 2021, Vol. 41
2021, Vol. 41文章信息
- 刘媛媛, 李霞, 王小博, 王绍强, 钱钊晖
- LIU Yuanyuan, LI Xia, WANG Xiaobo, WANG Shaoqiang, QIAN Zhaohui
- 2001-2018年中国-老挝交通走廊核心区植被稳定性对极端干旱的响应
- Vegetation stability in response to extreme droughts from 2001 to 2018 in the core area of China-Laos transportation corridors
- 生态学报. 2021, 41(7): 2537-2547
- Acta Ecologica Sinica. 2021, 41(7): 2537-2547
- http://dx.doi.org/10.5846/stxb202002190295
-
文章历史
- 收稿日期: 2020-02-19
- 网络出版日期: 2021-01-27





2. 中国科学院大学资源与环境学院, 北京 100049;
3. 生态环境部对外合作与交流中心, 北京 100035
2. College of Resources and Environment at University of Chinese Academy of Sciences, Beijing 100049, China;
3. Foreign Environmental Cooperation Center, Ministry of Ecology and Environment of the People's Republic of China, Beijing 100035, China
干旱是一种水分持续亏缺的异常气象现象, 会对全球生态环境、社会、经济和农业造成重大负面影响[1-2]。相比洪涝、高温与地震等自然灾害, 干旱影响的范围更广, 造成的损失更大[3-4]。过去一个世纪以来, 在全球范围内极端气候事件的发生频率与严重程度不断提高[5-9]。研究干旱的时空分布格局不仅可为了解干旱的发生机制提供理论依据, 还能为预防干旱灾害提供有效措施[10]。同时, 干旱严重影响陆地生态系统功能的稳定性, 如光合能力、生态系统生产力和陆地碳汇[11]。目前, 基于野外实验与遥感方法的很多研究表明, 干旱对植被生长和陆地生态系统造成了严重的负面影响[7, 12-13]。因此, 进一步了解植被对干旱的响应非常重要, 可为提高陆地生态系统的稳定性提供理论依据。
目前, 多种干旱指数已被用于全球或区域干旱监测[14-16]。Palmer干旱指数(Palmer Drought Severity Index, PDSI)自提出以来, 已被广泛应用于气象、水文和生态等研究领域, 在评估区域水分平衡与干旱严重程度等方面得到了良好的应用[17-20]。此外, 很多研究应用PDSI分析了干旱在不同区域对植被生长的影响[9, 21]。Zhang等[21]利用PDSI和归一化植被指数NDVI研究了中国干旱的分布情况和干旱对植被生产力的影响。Dong等[9]利用植被指数和自校准PDSI分析了美国加利福尼亚州的严重干旱对植被的影响。
中老交通走廊核心区位于中国云南与老挝接壤地区, 拥有丰富的自然资源。该地区虽然有较高的年降水量, 但植被对日益严重的干旱较为敏感。中老交通走廊核心区主要的植被类型是常绿阔叶林与木本稀树草原, 覆盖面积占总面积的90%以上。近期研究表明, 相比其他植被类型, 常绿阔叶林的干旱情况有加重的趋势[22], 且会经历更长时间的干旱, 可能会面临更大的干旱风险[23-24]。此外, 中老交通走廊核心区所处的老挝北部山区是老挝森林砍伐率最高的地区[25], 建设铁路和水电站等基础设施会加剧该地区的森林砍伐, 可能导致该地植被的稳定性下降。研究区的植被状况受到气候变化与人类活动的影响, 会改变湄公河流域的水文条件[26]。作为湄公河流域的上游, 研究区的植被状况是影响整个流域生态环境质量的关键因素[27]。
植被的稳定性为植被指数的均值与标准差之比。植被的稳定性越高, 抵御干旱的能力越强。植被的稳定性通常用抵抗力和恢复力来评估。抵抗力量化了干旱对生态系统抵抗力的直接影响, 表现为干旱期间植被维持其原始水平的能力[28]。恢复力定义了干旱后生态系统功能恢复到正常状态的速度[29]。目前, 已有多个研究通过实验或模型分析了植被对干旱的抵抗力和恢复力[29-30], 但在研究区所在的湄公河流域开展植被对干旱的抵抗力和稳定性的研究很少。因此, 研究中老交通走廊核心区的干旱时空分布特征以及植被的稳定性、抵抗力和恢复力对干旱的响应十分必要, 有助于提高陆地生态系统的稳定性和抗灾能力。
本研究根据1980—2018年的PDSI数据确定了中老交通走廊核心区的干旱事件, 并对PDSI做了趋势分析。基于2001—2018年的年均EVI数据, 量化了中老交通走廊核心区植被的稳定性(年均EVI与其标准差之比)及其对干旱的抵抗力和恢复力。本文研究目标包括:1) 阐明中老交通走廊核心区极端干旱事件的时空分布特征;2) 评估研究区不同植被类型的稳定性及其对极端干旱事件的抵抗力和恢复力。
1 数据与方法 1.1 研究区中老交通走廊核心区是指中老交通走廊(主要是昆曼公路中国-老挝段和中老铁路)经过地区。研究区包括中国云南省的西双版纳傣族自治州1个地级行政区以及老挝北部的琅南塔省、博胶省、乌多姆塞省、琅勃拉邦、万象省和万象市6个省级行政区(图 1), 总面积约8.4万km2, 地理坐标为99.9—103.4°E, 17.8—22.6°N, 中老边境长约710 km。地貌类型以山地和高原为主, 高程在145—2415 m之间。研究区地处亚热带季风和热带季风两个气候区, 年均日最高温约28.5℃, 多年平均降水量约1690 mm, 雨量充沛但分布不均。该地区气候终年温暖湿润, 无四季之分, 可分为旱季和雨季两个季节, 其中11月至次年4月是旱季, 5月至10月是雨季[31]。研究区的土地覆盖类型以常绿阔叶林和木本稀树草原为主, 分别占研究区总面积的57.42%和33.75%, 而草地、农田和城市仅占研究区总面积的4.09%、2.79%和0.17%(图 1)。

|
