菌吸收大量氮素用以自身生长需要, 并改善宿主植物的氮素营养[37], 从而减少硝化、反硝化作用底物浓度而降低N2O排放速率;另一方面AM真菌能够改变植物根系分泌物的组成及数量, 从而导致根际微生物群落发生变化, 包括一些硝化、反硝化细菌群落, 从而影响N2O排放[38-39]。
土壤N2O的排放主要是微生物活动过程产生的, 硝化细菌和反硝化细菌的数量是影响N2O排放的关键因素[40]。本试验结果表明, 施用生物炭改变了AOA和AOB的基因丰度, 且AOA和AOB的基因丰度与生物炭施用量呈极显著负相关关系, AOA基因丰度随生物炭施用量的增加表现出显著降低的趋势, 而T1和T2两处理间对AOB的基因丰度无显著影响, 说明增加生物炭施用量对AOB基因丰度的影响较小, 总的来说生物炭抑制了AOA和AOB的生长。这可能是因为生物炭对NH4+-N的吸附以及AM真菌对NH4+-N的吸收固定, 减少了硝化反应的底物浓度, 从而抑制了氨氧化菌的生长[5, 41]。nirK、nirS、nosZ基因丰度的高低直接影响反硝化作用的进程[42], 本研究表明, 生物炭处理降低了nirK和nirS基因丰度, 其中T1和T2处理较T0处理显著提高了nirK基因丰度, 但T1和T2两处理间无显著差异, 而T2处理较T0处理显著增加了nirS基因丰度, T1处理对比T0处理无显著影响, 说明nirK基因型反硝化细菌对低生物炭施用量比较敏感, 而nirS基因型反硝化细菌对高生物炭施用量比较敏感, 生物炭对nirK和nirS基因型反硝化细菌的繁殖有抑制作用。这与刘杏认等[3]的研究结果不同, 可能是因为盆栽试验与大田试验土壤反硝化微生物所处的环境差异较大, 从而使土壤反硝化微生物对生物炭的反映不尽相同。nirK和nirS基因丰度与生物炭施用量呈极显著负相关。虽然生物炭处理增加了nosZ基因丰度, nosZ基因丰度与生物炭施用量呈正相关, 但未达到显著水平。生物炭增加nosZ基因丰度促使N2O的还原, 解释了生物炭对旱地农田土壤N2O减排具有较大潜力。
AM真菌与硝化、反硝化微生物的相互作用对N2O排放具有重要意义。相关分析表明, AM真菌与AOA、AOB、nirK、nirS基因丰度呈极显著负相关(P < 0.01), 与nosZ呈极显著正相关(P < 0.01)。Chen等[43]通过室内盆栽模拟试验研究发现AM真菌可显著降低菌根际土壤AOA、AOB基因丰度, 并且还分别间接、直接影响菌根室、菌丝室中AOA的群落组成。Veresoglou等[44]研究发现接种AM真菌可能降低土壤中AOA、AOB的基因丰度。主要是由于AM真菌、氨氧化微生物(AOA和AOB)均以NH4+-N为底物, 二者在竞争土壤NH4+-N时, 氨氧化微生物(AOA和AOB)一般处于劣势, 在土壤中NH4+较低时, AM真菌优先利用NH4+并抑制氨氧化微生物对NH4+的利用, 造成AOA和AOB丰度和土壤硝化速率的降低, 进而减少了硝化过程中产生的N2O。Bender等[1]研究发现野生型番茄接种AM真菌后土壤中nirK基因丰度和AM真菌侵染率呈线性负相关, 和nosZ基因丰度则呈正相关, 且土壤N2O排放量显著下降。表明AM真菌降低了土壤中硝化、反硝化微生物的基因丰度, 提高了nosZ基因的丰度, 进而降低N2O的排放。集成推进树(ABT)分析表明, AOA对N2O排放的影响最大, 其次是AM真菌、nirS和nosZ, 刘杏仁等[3]研究认为盆栽小麦土壤N2O的排放主要受AOA、nirS和nosZ影响, Liu等[13]研究认为华北农田玉米季N2O的排放主要受AOB和nosZ影响, 主要由于土壤类型和所种植的农作物种类不同, 造成影响N2O排放的主要因子不同。
总的来说, 生物炭处理提高土壤AM真菌侵染率、调节土壤理化性质, 生物炭和AM真菌协同调节土壤微生物, 特别是对硝化、反硝化相关基因的调节, 进而影响土壤N循环和N2O气体的排放, 但生物炭和AM真菌之间的协调机制需要进行更进一步研究。
| [1] |
Bender S F, Plantenga F, Neftel A, Jocher M, Oberholzer H R, Köhl L, Giles M, Daniell T J, van der Heijden M G A. Symbiotic relationships between soil fungi and plants reduce N2O emissions from soil. The ISME Journal, 2014, 8(6): 1336-1345. DOI:10.1038/ismej.2013.224 |
| [2] |
Meng Q F, Yue S C, Hou P, Cui Z L, Chen X P. Improving yield and nitrogen use efficiency simultaneously for maize and wheat in China: a review. Pedosphere, 2016, 26(2): 137-147. DOI:10.1016/S1002-0160(15)60030-3 |
| [3] |
刘杏认, 赵光昕, 张晴雯, 田秀平. 生物炭对华北农田土壤N2O通量及相关功能基因丰度的影响. 环境科学, 2018, 39(8): 3816-3825. |
| [4] |
贾仲君, 翁佳华, 林先贵, Conrad R. 氨氧化古菌的生态学研究进展. 微生物学报, 2010, 50(4): 431-437. |
| [5] |
陈永亮, 陈保冬, 刘蕾, 胡亚军, 徐天乐, 张莘. 丛枝菌根真菌在土壤氮素循环中的作用. 生态学报, 2014, 34(17): 4807-4815. |
| [6] |
