 2021, Vol. 41
2021, Vol. 41文章信息
- 蒋裕良, 蒙涛, 许为斌, 谭卫宁, 张强, 刘演
- JIANG Yuliang, MENG Tao, XU Weibin, TAN Weining, ZHANG Qiang, LIU Yan
- 喀斯特山顶区域濒危裸子植物群落特征及其地形关联
- Community characteristics of endangered Gymnosperms and their topographic relationships in a karst mountaintop region
- 生态学报. 2021, 41(5): 1868-1877
- Acta Ecologica Sinica. 2021, 41(5): 1868-1877
- http://dx.doi.org/10.5846/stxb201904020635
-
文章历史
- 收稿日期: 2019-04-02
- 网络出版日期: 2021-01-05





2. 广西木论国家级自然保护区管理中心, 环江 547199
2. Administration of Mulun National Nature Reserve of Guangxi, Huanjiang 547199, China
喀斯特森林植被是一种地域性植被, 它的区系组成、外貌结构、生产力和生物量等有很强的特殊性[1-3]。喀斯特森林植被以裸子植物群落最为少见, 群落类型极少, 主要分布在亚热带以及北热带海拔大于700 m的狭窄区域[4]。喀斯特森林裸子植物树种也比较稀少, 云南石林地区记录了10种, 主要为松科和柏科树种, 占云南裸子植物树种的11.0%[5-6];贵州茂兰喀斯特森林记录12种[5];广西弄岗喀斯特森林仅记录3种[7];广西木论喀斯特森林记录11种, 占广西裸子植物树种的23.6%, 但除杉木(Cunninghamia lanceolata)以外, 其余均为濒危物种[8]。越南黄金柏(Xanthocyparis vietnamensis)是2012年在广西木论喀斯特森林发现的中国新纪录, 该物种最早发现于越南北部河江省(Ha Giang Province)广薄区(Quan Ba District)的喀斯特山地, 于2002年被认定为柏科新属新种[9], 2004年被IUCN列入极危物种(CR), 2012年被改列濒危物种(EN)[10-11]。根据野外调查(数据截至2019年7月), 木论喀斯特森林越南黄金柏已知个体数仅有28株。包括越南黄金柏在内, 喀斯特濒危裸子植物常常零散分布在山顶或者中上坡部位, 地势陡峭、生境严苛, 再加上种群较小等原因, 较难开展常规的种群和群落调查, 因此缺乏足够的研究报道。有研究指出, 华南五针松(Pinus kwangtungensis)[12]、岩生翠柏(Calocedrus rupestris)[13-15]和短叶黄杉(Pseudotsuga brevifolia)在喀斯特山顶地段有良好的适应性, 其种群能够长期稳定地存在[16]。
地形是影响山地森林植物分布的主要非地带性环境因子。地形因子直接影响土层厚度、太阳辐射与降水的空间再分配, 进而改变土壤、光照、温度和湿度等非生物因子, 对植物物种组成和生物量的空间分布产生重要的影响[17-20]。地形因素结合其他因素的作用, 可以解释局域群落的物种组成和分布特征以及濒危植物丰富度的变异[21]。喀斯特森林地形地貌特殊, 小生境复杂多样, 非生物因子具有高度的异质性[22]。张忠华等[23]、黄甫昭等[24]和Du等[25-26]研究均表明喀斯特森林地形空间异质性对物种组成和分布格局具有显著性影响。所以, 了解喀斯特森林濒危裸子植物分布与地形的关联, 很可能是发现它们生存潜力、维持机制及受威胁因素的关键。
自然界中的物种并不是独立存在的, 必须与其他物种共存于群落中。杨永等[27]从实际保护案例中发现, 裸子植物濒危的一个重要现象可能是由于竞争力不够, 在被子植物占优势的群落中处于较弱的竞争态势, 从而导致物种在自然演替过程中被逐渐淘汰而濒危。群落物种之间的谱系关系能够体现物种生态位的相似性, 群落谱系结构能够反映局域尺度上物种共存的生态过程[28-29]。群落谱系结构分析与群落生态学研究的结合, 可以为濒危裸子植物生存现状的评估提供新的角度。为此, 本文以木论喀斯特山顶区域典型的濒危裸子植物群落为研究对象, 通过分析群落生物量、物种多样性、谱系结构等的空间变异及其与地形的相关性, 了解群落特征的变化规律, 以期深化对喀斯特濒危裸子植物群落中物种共存机制的理解, 为濒危物种的保护和恢复提供理论参考。
1 方法 1.1 研究区域概况广西木论国家级自然保护区(107°54′01″—108°05′51″E, 25°06′09″—25°12′25″N)位于广西环江毛南族自治县西北部, 地处云贵高原南缘, 北与贵州茂兰国家级自然保护区相交, 区境呈西南至东北向, 东西长19.80 km, 南北宽10.75 km, 林区总面积89.69 km2。总体地势西北高、东南低, 地形地貌由典型的喀斯特峰丛洼地、谷地和峰林洼地、谷地及洞穴等构成, 以锥状峰丛峰林为主体, 锥峰海拔高度约500—1000 m, 相对高度约100—400 m, 锥峰形体整齐、两坡对称、平均约45°。该区属于中亚热带湿润季风气候, 年均日照1451 h, 年均气温16.9℃, 年均降水量1675 mm。保护区以喀斯特森林生态系统为重点保护对象, 原生林保存较为完好, 森林植被为中亚热带喀斯特常绿阔叶落叶混交林。区内有维管束植物207科682属1446种, 其中蕨类植物35科66属192种, 裸子植物7科12属14种(包含栽培种), 被子植物165科604属1240种;珍稀濒危植物种类丰富, 共有98种, 其中国家重点保护野生植物(第一批)17种, 广西壮族自治区重点保护野生植物(第一批)76种[8]。