| 图 1 研究区地理位置及土地覆盖类型 Fig. 1 Location and land use types of the study area |
本研究采用的土地覆盖数据为MODIS卫星的MCD12Q1产品(https://ladsweb.modaps.eosdis.nasa.gov/search/), 其空间分辨率为500 m, 时间为2017年。本研究在植被覆盖区开展, 研究区不包括水体和城市。基于MODIS土地覆盖产品, 本文选取了研究区5类主要的植被类型作为研究对象, 分别是常绿阔叶林(EBF)、混交林(MF)、木本稀树草原和稀树草原(SAV)、草地(GRA)和农田(GRO)。
目前, NDVI是应用最广泛的植被指数[32-33]。然而, 相比于过去常用的NDVI, EVI增加了对植被的监测能力且能更好地反映植被的生长覆盖状态[34-35], 目前已被广泛应用于许多研究[36-37]。本研究所采用的EVI数据为MODIS卫星的MOD13Q1产品(https://ladsweb.modaps.eosdis.nasa.gov/search/), 其空间分辨率250 m×250 m, 时间分辨率16天, 时间范围为2001—2018年。利用MODIS MRT(MODIS Reprojection Tools)处理工具对MOD13Q1产品进行数据格式转换和投影转换等预处理操作。为了消除异常值的影响, 采用最大合成法(MVC)制作EVI月值序列, 最后计算逐年EVI均值。
Palmer气象干旱指数(Palmer Drought Severity Index, PDSI) [14]已被广泛用于全球或区域干旱监测中, 是中长期干旱监测的重要指标之一。在本研究中, PDSI、降水量和日最高温数据均来自TerraClimate月尺度气象数据集[38], 时间范围为1980—2018年, 空间分辨率约为2.5 arc minutes。根据PDSI的值, 旱涝从极端湿润到极端干旱共分为10个等级(表 1)[14]。极端干旱事件可根据受旱面积比例和干旱程度来统计[39], 因此, 本文综合考虑极端干旱面积比例和PDSI值选出2001—2018年的极端干旱年份。PDSI、气温、降水和土地覆盖数据经投影与裁剪等预处理后统一重采样为250 m空间分辨率的栅格数据。
| Palmer气象干旱指数PDSI Palmer Drought Severity Index |
类型 Type |
Palmer气象干旱指数PDSI Palmer Drought Severity Index |
类型 Type |
|
| ≥ 4.00 | 极端湿润 | -0.50—0.99 | 初始干旱 | |
| 3.00—3.99 | 严重湿润 | -1.00—1.99 | 轻微干旱 | |
| 2.00—2.99 | 中等湿润 | -2.00—2.99 | 中等干旱 | |
| 1.00—1.99 | 轻微湿润 | -3.00—3.99 | 严重干旱 | |
| 0.50—0.99 | 初始湿润 | ≤ -4.00 | 极端干旱 | |
| 0.49—0.49 | 正常 |
植被的稳定性定义为Ym/δ, 其中Ym是所有年份的年均EVI, δ是相同时间间隔下EVI的标准差。抵抗力和恢复力被用于评估极端气候对植被稳定性的影响, 其中, 抵抗力指植被结构和功能保持其原始水平的能力, 恢复力指植被恢复到干旱前水平的速度[40]。EVI可以用来表征植被的结构和光能利用效率, 且MODIS产品具有很高的时间分辨率, 因此, EVI是表征植被的稳定性及其对极端干旱的抵抗力和恢复力的一个良好指标。同之前研究的方法相似[11, 41], 抵抗力指数(Ω)和恢复力指数(Δ)的计算公式如下:
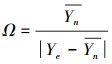
|
(1) |

|
(2) |
式中, Yn代表正常年份(所有非极端年份EVI的均值), Ye代表极端干旱发生年份的EVI, Yd代表最干旱年份的EVI, Ym代表所有年份的年均EVI。植被的EVI值在最干旱年份受到的扰动越小, 其恢复力越好[41], 因此, 本文采用上述恢复力指数分析植被的恢复力。本研究中的抵抗力指数和恢复力指数是无单位的, 因此可在不同植被类型之间直接比较。抵抗力越大, 干旱期间EVI减少的越小。恢复力越大, 最干旱年份EVI受到的扰动越小。抵抗力指数和恢复力指数越高, 表明抵抗力和恢复力越高, 植被越稳定。抵抗力指数和恢复力指数越低, 植被越不稳定, 越易受到干旱的影响。
1.4 统计分析本研究运用单因素方差分析(One-way ANOVA)分析不同植被类型稳定性、抵抗力和恢复力之间的差异, 显著性水平设置为0.05, P<0.05被认为具有统计学意义。
2 结果 2.1 干旱事件的年际分布1980—2018年间, 中老交通走廊核心区的年均PDSI值在-3.73至2.52间大幅波动(图 2)。根据PSDI值和极端干旱面积占比, 中老交通走廊核心区2010年、1993年、1998年和1992年发生的干旱在1980—2018年间最为极端。其中, PDSI在2010年达到最低值, 其次为1993年、1998年和1992年, 以上4年极端干旱面积分别占总面积的47.46%、15.10%、14.94%和10.73%(图 2)。旱季的PDSI值介于-3.83和1.95之间, 最严重的旱季干旱发生在2004年, 其次为1993年和1980年。雨季PDSI值介于-4.60和3.41之间, 最严重的雨季干旱发生在1992年, 其次为2010年和1983年(图 2)。

|
| 图 2 1980—2018年年均PDSI及干旱面积比例 Fig. 2 The mean PDSI and percentage of dry area from 1980—2018 PDSI: Palmer干旱指数Palmer Drought Severity Index |