Kang F R, Yang B, Wujisiguleng, Yang X, Wang L, Guo J X, Sun W, Zhang Q, Zhang T. Arbuscular mycorrhizal fungi alleviate the negative effect of nitrogen deposition on ecosystem functions in meadow grassland. Land Degradation & Development, 2020, 31(6): 748-759. |
| [7] |
Qiu Y P, Jiang Y, Guo L J, Zhang L, Burkey K O, Zobel R W, Reberg-Horton S C, Shew H D, Hu S J. Shifts in the composition and activities of denitrifiers dominate CO2 stimulation of N2O emissions. Environmental Science & Technology, 2019, 53(19): 11204-11213. |
| [8] |
Zhang H, Voroney R P, Price G W. Effects of temperature and processing conditions on biochar chemical properties and their influence on soil C and N transformations. Soil Biology and Biochemistry, 2015, 83: 19-28. DOI:10.1016/j.soilbio.2015.01.006 |
| [9] |
陈温福, 张伟明, 孟军. 农用生物炭研究进展与前景. 中国农业科学, 2013, 46(16): 3324-3333. DOI:10.3864/j.issn.0578-1752.2013.16.003 |
| [10] |
Li S L, Wang S, Fan M C, Wu Y, Shangguan Z P. Interactions between biochar and nitrogen impact soil carbon mineralization and the microbial community. Soil and Tillage Research, 2020, 196: 104437. DOI:10.1016/j.still.2019.104437 |
| [11] |
刘杏认, 张星, 张晴雯, 李贵春, 张庆忠. 施用生物炭和秸秆还田对华北农田CO2、N2O排放的影响. 生态学报, 2017, 37(20): 6700-6711. |
| [12] |
姬强, 马媛媛, 刘永刚, 王锐, 孙权. 秸秆生物质炭对土壤结构体与活性碳分布、转化酶动力学参数及小麦生长的影响. 生态学报, 2019, 39(12): 4366-4375. |
| [13] |
Liu X R, Ren J Q, Zhang Q W, Liu C. Long-term effects of biochar addition and straw return on N2O fluxes and the related functional gene abundances under wheat-maize rotation system in the North China Plain. Applied Soil Ecology, 2019, 135: 44-55. DOI:10.1016/j.apsoil.2018.11.006 |
| [14] |
Gomez J D, Denef K, Stewart C E, Zheng J, Cotrufo M F. Biochar addition rate influences soil microbial abundance and activity in temperate soils. European Journal of Soil Science, 2014, 65(1): 28-39. DOI:10.1111/ejss.12097 |
| [15] |
Hammer E C, Forstreuter M, Rillig M C, Kohler J. Biochar increases arbuscular mycorrhizal plant growth enhancement and ameliorates salinity stress. Applied Soil Ecology, 2015, 96: 114-121. DOI:10.1016/j.apsoil.2015.07.014 |
| [16] |
Zhang L, Qiu Y P, Cheng L, Wang Y, Liu L L, Tu C, Bowman D C, Burkey K O, Bian X M, Zhang W J, Hu S J. Atmospheric CO2 enrichment and reactive nitrogen inputs interactively stimulate soil cation losses and acidification. Environmental Science & Technology, 2018, 52(12): 6895-6902. |
| [17] |
Hammer E C, Balogh-Brunstad Z, Jakobsen I, Olsson P A, Stipp S L S, Rillig M C. A mycorrhizal fungus grows on biochar and captures phosphorus from its surfaces. Soil Biology and Biochemistry, 2014, 77: 252-260. DOI:10.1016/j.soilbio.2014.06.012 |
| [18] |