1.2 样方设置与调查木论保护区内濒危裸子植物多为零散分布, 其典型群落比较少见。基于广泛的实地踏查, 选出裸子植物个体数量相对较多且分布较为集中的典型群落开展调查。该群落位于海拔约820 m的近山顶部位(107°58′04″E, 25°06′54″N), 有濒危裸子植物华南五针松、越南黄金柏和岩生翠柏。由于山顶区域面积狭小、地形崎岖险峻, 不易于设置大面积样方。综合考虑, 以越南黄金柏植株为中心点, 设置12个大小为100 m2的样方, 样方东西(或南北)边长10 m, 样方间存在部分重叠;全部样方基本将40 m×20 m范围内的所有濒危裸子植物个体包含在内(图 1)。对样方内基径大于1 cm的木本植物进行每木调查, 记录种类、基径(离地面1—10 cm处测量)、坐标等数据;对于竹类植物, 记录其种类、丛数、每丛竿数、竹竿基径和坐标。

|
| 图 1 濒危裸子植物个体分布和样方设置示意图 Fig. 1 Sketch for individual distributions of endangered Gymnosperms and sampling plots 图中样方的北边是危崖, 南边是山顶, 调查范围的相对高差约20 m |
测量和计算样方的相对海拔、坡向、坡度和凹凸度等地形因子指标。选择某个样方的中心点作为参考点, 其他样方中心点与该点的高差记为相应样方的相对海拔。两点间的高差测量使用森林罗盘仪等。凹凸度、坡度和坡向的测定方法如下:
凹凸度:样方中心与样方4个边线中点的高差的平均值。当样方中点的高度大于边线4个中点时, 该样方凹凸度为正值, 即正地形, 反之为负地形。
坡度:从样方4个边线中点取3个点组成一个平面, 4个点可以组合成4个不同的平面, 这4个平面与水平面夹角度数的平均值为该样方的坡度值。
坡向(A):定正北为0°, 顺时针方向至360°仍为正北;先计算
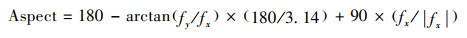
|
式中fx为样方西边线中点与东边线中点的高差, fy为样方北边线中点与南边线中点的高差。当Aspect ≤180时, A = Aspect;当Aspect > 180时, A = 360-Aspect。
1.3 群落特征分析 1.3.1 生物量取样方所有物种的个体数(N)和总基面积(BA)来表征群落生物量, 用相对基面积(RBAj)来表征物种(或类群) j的相对生物量, BA和RBAj计算方法如下:
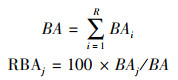
|
式中R为物种丰富度, 即样方内的物种数;BAi为第i个物种的基面积, BAj为物种(或类群)j的基面积。
1.3.2 物种多样性采用物种丰富度(R)、Shannon多样性指数(H)、Simpson多样性指数(D)和Pielou均匀度指数(J)分析群落物种多样性特征[30], 其中H、D和J的计算公式为:
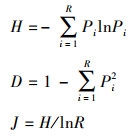
|
式中Pi= Ni / N, Ni为第i个物种的个体数, N为个体总数。
1.3.3 双子叶植物谱系关系利用在线软件phylomatic[31](http://phylodiversity.net/phylomatic/)构建谱系树, 将群落调查所得的双子叶植物名录及其科属信息输入phylomatic (Version 3)中, 自动生成基于被子植物分类系统APGIII且具有进化枝长度的谱系结构树(phylomatic tree, version:Zanne 2014)[32]。
选择净关系指数(NRI, net relatedness index)和最近关系指数(NTI, nearest taxon index)来分析双子叶植物的谱系结构。NRI和NTI的计算公式如下[33-34]:

|
式中MPD是样方内所有物种对的平均谱系距离的实际观测值, rndMPDT是谱系库里零模型中的平均成对谱系距离的期望值, sdrndMPD是rndMPDT的标准偏差;MNPD是样方内所有物种与亲缘关系最近物种的平均谱系距离的实际观测值, rndMNPDT是零模型中的最近谱系距离的期望值, sdrndMNPD是它的标准差(本文采用随机化谱系树末端分支方案, n=999)。NRI或NTI显著大于0时, 意味着样方内物种的谱系关系比群落谱系库的随机化情形更紧密, 表现为谱系聚集, 反之为谱系发散[28]。NRI和NTI的计算在Phylocom中完成。
1.4 数据分析 1.4.1 半变异函数分析采用半变异函数定量分析群落特征的空间变异性, 该函数的通式为[35]:
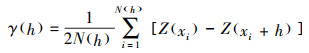
|
式中, γ(h)为半变异函数值;h为样方的间隔距离(中心点距离), 其小于最大间隔距离;N(h)为间隔距离为h时的样方对数;Z(xi)和Z(xi+h)分别为变量Z在空间位置xi和xi+h处的观测值。
以γ(h)为纵轴, h为横轴绘制半变异函数散点图, 用线性模型、球状模型、指数模型和高斯模型分别进行拟合, 并由决定系数(R2)判定最优拟合模型。理论模型通过块金值(C0, nugget)、基台值(C0+C1, sill)、块金系数[C0/(C0+C1)]和变程(range)等参数来描述变量的空间变化规律[36]。块金值表示由随机因素引起的空间变异, 基台值为随机性变异和结构性变异的总和, 反映总的空间变异程度;两者值越大表明异质性程度越高, 但它们受自身因素和测量单位的影响较大, 不能用于不同变量间的比较。块金系数表示随机空间变异占总空间变异的程度, 其值越低表明变量的空间相关性越强。一般根据块金系数大小, 可将变量的空间自相关程度分为较强(≤25%)、中等(25%—75%)、较弱(≥75%)[37]。变程表示变量存在空间自相关性的距离上限, 变程越小, 说明变量的空间连续性越差。上述分析在GS+9.0软件中完成。