2001—2018年间, 出现极端干旱的年份有2005年、2010年、2015年、2016年和2017年, 以上年份极端干旱面积占总面积的比例分别为13.37%、47.46%、10.41%、12.00%和3.05%。如图 2所示, 2005年和2010年在旱季和雨季都存在严重和极端干旱, 但2015年和2016年仅在雨季存在大面积的严重与极端干旱, 而旱季严重和极端干旱占比较低。从植被EVI的变化来看(图 3), 2005年和2010年极端干旱导致EVI较相邻年EVI有明显降低, 而2015年和2016年极端干旱对EVI的影响不大, 常绿阔叶林、木本稀树草原和混交林的EVI甚至比相邻年的EVI还高, 植物没有受到干旱的不利影响, 这说明2015年和2016年仅是气象干旱年, 主要是雨季气象干旱, 对植物来讲不是极端干旱年。综合考虑旱季和雨季的极端干旱面积比例和PDSI值, 将2005年和2010年定义为2001—2018年间的极端干旱年份(图 2)。

|
| 图 3 2001—2018年不同植被类型的年均EVI值和PDSI值 Fig. 3 The annual mean PDSI value in different vegetation types from 2001—2018 |
1980—2018年期间, 中老交通走廊旱季和雨季干旱的空间分布差异不明显, PDSI低值主要出现在博胶省、琅南塔省、西双版纳傣族自治州和万象省西南部, 说明以上地区的干旱程度较重(图 4)。图 3为中老交通走廊核心区各植被类型PDSI的年际变化。总的来说, 常绿阔叶林和稀树草原的年均PDSI较高, 而混交林和农田的年均PDSI较低(图 3)。为了探讨PDSI与气象要素的关系, 本文进一步研究了温度和降水量与PDSI的相关性。中老交通走廊核心区各植被类型的年均PDSI与年降水量均呈显著正相关(P<0.01), 说明降水量是影响PDSI变化的关键因素之一。年均日最高温与年均PDSI呈显著负相关(P<0.01), 说明该地区的温度升高也是造成干旱的主要原因(表 2)。

|
| 图 4 1980—2018年年均PDSI值的空间分布 Fig. 4 Spatial distribution of average PDSI values from 1980—2018 |
| 因素Factor | 常绿阔叶林EBF | 混交林MF | 稀树草原SAV | 草地GRA | 农田GRO |
| 年均PDSI与年降水量的相关性Correlation between PDSI and annual accumulative precipitation | 0.643** | 0.569** | 0.614** | 0.560** | 0.505** |
| 年均PDSI与年均日最高温的相关性Correlation between PDSI and annual mean maximum temperature | -0.529** | -0.495** | -0.537** | -0.583** | -0.590** |
| *P<0.05;**P<0.01;EVI: 增强型植被指数Enhanced Vegetation Index; EBF: 常绿阔叶林evergreen broadleaf forest, MF: 混交林mixed forest, SAV: 木本稀树草原和稀树草原savannah, GRA: 草地grassland, GRO: 农田cropland | |||||
中老交通走廊植被的稳定性普遍较高, 且表现出较大的空间变异性(图 5)。研究区常绿阔叶林和稀树草原的稳定性较高, 而混交林、草地和农田的稳定性较低(图 5;图 6)。结果表明, 相比其他植被类型, 常绿阔叶林和稀树草原对干旱的稳定性明显较高(P<0.01, 图 6;表 3)。

|
| 图 5 增强型植被指数(EVI)的稳定性、EVI对干旱的抵抗力和恢复力 Fig. 5 Temporal stability of Enhanced Vegetation Index, resistance, and resilience of EVI to droughts |

|
| 图 6 不同植被类型的EVI对干旱的稳定性, 抵抗力和恢复力(平均值±标准方差) Fig. 6 Comparisons of temporal stability, resistance, and resilience of EVI to droughts among different vegetation types |
| 因素Factor | 植被类型Vegetation types | 常绿阔叶林EBF | 混交林MF | 稀树草原SAV | 草地GRA | 农田GRO |
| 稳定性 | 常绿阔叶林EBF | — | <0.001 | <0.001 | <0.001 | <0.001 |
| Stability | 稀树草原SAV | <0.001 | <0.001 | — | <0.001 | <0.001 |
| 抵抗力 | 常绿阔叶林EBF | — | <0.001 | <0.001 | <0.001 | <0.001 |
| Resistance | 稀树草原SAV | <0.001 | <0.001 | — | <0.001 | <0.001 |
| 恢复力 | 常绿阔叶林EBF | — | <0.001 | <0.001 | <0.001 | <0.001 |
| Resilience | 稀树草原SAV | <0.001 | <0.001 | — | <0.001 | <0.001 |
| 表中数值为单因素方差分析(one-way ANOVA)的结果(P值) | ||||||
植被对干旱的抵抗力在研究区内也表现出较大的空间变异性(图 5), 常绿阔叶林和稀树草原的抵抗力较高, 而混交林、草地和农田的抵抗力较低(图 5;图 6)。相比其他植被类型, 常绿阔叶林和稀树草原植被类型均明显表现出较高的抵抗力(P<0.001, 图 5;表 3)。植被对干旱的抵抗力与年均日高温和年均降水量呈显著正相关(P<0.01, 表 4)。
| 变量Variable | 抵抗力Resistance | 恢复力Resilience |
| 年均日高温Mean annual maximum temperature | 0.0122** | -0.1187** |
| 年均降水量Mean annual precipitation | 0.0225** | 0.0784** |
| *P<0.05;**P<0.01 | ||