张学林, 徐钧, 安婷婷, 侯小畔, 李潮海. 不同氮肥水平下玉米根际土壤特性与产量的关系. 中国农业科学, 2016, 49(14): 2687-2699. DOI:10.3864/j.issn.0578-1752.2016.14.004 |
| [19] |
Kalembasa S J, Jenkinson D S. A comparative study of titrimetric and gravimetric methods for the determination of organic carbon in soil. Journal of the Science of Food and Agriculture, 1973, 24(9): 1085-1090. DOI:10.1002/jsfa.2740240910 |
| [20] |
Jackson M L. Soil Chemical Analysis. Englewood Cliffs: Prentice-Hall, 1973.
|
| [21] |
Biermann B, Linderman R G. Quantifying vesicular-arbuscular mycorrhizae: a proposed method towards standardization. New Phytologist, 1981, 87(1): 63-67. DOI:10.1111/j.1469-8137.1981.tb01690.x |
| [22] |
李少朋, 毕银丽, 陈昢圳, Yryszhan Z, 刘生. 外源钙与丛枝菌根真菌协同对玉米生长的影响与土壤改良效应. 农业工程学报, 2013, 29(1): 109-116. |
| [23] |
van Geel M, Busschaert P, Honnay O, Lievens B. Evaluation of six primer pairs targeting the nuclear rRNA operon for characterization of arbuscular mycorrhizal fungal (AMF) communities using 454 pyrosequencing. Journal of Microbiological Methods, 2014, 106: 93-100. DOI:10.1016/j.mimet.2014.08.006 |
| [24] |
Francis C A, Roberts K J, Beman J M, Santoro A E, Oakley B B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(41): 14683-14688. DOI:10.1073/pnas.0506625102 |
| [25] |
Rotthauwe J H, Witzel K P, Liesack W. The ammonia monooxygenase structural gene amoA as a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations. Applied and Environmental Microbiology, 1997, 63(12): 4704-4712. DOI:10.1128/AEM.63.12.4704-4712.1997 |
| [26] |
Braker G, Fesefeldt A, Witzel K P. Development of PCR primer systems for amplification of nitrite reductase genes (nirK and nirS) to detect denitrifying bacteria in environmental samples. Applied and Environmental Microbiology, 1998, 64(10): 3769-3775. DOI:10.1128/AEM.64.10.3769-3775.1998 |
| [27] |
Throbäck I N, Enwall K, Jarvis Å, Hallin S. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE. FEMS Microbiology Ecology, 2004, 49(3): 401-417. DOI:10.1016/j.femsec.2004.04.011 |
| [28] |
李培培, 汪强, 文倩, 李慧, 吴传发, 熊伟东, 韩燕来. 不同还田方式对砂质潮土理化性质及微生物的影响. 生态学报, 2017, 37(11): 3665-3672. |
| [29] |
Baiamonte G, Crescimanno G, Parrino F, De Pasquale C. Effect of biochar on the physical and structural properties of a sandy soil. Catena, 2019, 175: 294-303. DOI:10.1016/j.catena.2018.12.019 |
| [30] |
韩召强, 陈效民, 曲成闯, 张晓玲, 张俊, 黄春燕, 刘云梅. 生物质炭对黄瓜连作土壤理化性状、酶活性及土壤质量的持续效应. 植物营养与肥料学报, 2018, 24(5): 1227-1236. |
| [31] |
Grunwald D, Kaiser M, Junker S, Marhan S, Piepho H P, Poll C, Bamminger C, Ludwig B. Influence of elevated soil temperature and biochar application on organic matter associated with aggregate-size and density fractions in an arable soil. Agriculture, Ecosystems & Environment, 2017, 241: 79-87. |