1.4.2 描述性统计与相关分析对群落特征数据进行描述性统计分析, 根据变异系数绝对值大小, 各指标的变异性可分为弱变异(≤10%)、中变异(10%—100%)、强变异(≥100%)[38]。采用Pearson相关系数检验群落特征与相对海拔、坡向、坡度和凹凸度的相关性。以上分析在SPSS17.0软件中完成。
2 结果与分析 2.1 群落物种组成调查记录到植物73种, 隶属于39科58属;裸子植物2科3属3种, 双子叶植物35科52属67种, 单子叶植物2科3属3种。物种数最多的科为壳斗科(7种), 其次是蔷薇科(6种)、樟科(5种)和鼠李科(5种);物种数最多的属是青冈属(Cyclobalanopsis)、石楠属(Photinia)、樟属(Cinnamomum)和榕属(Ficus), 各有3种;64.1%的科和81.0%的属只有1个物种。
本研究群落为常绿落叶针阔混交林, 主要乔木树种有圆果化香树(Platycarya longipes)、青冈(Cyclobalanopsis glauca)、密花树(Myrsine seguinii)、罗城鹅耳枥(Carpinus luochengensis)、乌冈栎(Quercus phillyreoides)、异叶花椒(Zanthoxylum dimorphophyllum )、越南黄金柏、华南五针松等等。乔木下层有单子叶植物芸香竹(Bonia amplexicaulis)、贵州悬竹(Ampelocalamus calcareus)和石山棕(Guihaia argyrata), 石山棕仅见1株。
2.2 群落特征的描述性统计调查共记录个体1499株, 平均基径约3.2 cm;濒危裸子植物共计55株, 越南黄金柏、华南五针松、岩生翠柏分别有17、12、26株。群落为旱生类型, 树木普遍比较矮小, 高度多为4—6 m;群落上层有越南黄金柏和华南五针松的大树零散分布, 个别大树高度约8 m;群落下层常见芸香竹和贵州悬竹, 分别有133丛和8丛(株)。
群落特征的描述性统计结果见表 1, 样方个体数为152—271株, 总基面积为3890—5445 cm2;濒危裸子植物相对基面积(RBAGy)为3.12%—50.19%, 平均值为24.35%, 说明濒危裸子植物在群落中占据一定的优势。由于样方设置服从越南黄金柏成年个体的分布, 越南黄金柏相对基面积(RBA1)区别于华南五针松(RBA2)和岩生翠柏(RBA3)的相对基面积, RBA1的平均值较大且偏度和峰度均较小。
| 群落特征 Community characteristic |
最大值 Maximum |
最小值 Minimum |
平均值 Mean |
标准差 Standard deviation |
偏度 Skewness |
峰度 Kurtosis |
变异系数 Coefficient of variation/% |
K-S检验 K-S test (P value) |
| 个体数 Individual number | 271 | 152 | 221.50 | 37.16 | -0.49 | -0.52 | 16.8 | 0.930 |
| 总基面积 Total basal area of all species | 5445 cm2 | 3890 cm2 | 4526 cm2 | 452.03 | 0.29 | 0.02 | 10.0 | 0.773 |
| 濒危裸子植物相对基面积 Relative basal area of endangered Gymnosperms |
50.19% | 3.12% | 24.35% | 0.12 | 0.10 | 1.35 | 50.5 | 0.834 |
| 越南黄金柏相对基面积 Relative basal area of Xanthocyparis vietnamensis |
28.89% | 0.51% | 15.76% | 0.12 | -0.31 | -2.11 | 77.3 | 0.419 |
| 华南五针松相对基面积 Relative basal area of Pinus kwangtungensis |
30.12% | 0.00% | 7.13% | 0.11 | 1.26 | 0.16 | 155.6 | 0.043 |
| 岩生翠柏相对基面积 Relative basal area of Calocedrus rupestris |
4.80% | 0.00% | 1.46% | 0.02 | 1.49 | 1.16 | 106.8 | 0.085 |
| 物种丰富度 Species richness | 44 | 33 | 39.75 | 3.31 | -1.17 | 0.70 | 8.3 | 0.546 |
| 香农多样性指数 Shannon′s diversity index | 3.36 | 2.63 | 3.23 | 0.20 | -2.75 | 8.13 | 6.2 | 0.184 |
| 辛普森多样性指数 Simpson′s diversity index | 0.96 | 0.84 | 0.94 | 0.03 | -3.24 | 10.80 | 3.4 | 0.080 |
| 均匀度指数 Pielou′s evenness index | 0.91 | 0.71 | 0.87 | 0.06 | -2.75 | 8.13 | 6.4 | 0.184 |
| 双子叶植物净关系指数 Net relatedness index of Dicotyledon |