植被对干旱的恢复力在研究区不同植被类型之间也具有显著差异。其中, 常绿阔叶林和稀树草原的恢复力较高, 混交林、草地和农田的恢复力较低(图 5;图 6)。研究区内常绿阔叶林和稀树草原比其他植被类型也表现出较高的恢复力(P<0.001, 图 6;表 3)。
3 讨论如图 2所示, 2001—2018年中老交通走廊核心区经历了5次极端干旱事件, 出现极端干旱的年份有2005年、2010年、2015年、2016年和2017年, 其中严重干旱面积分布最广的年份为2010年, 这与Guo等[42]和Son等[31]的研究结果一致。本研究发现降水减少和温度升高是影响中老交通走廊核心区干旱的主要原因(表 3)。
如图 6所示, 常绿阔叶林对干旱的抵抗力高于木本稀树草原、混交林、草地和农田, 表明中老交通走廊核心区的常绿阔叶林具有较高的抗旱能力。研究表明, 植被生产力和陆地碳汇的时间变异性主要是由非常绿生态系统造成的, 如草地、农田和落叶林等[43]。Huang等[11]发现在全球尺度上常绿阔叶林比其他植被类型具有更高的稳定性。常绿阔叶林具有较高的抗旱能力是由多种因素造成的。首先, 从植物根系看, 林木的根系发达, 对干旱造成的水分缺失有较强的抵抗力, 在严重干旱时可利用深层土壤的水分。因此, 林地对干旱的抵抗力相对较强。而草本植物根系较浅, 只能利用表层土壤水[44], 容易受到水分缺失的影响, 因而对干旱更为敏感, 对干旱的抵抗力较弱。农作物与草本植物一样根系较浅, 且其稳定性和对干旱的抵抗力会受人类管理措施的影响。中老交通走廊核心区地形以山地为主, 缺乏灌溉条件, 导致农田对干旱的抵抗力较低。其次, 从植物生理来看, 相比水分利用, 太阳辐射是限制常绿阔叶林生长的主要因素[45], 基于遥感和野外实验的多种研究表明, 常绿阔叶林新叶发育和叶面积指数增加的主要驱动因素是旱季较高的太阳辐射[45]。常绿阔叶林旱季增长的光能利用效率(LUE)可能会增强植物对干旱的抵抗力。在旱季干旱前, 常绿阔叶林会保留新叶去掉旧叶, 从而维持较高的光能利用效率[11, 45]。以上原因均会造成常绿阔叶林比其他植被类型更加稳定和抗旱。
温度升高造成的热浪等极端气候事件通过减少水的供应和生态系统二氧化碳的净交换量, 降低了植被的初级生产力, 由此产生的干旱胁迫可能会进一步降低植物的生存能力[46]。然而, 本研究中, 植被对极端干旱的抵抗力与日高温呈正相关(表 4), 说明温度升高造成干旱(表 2)的同时也提高了植被对极端干旱的抵抗力。该结果可能是由于大多数常绿阔叶林的季节最高温仍低于冠层光合能力的最适温度[47], 因此, 尽管温度升高会造成干旱, 但大气温度与常绿阔叶林最适温度间的差距会防止温度升高对生态系统光合作用造成负面影响。此外, 日最高温对植被生长的影响具有明显的时滞效应[48], 因此, 温度升高对植物的影响很复杂, 具有一定的不确定性。
常绿阔叶林是中老交通走廊核心区的主要植被类型(图 1), 常绿阔叶林的高稳定性表明其在维持该地区生态系统稳定性上发挥了关键作用。在未来干旱日益严重的情景下, 常绿阔叶林的稳定性是否会下降仍有待进一步研究。中老交通走廊核心区所处的北部山区是老挝森林砍伐率最高的地区[25], 加强该地区森林的合理开发利用和提高对常绿阔叶林的保护力度十分重要。
总体而言, 本研究为研究中老交通走廊核心区植被稳定性对极端干旱的响应提供了参考, 为理解干旱的机理和影响因素、提高陆地生态系统的稳定性和恢复力提供了理论依据。然而, 本研究仍存在一定的局限性。例如, 尽管遥感提供了一种研究大尺度植被与干旱关系的方法, 但其更深层的响应机制需要做进一步的实地观察与控制实验。本文仅考虑了气象干旱对植被的影响, 缺少对土壤干旱的分析, 而热带和亚热带季风区的降水较少时, 极有可能土壤并不缺水, 植物仍正常生长, 因此, 土壤干旱对植被生长的影响仍有待进一步研究。此外, 本研究没有考虑长期干旱造成的滞后效应, 如干旱导致的树木死亡和火灾、干旱对植被的滞后效应和累积效应等。土壤、地形、地质和社会经济发展等因素对干旱的影响也有待进一步研究。为了保障中老交通走廊核心区的安全建设, 可根据当地情况采取短期或长期风险缓解措施, 从而有效缓解干旱对该地区造成的损失。
4 结论本文基于PDSI数据, 分析了1980—2018年中老交通走廊核心区干旱的时空分布特征;此外, 基于PDSI和EVI数据, 量化了中老交通走廊核心区不同植被类型的稳定性(年均EVI与其标准差之比)及其对干旱的抵抗力(干旱期间植被结构和功能保持其原始水平的能力)和恢复力(植被恢复到干旱前水平的速度)。得出以下结论:
(1) 2001—2018年间, 中老交通走廊核心区共发生5次极端干旱事件, 出现极端干旱事件的年份有2005年、2010年、2015年、2016年和2017年, 以上年份极端干旱面积占总面积的比例分别为13.37%、47.46%、10.41%、12.00%和3.05%, 其中极端干旱面积分布最广的年份为2010年。
(2) 1980—2018年期间, 中老交通走廊旱季和雨季干旱的空间分布差异不明显, 博胶省、琅南塔省、西双版纳傣族自治州和万象省西南部的干旱程度较重。降水减少和温度升高是造成该地区干旱的主要原因。
(3) 中老交通走廊核心区内常绿阔叶林的稳定性显著高于木本稀树草原、混交林、草地和农田。和其他植被类型相比, 常绿阔叶林虽然暴露在干旱环境中的时间更长, 但其对干旱事件具有最高的抵抗力和恢复力, 在维持中老交通走廊核心区的生态系统稳定性上发挥着重要作用。研究区内各植被类型对干旱的抵抗力与温度和降水呈显著正相关关系。
| [1] |
Piao S L, Ciais P, Huang Y, Shen Z H, Peng S S, Li J S, Zhou L P, Liu H Y, Ma Y C, Ding Y H, Friedlingstein P, Liu C Z, Tan K, Yu Y Q, Zhang T Y, Fang J Y. The impacts of climate change on water resources and agriculture in China. Nature, 2010, 467(7311): 43-51. DOI:10.1038/nature09364 |
| [2] |