| [32] |
Nguyen T T N, Xu C Y, Tahmasbian I, Che R X, Xu Z H, Zhou X H, Wallace H M, Bai S H. Effects of biochar on soil available inorganic nitrogen: a review and meta-analysis. Geoderma, 2017, 288: 79-96. DOI:10.1016/j.geoderma.2016.11.004 |
| [33] |
宋大利, 习向银, 黄绍敏, 张水清, 袁秀梅, 黄伏森, 刘阳, 王秀斌. 秸秆生物炭配施氮肥对潮土土壤碳氮含量及作物产量的影响. 植物营养与肥料学报, 2017, 23(2): 369-379. |
| [34] |
刘遵奇, 兰宇, 杨铁鑫, 张艺潇, 孟军. 减肥条件下生物炭施用方式对土壤肥力及酶活性的影响. 农业资源与环境学报, 2020, 37(4): 544-551. |
| [35] |
Atkinson C J, Fitzgerald J D, Hipps N A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: a review. Plant and Soil, 2010, 337(1/2): 1-18. |
| [36] |
Warnock D D, Lehmann J, Kuyper T W, Rillig M C. Mycorrhizal responses to biochar in soil-concepts and mechanisms. Plant and Soil, 2007, 300(1/2): 9-20. |
| [37] |
Hodge A, Fitter A H. Substantial nitrogen acquisition by arbuscular mycorrhizal fungi from organic material has implications for N cycling. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(31): 13754-13759. DOI:10.1073/pnas.1005874107 |
| [38] |
韦莉莉, 卢昌熠, 丁晶, 俞慎. 丛枝菌根真菌参与下植物-土壤系统的养分交流及调控. 生态学报, 2016, 36(14): 4233-4243. |
| [39] |
赵兰凤, 张新明, 程根, 张丽娟, 刘小锋, 李华兴. 生物炭对菜园土壤微生物功能多样性的影响. 生态学报, 2017, 37(14): 4754-4762. |
| [40] |
Cui X Q, Hao H L, Zhang C K, He Z L, Yang X E. Capacity and mechanisms of ammonium and cadmium sorption on different wetland-plant derived biochars. Science of the Total Environment, 2016, 539: 566-575. DOI:10.1016/j.scitotenv.2015.09.022 |
| [41] |
Li S L, Wang S, Shangguan Z P. Combined biochar and nitrogen fertilization at appropriate rates could balance the leaching and availability of soil inorganic nitrogen. Agriculture, Ecosystems & Environment, 2019, 276: 21-30. |
| [42] |
Ji C, Li S Q, Geng Y J, Miao Y C, Ding Y, Liu S W, Zou J W. Differential responses of soil N2O to biochar depend on the predominant microbial pathway. Applied Soil Ecology, 2020, 145: 103348. DOI:10.1016/j.apsoil.2019.08.010 |
| [43] |
Chen Y L, Chen B D, Hu Y J, Li T, Zhang X, Hao Z P, Wang Y S. Direct and indirect influence of arbuscular mycorrhizal fungi on abundance and community structure of ammonia oxidizing bacteria and archaea in soil microcosms. Pedobiologia, 2013, 56(4/6): 205-212. |
| [44] |
Veresoglou S D, Sen R, Mamolos A P, Veresoglou D S. Plant species identity and arbuscular mycorrhizal status modulate potential nitrification rates in nitrogen-limited grassland soils. Journal of Ecology, 2011, 99(6): 1339-1349. DOI:10.1111/j.1365-2745.2011.01863.x |