1.86 | -2.23 | 1.04 | 1.19 | -2.23 | 5.32 | 114.5 | 0.266 |
| 双子叶植物最近关系指数 Nearest taxon index of Dicotyledon |
0.93 | -2.17 | -0.31 | 0.84 | -0.94 | 1.04 | -268.1 | 0.883 |
总基面积、物种丰富度、Shannon多样性指数、Simpson多样性指数和Pielou均匀度指数为弱变异性;个体数、RBAGy和RBA1为中变异性;RBA2、RBA3、双子叶植物净关系指数(NRI)和最近关系指数(NTI)为强变异性。根据K-S检验结果, 除RBA2外, 各指标均服从正态分布(P > 0.05)。
12个样方中仅有1个样方的NRI显著小于0, 其余处于随机置信区间内(绝对值小于1.96), 多数(比例为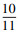

由表 2可知, 物种丰富度的最佳拟合模型为线性模型, 其他指标均为高斯模型。除个体数的模型拟合度较低(0.441)外, 其余指标的模型拟合度均较高(0.736—0.986), 说明半变异函数理论模型总体上能较好地反映群落特征的空间变化。
| 群落特征 Community characteristic |
模型 Model |
块金值 Nugget (C0) |
基台值 Sill (C0+C1) |
块金系数 C0/(C0+C1) |
变程 Range /m |
拟合度 R2 |
| 个体数 Individual number | 高斯模型 | 1 | 1388 | 0.001 | 5.66 | 0.441 |
| 总基面积 Total basal area of all species | 高斯模型 | 100 | 263300 | 0.000 | 4.75 | 0.795 |
| 濒危裸子植物相对基面积 Relative basal area of endangered Gymnosperms |
高斯模型 | 0.1 | 210.7 | 0.000 | 8.23 | 0.736 |
| 越南黄金柏相对基面积 Relative basal area of Xanthocyparis vietnamensis |
高斯模型 | 14.8 | 231.4 | 0.064 | 11.86 | 0.986 |
| 华南五针松相对基面积 Relative basal area of Pinus kwangtungensis |
高斯模型 | 1 | 220 | 0.005 | 16.78 | 0.912 |
| 岩生翠柏相对基面积 Relative basal area of Calocedrus rupestris |
高斯模型 | 0.01 | 3.61 | 0.003 | 14.15 | 0.865 |
| 物种丰富度 Species richness | 线性模型 | 1.3 | 18.6 | 0.070 | 16.69 | 0.786 |
| 香农多样性指数 Shannon′s diversity index | 高斯模型 | 0.001 | 0.660 | 0.002 | 71.01 | 0.842 |
| 辛普森多样性指数 Simpson′s diversity index | 高斯模型 | 0.00001 | 0.01745 | 0.001 | 71.01 | 0.849 |
| 均匀度指数 Pielou′s evenness index | 高斯模型 | 0.00001 | 0.02002 | 0.000 | 42.44 | 0.823 |
| 双子叶植物净关系指数 Net relatedness index of Dicotyledon |
高斯模型 | 0.01 | 4.03 | 0.002 | 28.49 | 0.966 |
| 双子叶植物最近关系指数 Nearest taxon index of Dicotyledon |
高斯模型 | 0.001 | 0.51 | 0.002 | 6.51 | 0.755 |
受自身因素和测量单位的影响, 不同指标的块金值和基台值差异较大;在同样表征物种相对生物量的指标RBA1、RBA2和RBA3中, 块金值和基台值以RBA1最大, RBA3最小。各指标的块金系数均小于25%, 说明它们均有较强的空间相关性。
个体数和总基面积变程较小, 分别为5.66 m和4.75 m;相对基面积的变程为8.23—16.78 m, 以RBAGy最小, RBA2最大;多样性指数的变程为16.69—71.01 m, 以Shannon多样性指数和Simpson多样性指数最大, 物种丰富度最小;双子叶植物谱系结构指数NRI和NTI的变程分别为28.49 m和6.51 m。
2.4 群落特征与地形因子的相关性由表 3可知, 总基面积与凹凸度存在显著的正相关关系(P < 0.05);濒危裸子植物相对基面积与坡度存在显著的正相关关系(P < 0.05);岩生翠柏相对基面积与相对海拔存在显著的负相关关系(P < 0.05);双子叶植物最近关系指数与相对海拔存在极显著的正相关关系(P < 0.01), 与坡向和坡度存在显著的负相关关系(P < 0.05)。
| 群落特征 Community characteristic |
地形因子 Topographic factor | |||
| 相对海拔 Relative elevation |
坡向 Slope aspect |
坡度 Slope degree |
凹凸度 Convexity |
|
| 个体数 Individual number | 0.046 | 0.071 | 0.265 | -0.010 |
| 总基面积 Total basal area of all species | -0.114 | -0.038 | 0.447 | 0.705* |