Heddinghaus T R, Sabol P. A review of the Palmer Drought Severity Index and where do we go from here?//Proceedings of the 7th Conference on Applied Climatology. Boston, MA: American Meteorological Society, 1991, 242-246. |
| [3] |
Wilhite D A, Sivakumar M V K, Pulwarty R. Managing drought risk in a changing climate: the role of national drought policy. Weather and Climate Extremes, 2014, 3: 4-13. DOI:10.1016/j.wace.2014.01.002 |
| [4] |
Mishra A K, Singh V P. A review of drought concepts. Journal of Hydrology, 2010, 391(1/2): 202-216. |
| [5] |
John R, Chen J Q, Ou-Yang Z T, Xiao J F, Becker R, Samanta A, Ganguly S, Yuan W P, Batkhishig O. Vegetation response to extreme climate events on the Mongolian Plateau from 2000 to 2010. Environmental Research Letters, 2013, 8: 035033. DOI:10.1088/1748-9326/8/3/035033 |
| [6] |
Schwalm C R, Williams C A, Schaefer K, Baldocchi D, Black T A, Goldstein A H, Law B E, Oechel W C, Paw U K T, Scott R L. Reduction in carbon uptake during turn of the century drought in western North America. Nature Geoscience, 2012, 5(8): 551-556. DOI:10.1038/ngeo1529 |
| [7] |
Zhang L, Xiao J F, Li J, Wang K, Lei L P, Guo H D. The 2010 spring drought reduced primary productivity in southwestern China. Environmental Research Letters, 2012, 7(4): 045706. DOI:10.1088/1748-9326/7/4/045706 |
| [8] |
Ponce-Campos G E, Moran M S, Huete A, Zhang Y G, Bresloff C, Huxman T E, Eamus D, Bosch D D, Buda A R, Gunter S A, Scalley T H, Kitchen S G, McClaran M P, McNab W H, Montoya D S, Morgan J A, Peters D P C, Sadler E J, Seyfried M S, Starks P J. Ecosystem resilience despite large-scale altered hydroclimatic conditions. Nature, 2013, 494(7437): 349-352. DOI:10.1038/nature11836 |
| [9] |
Dong C Y, MacDonald G M, Willis K, Gillespie T W, Okin G S, Williams A P. Vegetation Responses to 2012-2016 Drought in Northern and Southern California. Geophysical Research Letters, 2019, 46(7): 3810-3821. DOI:10.1029/2019GL082137 |
| [10] |
Jiang W X, Wang L C, Feng L, Zhang M, Yao R. Drought characteristics and its impact on changes in surface vegetation from 1981 to 2015 in the Yangtze River Basin, China. International Journal of Climatology, 2020, 40(7): 3380-3397. DOI:10.1002/joc.6403 |
| [11] |
Huang K, Xia J Y. High ecosystem stability of evergreen broadleaf forests under severe droughts. Global Change Biology, 2019, 25(10): 3494-3503. DOI:10.1111/gcb.14748 |
| [12] |
Zhao M S, Running S W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science, 2010, 329(5994): 940-943. DOI:10.1126/science.1192666 |
| [13] |
Fang W, Huang S Z, Huang Q, Huang G H, Wang H, Leng G Y, Wang L, Guo Y. Probabilistic assessment of remote sensing-based terrestrial vegetation vulnerability to drought stress of the Loess Plateau in China. Remote Sensing of Environment, 2019, 232: 111290. DOI:10.1016/j.rse.2019.111290 |