| 濒危裸子植物相对基面积 Relative basal area of endangered Gymnosperms |
-0.166 | -0.334 | 0.674* | 0.525 |
| 越南黄金柏相对基面积 Relative basal area of Xanthocyparis vietnamensis |
0.322 | -0.194 | 0.206 | 0.257 |
| 华南五针松相对基面积 Relative basal area of Pinus kwangtungensis |
-0.440 | -0.127 | 0.453 | 0.261 |
| 岩生翠柏相对基面积 Relative basal area of Calocedrus rupestris |
-0.693* | -0.217 | 0.478 | 0.279 |
| 物种丰富度 Species richness | 0.230 | -0.087 | -0.379 | 0.049 |
| 香农多样性指数 Shannon′s diversity index | -0.130 | -0.098 | -0.091 | -0.067 |
| 辛普森多样性指数 Simpson′s diversity index | -0.155 | -0.023 | -0.056 | -0.109 |
| 均匀度指数 Pielou′s evenness index | -0.130 | -0.098 | -0.091 | -0.067 |
| 双子叶植物净关系指数 Net relatedness index of Dicotyledon |
0.131 | -0.046 | -0.241 | 0.080 |
| 双子叶植物最近关系指数 Nearest taxon index of Dicotyledon |
0.749** | -0.606* | -0.656* | -0.177 |
| *: P < 0.05; **: P < 0.01. | ||||
喀斯特森林群落的立地条件特殊, 基岩化学特性导致其土层浅薄和干旱, 植物易受到干旱胁迫的影响, 故而生产力和生物量普遍低于相同气候带的非喀斯特森林[2, 39]。喀斯特山顶区域干旱最为突出, 生境苛刻, 树木明显矮化、生长缓慢, 以小径级居多。群落个体平均基径约3.2 cm, 样方平均个体密度约2.2株/m2, 与海南岛热带云雾林群落(平均胸径4.6—6.4 cm, 个体密度101.6—185.0株/hm2)[40]和茂兰喀斯特灌木群落、次顶级和顶级森林群落(平均胸径3.9—9.6 cm, 个体密度1956—5600株/hm2)[41]相比, 其个体密度大、树木径级小的特征比较突出。群落中残存着濒危裸子植物越南黄金柏、华南五针松和岩生翠柏, 它们少数个体形体高大(最大基径约42 cm), 是群落生物量的重要组成。个体数和总基面积均表现为强烈的空间自相关性, 说明它们的空间变异主要由非随机性因素(比如地形、土壤养分和水分等)引起;但它们的空间自相关范围较小, 仅在6 m以内, 这体现了喀斯特濒危裸子植物群落结构的复杂性。
本研究中群落生物量分布与地形特征密切相关, 表现出总基面积与凹凸度呈显著的正相关关系, 即总基面积的较大值往往出现在明显凸起的正地形部位;此外, 濒危裸子植物的相对基面积随着坡度的增大而增加, 表明较大的坡度对濒危裸子植物的生长和生物量积累起到了积极的作用。地形的变化控制着光、温、水和土壤养分等资源因子的空间再分配而成为植被生物量分布格局的重要影响因子[19-20]。悬崖生态学相关研究表明, 悬崖生境光、温和水条件并不十分严苛, 变化相对温和, 比较有利于稀有物种的生长, 其上的植被往往独特于周围的植被, 常保存有一些古老的森林群落[42-43], 与本研究的结果符合。木论喀斯特森林处于亚热带季风气候区, 雨热同期、雨量丰富, 以致岩石溶蚀和土壤侵蚀的强度较大[44]。有研究表明, 凹凸度较大的地形更易受到侵蚀, 土壤水分和营养元素流失而不利于植物生长[45], 该结果与本研究的结果存在矛盾, 不支持总基面积与凹凸度的正相关性。长期的喀斯特作用使得水、土、岩和植物联为一体, 相互依赖、相互影响。因此我们认为喀斯特植被可以改变地形, 一般来说, 树木稀疏的地方地表侵蚀量较大, 而树木繁茂的地方地表侵蚀量较小[46];山顶区域的群落生物量空间连续性较差, 非均质性较强, 且树木生长缓慢, 从较长的时空尺度上看, 群落生物量的空间异质性可能对地形凹凸度起到塑造作用, 树木繁茂、生物量高的部位可能会逐渐形成正地形。
3.2 物种多样性特征本研究群落物种分属39科58属73种, 种类丰富, 但科(属)内的物种组成比较简单, 大多数科(属)内仅有1个物种;壳斗科(青冈属)、蔷薇科(石楠属)和樟科(樟属)等少数科(属)的科(属)内物种数较多, 表现出相对较高的分化和演化程度。10 m×10 m样方的物种数为33—44种, 平均为39.8种, 略高于邻近区域的茂兰喀斯特森林坡地和槽谷的群落样方(约37种)[47]。上述结果表明, 木论喀斯特山顶区域的植物群落物种丰富多样、组织水平较高。该区的生态条件可能具有较强的复杂性和特殊性, 有利于众多科属共存, 但不利于同科属内的多物种共存。
群落物种多样性的4个指标均为弱变异性, 与地形因子无显著的相关关系。半变异函数理论模型较好地反映了物种多样性的空间分布特征, Shannon多样性指数、Simpson多样性指数和Pielou均匀度指数是物种数量和物种个体数量特征的综合反映, 它们分别在71.01 m、71.01 m、42.44 m范围内呈现强烈的空间自相关, 空间连续性较好;而物种丰富度仅反映了物种数量的高低水平, 其在16.69 m范围内呈现强烈的空间自相关, 空间连续性相对较差;随机因素对这4个指标空间分布的贡献较小, 它们的变异主要由非随机性因素(比如土壤基质、气候、植被本身特性等)引起。对于喀斯特山顶区域群落物种多样性的研究, 应适当增加调查样方的间距, 减小空间自相关的影响, 以便准确地理解、掌握和预测该区群落物种多样性的变化规律。
3.3 双子叶植物谱系结构特征与地形因子的关系濒危裸子植物在群落中的相对基面积为23.0%, 个体数占比仅有3.7%;单子叶植物处于群落的下层空间, 竞争力相对有限;从物种数、个体数和生物量来看, 双子叶植物在群落中均占有绝对的优势。由于裸子植物和单子叶植物的存在对群落谱系结构有较大影响[48], 本研究只对双子叶植物进行谱系结构分析, 发现样方之间NRI和NTI有强变异性, 多个样方呈现随机格局, 仅个别样方表现为显著的发散格局。许多在其他地方开展的研究发现, 群落谱系结构与研究尺度(空间、物种库和分类群)相关, 尺度越小, 越可能为谱系发散;小空间尺度下生境资源通常相对有限、较为均质, 此时群落构建的关键因子是竞争排斥[33, 49-50]。喀斯特森林小生境类型丰富, 生境资源分布的空间异质性强烈, 较小的空间尺度也会存在较多环境变量[22]。因而我们认为, 生境过滤在喀斯特森林较小的空间尺度下对物种集聚也会起较大的作用, 100 m2的山顶群落表现出谱系结构的随机格局, 可能是生境过滤与竞争排斥双重作用的结果。
NRI和NTI均呈强烈的空间自相关性, 两者的空间自相关范围差距较大(分别为28.49 m和6.51 m), NRI的空间连续性较好, 这可能与指标本身的特性有关。NRI侧重于从整体上描述样方物种形成的谱系结构, 而NTI侧重最近邻体的种间亲缘关系[33]。对于探索小空间尺度上的群落谱系结构异质性, NTI会比NRI更敏感。NTI与地形因子密切相关, 表现出随着相对海拔的降低、坡度的增大以及坡向远离正北方向, 由随机向发散转变的趋势。这种趋势可能是群落内竞争排斥作用加强的体现。对于喀斯特山地来说, 一般相对海拔较低的区域水分条件相对较好(岩生翠柏相对基面积与相对海拔的负相关关系也暗示较低的相对海拔比较有利于它的生长), 坡度较大的生境光、温和水条件的变化相对温和(见前文讨论), 远离正北方向的坡面光温条件相对较好。通常在相对良好的生存条件下生境过滤作用相对较弱, 有利于更多物种共存, 由此促进了竞争排斥作用, 使得亲缘关系较近的物种共存概率降低, 谱系结构趋向发散[18]。
4 结论本研究群落的个体数、总基面积和NTI在较小的空间尺度上具有空间自相关性, 空间连续性较差;濒危裸子植物的相对基面积具有随坡度增大而增加的趋势;双子叶植物谱系结构呈现随机格局, NTI具有随相对海拔降低、坡度增大以及坡向远离正北方向而呈现由随机向发散转变的趋势。这说明了喀斯特濒危裸子植物群落结构比较复杂, 群落构建过程可能是生境过滤和竞争排斥共同作用的结果, 但随着相对海拔降低、坡度增大以及坡向远离正北方向, 竞争排斥作用可能会增强。上述结果有助于理解喀斯特濒危裸子植物对山顶部位的、地势陡峭且接近正北坡向的生境的依赖现象。
| [1] |
孙鸿烈. 中国生态系统. 北京: 科学出版社, 2005: 327-365.
|
| [2] |
于维莲, 董丹, 倪健. 中国西南山地喀斯特与非喀斯特森林的生物量与生产力比较. 亚热带资源与环境学报, 2010, 5(2): 25-30. DOI:10.3969/j.issn.1673-7105.2010.02.004 |
| [3] |
郭柯, 刘长成, 董鸣. 我国西南喀斯特植物生态适应性与石漠化治理. 植物生态学报, 2011, 35(10): 991-999. |
| [4] |
李先琨, 苏宗明, 吕仕洪, 欧祖兰, 向悟生, 区智, 陆树华. 广西岩溶植被自然分布规律及对岩溶生态恢复重建的意义. 山地学报, 2003, 21(2): 129-139. DOI:10.3969/j.issn.1008-2786.2003.02.001 |
| [5] |
徐远杰. 典型喀斯特地区木本植物区系比较研究[D]. 昆明: 西南林学院, 2007.
|
| [6] |
冯建孟, 朱有勇. 云南地区裸子植物的区系组成及物种多样性分布格局. 西南大学学报: 自然科学版, 2011, 33(10): 51-56. |
| [7] |
吴望辉. 广西弄岗国家级自然保护区植物区系地理学研究[D]. 桂林: 广西师范大学, 2011.
|
| [8] |
彭日成. 广西木论国家级自然保护区维管束植物区系研究[D]. 桂林: 广西师范大学, 2013.
|
| [9] |
Farjon A, Hiep N T, Harder D K, Loc P K, Averyanov L. A new genus and species in Cupressaceae (Coniferales) from northern Vietnam, Xanthocyparis vietnamensis. Novon, 2002, 12(2): 179-189. DOI:10.2307/3392953 |
| [10] |
蒙涛, 彭日成, 钟国芳, 游旨价, 谭卫宁, 许为斌. 黄金柏属——中国柏科一新记录属. 广西植物, 2013, 33(3): 388-391, 400. DOI:10.3969/j.issn.1000-3142.2013.03.017 |
| [11] |
Thomas P. Xanthocyparis vietnamensis. The IUCN Red List of Threatened Species 2013: e.T44028A2991576.[2019-02-19]. https://www.iucnredlist.org/species/44028/2991576.