| [14] |
Palmer W C. Meteorological Drought. Washington DC, USA: US Department of Commerce, Weather Bureau, 1965, 58-58. |
| [15] |
Belayneh A, Adamowski J, Khalil B. Short-term SPI drought forecasting in the Awash River Basin in Ethiopia using wavelet transforms and machine learning methods. Sustainable Water Resources Management, 2016, 2(1): 87-101. DOI:10.1007/s40899-015-0040-5 |
| [16] |
Manatsa D, Mushore T, Lenouo A. Improved predictability of droughts over southern Africa using the standardized precipitation evapotranspiration index and ENSO. Theoretical and Applied Climatology, 2017, 127(1/2): 259-274. |
| [17] |
张永, 陈发虎, 勾晓华, 靳立亚, 田沁花, 王有生, 彭剑峰. 中国西北地区季节间干湿变化的时空分布——基于PDSI数据. 地理学报, 2007, 62(11): 1142-1152. |
| [18] |
Wang M, Gu Q X, Jia X J, Ge J W. An assessment of the impact of Pacific Decadal Oscillation on autumn droughts in North China based on the Palmer drought severity index. International Journal of Climatology, 2019, 39(14): 5338-5350. DOI:10.1002/joc.6158 |
| [19] |
Yan H, Wang S Q, Lu H Q, Yu Q, Zhu Z C, Myneni R B, Liu Q, Shugart H H. Development of a remotely sensing seasonal vegetation-based Palmer Drought Severity Index and its application of global drought monitoring over 1982-2011. Journal of Geophysical Research, 2014, 119(15): 9419-9440. |
| [20] |
Zhong Z Q, He B, Guo L L, Zhang Y F. Performance of various forms of the palmer drought severity Index in China from 1961 to 2013. Journal of Hydrometeorology, 2019, 20(9): 1867-1885. DOI:10.1175/JHM-D-18-0247.1 |
| [21] |
Zhang L, Xiao J F, Zhou Y, Zheng Y, Li J, Xiao H. Drought events and their effects on vegetation productivity in China. Ecosphere, 2016, 7(12): e01591. |
| [22] |
Boisier J P, Ciais P, Ducharne A, Guimberteau M. Projected strengthening of Amazonian dry season by constrained climate model simulations. Nature Climate Change, 2015, 5(7): 656-660. DOI:10.1038/nclimate2658 |
| [23] |
Fu R, Yin L, Li W H, Arias P A, Dickinson R E, Huang L, Chakraborty S, Fernandes K, Liebmann B, Fisher R, Myneni R B. Increased dry-season length over southern Amazonia in recent decades and its implication for future climate projection. Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(45): 18110-18115. DOI:10.1073/pnas.1302584110 |
| [24] |
Xu X T, Medvigy D, Powers J S, Becknell J M, Guan K Y. Diversity in plant hydraulic traits explains seasonal and inter-annual variations of vegetation dynamics in seasonally dry tropical forests. New Phytologist, 2016, 212(1): 80-95. DOI:10.1111/nph.14009 |
| [25] |
Krings T. Hydropower, rice farmers and the state: the case of deforestation in Laos//Coping with changing environments. Routledge, 2018, 75-96. |
| [26] |
Manh N V, Dung N V, Hung N N, Kummu M, Merz B, Apel H. Future sediment dynamics in the Mekong Delta floodplains: impacts of hydropower development, climate change and sea level rise. Global and Planetary Change, 2015, 127: 22-33. DOI:10.1016/j.gloplacha.2015.01.001 |
| [27] |
Wu F F, Wang X, Cai Y P, Yang Z F, Li C H. Spatiotemporal analysis of temperature-variation patterns under climate change in the upper reach of Mekong River basin. Science of the Total Environment, 2012, 427-428: 208-218. DOI:10.1016/j.scitotenv.2012.03.081 |
| [28] |
van Ruijven J, Berendse F. Diversity enhances community recovery, but not resistance, after drought. Journal of Ecology, 2010, 98(1): 81-86. DOI:10.1111/j.1365-2745.2009.01603.x |
| [29] |
Ivits E, Horion S, Erhard M, Fensholt R. Assessing European ecosystem stability to drought in the vegetation growing season. Global Ecology and Biogeography, 2016, 25(9): 1131-1143. DOI:10.1111/geb.12472 |
| [30] |
Hu Z M, Guo Q, Li S G, Piao S L, Knapp A K, Ciais P, Li X R, Yu G R. Shifts in the dynamics of productivity signal ecosystem state transitions at the biome-scale. Ecology Letters, 2018, 21(10): 1457-1466. DOI:10.1111/ele.13126 |
| [31] |
Son N T, Chen C F, Chen C R, Chang L Y, Minh V Q. Monitoring agricultural drought in the Lower Mekong Basin using MODIS NDVI and land surface temperature data. International Journal of Applied Earth Observation and Geoinformation, 2012, 18: 417-427. DOI:10.1016/j.jag.2012.03.014 |
| [32] |
Zhou Q W, Luo Y, Zhou X, Cai M Y, Zhao C W. Response of vegetation to water balance conditions at different time scales across the karst area of southwestern China: A remote sensing approach. Science of the Total Environment, 2018, 645: 460-470. DOI:10.1016/j.scitotenv.2018.07.148 |
| [33] |
Mohammat A, Wang X H, Xu X T, Peng L Q, Yang Y, Zhang X P, Myneni R B, Piao S L. Drought and spring cooling induced recent decrease in vegetation growth in Inner Asia. Agricultural and Forest Meteorology, 2013, 178-179: 21-30. DOI:10.1016/j.agrformet.2012.09.014 |
| [34] |
Huete A, Didan K, Miura T, Rodriguez E P, Gao X, Ferreira L G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sensing of Environment, 2002, 83(1/2): 195-213. |
| [35] |
Adole T, Dash J, Atkinson P M. Characterising the land surface phenology of Africa using 500 m MODIS EVI. Applied Geography, 2018, 90: 187-199. DOI:10.1016/j.apgeog.2017.12.006 |
| [36] |
何奕萱, 易桂花, 张廷斌, 李景吉, 别小娟, 闾利, 郑飞鸽. 红河流域"通道-阻隔"作用下2000-2014年植被EVI变化趋势与驱动力. 生态学报, 2018, 38(6): 2056-2064. |
| [37] |