|
| [12] |
杜道林, 刘玉成, 苏杰. 茂兰喀斯特山地广东松种群结构和动态初步研究. 植物生态学报, 1996, 20(2): 159-166. |
| [13] |
宁世江, 赵天林, 唐润琴, 覃文更, 梁其彪, 李瑞棠, 莫权辉, 谭卫宁. 木论喀斯特林区翠柏群落学特征的初步研究. 广西植物, 1997, 17(4): 321-330. |
| [14] |
宁世江, 赵天林, 李瑞棠, 唐润琴, 金代钧, 梁其彪, 覃文更, 谭卫宁. 木论喀斯特林区珍稀濒危植物资源的研究. 广西植物, 1998, 18(3): 247-255. |
| [15] |
农东新, 吴望辉, 蒋日红, 黄俞淞, 许为斌. 广西翠柏属(柏科)植物小志. 广西植物, 2011, 31(2): 155-159. DOI:10.3969/j.issn.1000-3142.2011.03.004 |
| [16] |
刘济明. 贵州茂兰喀斯特山地顶部森林群落种子库研究. 林业科学研究, 2000, 13(1): 44-50. DOI:10.3321/j.issn:1001-1498.2000.01.007 |
| [17] |
Liu J J, Tan Y H, Slik J W F. Topography related habitat associations of tree species traits, composition and diversity in a Chinese tropical forest. Forest Ecology and Management, 2014, 330: 75-81. DOI:10.1016/j.foreco.2014.06.045 |
| [18] |
赵鸣飞, 薛峰, 王宇航, 王国义, 邢开雄, 康慕谊, 王菁兰. 山西芦芽山针叶林草本层群落谱系结构与多样性的海拔格局. 植物生态学报, 2017, 41(7): 707-715. |
| [19] |
Homeier J, Breckle S W, Günter S, Rollenbeck R T, Leuschner C. Tree Diversity, forest structure and productivity along altitudinal and topographical gradients in a species-rich Ecuadorian montane rain forest. Biotropica, 2010, 42(2): 140-148. DOI:10.1111/j.1744-7429.2009.00547.x |
| [20] |
Lin D M, Lai J S, Muller-Landau H C, Mi X C, Ma K P. Correction: topographic variation in aboveground biomass in a subtropical evergreen broad-leaved forest in China. PLoS One, 2013, 8(5). DOI:10.1371/annotation/779db3d7-8d77-4c40-8f64-c694c2e30912 |
| [21] |
Liu J J, Slik J W F. Forest fragment spatial distribution matters for tropical tree conservation. Biological Conservation, 2014, 171: 99-106. DOI:10.1016/j.biocon.2014.01.004 |
| [22] |
刘方, 王世杰, 罗海波, 刘元生, 刘鸿雁. 喀斯特森林生态系统的小生境及其土壤异质性. 土壤学报, 2008, 45(6): 1055-1062. DOI:10.3321/j.issn:0564-3929.2008.06.007 |
| [23] |
张忠华, 胡刚, 祝介东, 倪健. 喀斯特森林土壤养分的空间异质性及其对树种分布的影响. 植物生态学报, 2011, 35(10): 1038-1049. |
| [24] |
黄甫昭, 丁涛, 李先琨, 郭屹立, 王斌, 向悟生, 文淑均, 李冬兴, 何运林. 弄岗喀斯特季节性雨林不同群丛物种多样性随海拔的变化. 生态学报, 2016, 36(14): 4509-4517. |
| [25] |
Du H, Peng W X, Song T Q, Zeng F P, Wang K L, Song M, Zhang H. Spatial pattern of woody plants and their environmental interpretation in the karst forest of southwest China. Plant Biosystems-An International Journal Dealing with all Aspects of Plant Biology, 2015, 149(1): 121-130. DOI:10.1080/11263504.2013.796019 |
| [26] |
Du H, Hu F, Zeng F P, Wang K L, Peng W X, Zhang H, Zeng Z X, Zhang F, Song T Q. Spatial distribution of tree species in evergreen-deciduous broadleaf karst forests in southwest China. Scientific Reports, 2017, 7(1): 15664. DOI:10.1038/s41598-017-15789-5 |
| [27] |
杨永, 刘冰, Njenga D M. 中国裸子植物物种濒危和保育现状. 生物多样性, 2017, 25(7): 758-764. |
| [28] |
Webb C O, Ackerly D D, McPeek M A, Donoghue M J. Phylogenies and community ecology. Annual Review of Ecology and Systematics, 2002, 33: 475-505. DOI:10.1146/annurev.ecolsys.33.010802.150448 |
| [29] |
Vamosi S M, Heard S B, Vamosi J C, Webb C O. Emerging patterns in the comparative analysis of phylogenetic community structure. Molecular Ecology, 2009, 18(4): 572-592. DOI:10.1111/j.1365-294X.2008.04001.x |
| [30] |
张凯, 陈丽茹, 徐慧敏, 李秧秧. 黄土高原水蚀风蚀交错带小流域植物群落特征的空间变异及其影响因素. 应用生态学报, 2019, 30(8): 2521-2530. |
| [31] |
Webb C O, Donoghue M J. Phylomatic: tree assembly for applied phylogenetics. Molecular Ecology Notes, 2005, 5(1): 181-183. DOI:10.1111/j.1471-8286.2004.00829.x |
| [32] |
Zanne A E, Tank D C, Cornwell W K, Eastman J M, Smith S A, FitzJohn R G, McGlinn D J, O'Meara B C, Moles A T, Reich P B, Royer D L, Soltis D E, Stevens P F, Westoby M, Wright I J, Aarssen L, Bertin R I, Calaminus A, Govaerts R, Hemmings F, Leishman M R, Oleksyn J, Soltis P S, Swenson N G, Warman L, Beaulieu J M. Three keys to the radiation of angiosperms into freezing environments. Nature, 2014, 506(7486): 89-92. DOI:10.1038/nature12872 |
| [33] |
Swenson N G, Enquist B J, Thompson J, Zimmerman J K. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities. Ecology, 2007, 88(7): 1770-1780. DOI:10.1890/06-1499.1 |
| [34] |
Webb C O, Ackerly D D, Kembel S W. Phylocom: software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics, 2008, 24(18): 2098-2100. DOI:10.1093/bioinformatics/btn358 |
| [35] |
Yost R S, Uehara G, Fox R L. Geostatistical analysis of soil chemical properties of large land areas. I. Semi-variograms. Soil Science Society of America Journal, 1982, 46(5): 1028-1032. DOI:10.2136/sssaj1982.03615995004600050028x |
| [36] |
Schneider K, Leopold U, Gerschlauer F, Barthold F, Giese M, Steffens M, Hoffmann C, Frede H G, Breuer L. Spatial and temporal variation of soil moisture in dependence of multiple environmental parameters in semi-arid grasslands. Plant and Soil, 2011, 340(1/2): 73-88. |
| [37] |
Cambardella C A, Moorman T B, Novak J M, Parkin T B, Karlen D L, Turco R F, Konopka A E. Field-scale variability of soil properties in central Iowa soils. Soil Science Society of America Journal, 1994, 58(5): 1501-1511. DOI:10.2136/sssaj1994.03615995005800050033x |
| [38] |
徐国策, 李占斌, 李鹏, 黄萍萍, 龙菲菲. 丹江中游典型小流域土壤总氮的空间分布. 地理学报, 2012, 67(11): 1547-1555. DOI:10.11821/xb201211011 |
| [39] |
Jiang Z H, Liu H Y, Wang H Y, Peng J, Meersmans J, Green S M, Quine T A, Wu X C, Song Z L. Bedrock geochemistry influences vegetation growth by regulating the regolith water holding capacity. Nature communications, 2020, 11: 2392. DOI:10.1038/s41467-020-16156-1 |
| [40] |
邵晓莉, 程毅康, 王茜茜, 王旭, 巫勇, 洪小江, 方燕山, 陆雍泉, 龙文兴. 海南岛热带云雾林地上生物量分布规律. 生态学杂志, 2018, 37(9): 2566-2572. |
| [41] |
夏焕柏. 茂兰喀斯特植被不同演替阶段的生物量和净初级生产力估算. 贵州林业科技, 2010, 38(2): 1-7, 49-49. |
| [42] |
Larson D W, Matthes U, Kelly P E. Cliff Ecology: Pattern and Process in Cliff Ecosystems. Cambridge: Cambridge University Press, 2000: 43-123, 175-245.
|
| [43] |
Tang C Q, Yang Y C, Ohsawa M, Momohara A, Yi S R, Robertson K, Song K, Zhang S Q, He L Y. Community structure and survival of tertiary relict Thuja sutchuenensis (Cupressaceae) in the subtropical Daba mountains, southwestern China. PLoS One, 2015, 10(4): e0125307. DOI:10.1371/journal.pone.0125307 |
| [44] |
彭旭东, 戴全厚, 李昌兰. 中国西南喀斯特坡地水土流失/漏失过程与机理研究进展. 水土保持学报, 2017, 31(5): 1-8. |
| [45] |
Wang B W, Zhang G H, Duan J. Relationship between topography and the distribution of understory vegetation in a Pinus massoniana forest in southern China. International Soil and Water Conservation Research, 2015, 3(4): 291-304. DOI:10.1016/j.iswcr.2015.10.002 |
| [46] |
石燕金, 周云超. 石漠化喀斯特皆伐迹地的土壤侵蚀特征. 中国水土保持科学, 2018, 16(5): 114-119. |
| [47] |
龙翠玲. 不同地形部位喀斯特森林物种多样性的比较研究——以贵州茂兰自然保护区为例. 中国岩溶, 2007, 26(1): 55-60. DOI:10.3969/j.issn.1001-4810.2007.01.009 |
| [48] |
Feng G, Svenning J C, Mi X C, Jia Q, Rao M D, Ren H B, Bebber D P, Ma K P. Anthropogenic disturbance shapes phylogenetic and functional tree community structure in a subtropical forest. Forest Ecology and Management, 2014, 313: 188-198. DOI:10.1016/j.foreco.2013.10.047 |
| [49] |
Willis C G, Halina M, Lehman C, Reich P B, Keen A, McCarthy S, Cavender-Bares J. Phylogenetic community structure in Minnesota oak savanna is influenced by spatial extent and environmental variation. Ecography, 2010, 33(3): 565-577. |
| [50] |
Swenson N G, Enquist B J, Pither J, Thompson J, Zimmerman J K. The problem and promise of scale dependency in community phylogenetics. Ecology, 2006, 87(10): 2418-2424. DOI:10.1890/0012-9658(2006)87[2418:TPAPOS]2.0.CO;2 |