Yao R, Wang L C, Huang X, Gong W, Xia X G. Greening in rural areas increases the surface urban heat island intensity. Geophysical Research Letters, 2019, 46(4): 2204-2212. DOI:10.1029/2018GL081816 |
| [38] |
Abatzoglou J T, Dobrowski S Z, Parks S A, Hegewisch K C. TerraClimate, a high-resolution global dataset of monthly climate and climatic water balance from 1958-2015. Scientific Data, 2018, 5: 170191. DOI:10.1038/sdata.2017.191 |
| [39] |
张宝庆. 黄土高原干旱时空变异及雨水资源化潜力研究[D]. 杨凌: 西北农林科技大学, 2014.
|
| [40] |
De Keersmaecker W, Lhermitte S, Tits L, Honnay O, Somers B, Coppin P. A model quantifying global vegetation resistance and resilience to short-term climate anomalies and their relationship with vegetation cover. Global Ecology and Biogeography, 2015, 24(5): 539-548. DOI:10.1111/geb.12279 |
| [41] |
Jha S, Das J, Sharma A, Hazra B, Goyal M K. Probabilistic evaluation of vegetation drought likelihood and its implications to resilience across India. Global and Planetary Change, 2019, 176: 23-35. DOI:10.1016/j.gloplacha.2019.01.014 |
| [42] |
Guo H, Bao A M, Liu T, Ndayisaba F, He D M, Kurban A, De Maeyer P. Meteorological drought analysis in the Lower Mekong Basin using satellite-based long-term CHIRPS product. Sustainability, 2017, 9(6): 901. DOI:10.3390/su9060901 |
| [43] |
Mitchard E T A. The tropical forest carbon cycle and climate change. Nature, 2018, 559(7715): 527-534. DOI:10.1038/s41586-018-0300-2 |
| [44] |
Hoffman A M, Smith M D. Gene expression differs in codominant prairie grasses under drought. Molecular Ecology Resources, 2018, 18(2): 334-346. DOI:10.1111/1755-0998.12733 |
| [45] |
Tang H, Dubayah R. Light-driven growth in Amazon evergreen forests explained by seasonal variations of vertical canopy structure. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(10): 2640-2644. DOI:10.1073/pnas.1616943114 |
| [46] |
McDowell N, Pockman W T, Allen C D, Breshears D D, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams D G, Yepez E A. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?. New Phytologist, 2008, 178(4): 719-739. DOI:10.1111/j.1469-8137.2008.02436.x |
| [47] |
Huang K, Xia J Y, Wang Y P, Ahlström A, Chen J Q, Cook R B, Cui E Q, Fang Y Y, Fisher J B, Huntzinger D N, Li Z, Michalak A M, Qiao Y, Schaefer K, Schwalm C, Wang J, Wei Y X, Xu X N, Yan L M, Bian C Y, Luo Y Q. Enhanced peak growth of global vegetation and its key mechanisms. Nature Ecology & Evolution, 2018, 2(12): 1897-1905. |
| [48] |
Wen Y Y, Liu X P, Yang J, Lin K, Du G M. NDVI indicated inter-seasonal non-uniform time-lag responses of terrestrial vegetation growth to daily maximum and minimum temperature. Global and Planetary Change, 2019, 177: 27-38. DOI:10.1016/j.gloplacha.2019.03.010 |