 2021, Vol. 41
2021, Vol. 41文章信息
- 刘星宏, 张青青, 徐海量, 张鹏, 张广鹏, 李宏
- LIU Xinghong, ZHANG Qingqing, XU Hailiang, ZHANG Peng, ZHANG Guangpeng, LI Hong
- 北疆盐碱地植物群落空间分布及物种多样性
- Spatial distribution and species diversity of saline-alkali plant communities in northern Xinjiang
- 生态学报. 2021, 41(4): 1501-1513
- Acta Ecologica Sinica. 2021, 41(4): 1501-1513
- http://dx.doi.org/10.5846/stxb201912112690
-
文章历史
- 收稿日期: 2019-12-11
- 修订日期: 2020-07-31





2. 中国科学院新疆生态与地理研究, 乌鲁木齐 830011
2. Xinjiang Institute of Ecology and Geography, Urumqi 830011, China
土壤盐渍化是荒漠化的主要类型之一, 我国盐碱地总面积约为1亿hm2, 主要分布在东北松嫩平原、华北黄淮海平原、西北半干旱-半荒漠地区、西北干旱荒漠地区及滨海地区[1]。其中, 内陆盐碱地面积占全国盐碱地总面积的95.00%。新疆地处欧亚大陆腹地, 据统计新疆盐碱土面积占全国盐碱土总面积的22.01%, 是我国最为干旱, 降水量稀少、蒸发量强烈、土壤盐碱化分布范围最广、盐碱化类型最多、土壤积盐最重的地区[2]。由于新疆北疆大部分地区盐碱土多以石膏棕漠土和石膏盐盘棕漠土这两种土壤类型存在, 土壤表面含盐量较高, 受到环境影响易形成盐壳, 影响植物幼苗萌发, 还会对新疆北疆工业、农业发展产生极大不便。然而, 随着全球气候变化、不合理开垦荒地、大面积毁林以及过渡放牧等自然和人为因素干扰下, 盐碱地植物生态环境已遭受不同程度的影响和破坏, 地表植被退化、生物多样性下降、水土流失等,导致部分珍贵、稀有的物种已濒临灭绝或被逐渐被其他物种所替代[3]。因此, 人们对盐碱地植物资源的保护引起了极大的重视。许多学者从盐碱地植物群落类型和组成[4]、群落物种多样性和分布[5]、土壤理化性质[3, 6-7]等方面对北疆盐碱地植物展开大量调查和研究。学者们的研究区域多集中于北疆准噶尔盆地南缘的古尔班通古特沙漠[8-9]和玛纳斯河流域[1], 在准噶尔盆地西部的伊犁河谷和准噶盆地北部的乌伦古湖[10-11]也有研究, 由于研究区域较为分散且为小尺度区域研究, 其环境和群落结构差异较大, 因此对群落自身的特征和认识也各有偏重。
数量分类和排序分析方法能够系统研究植物群落分类及其内部特征[12-14]。植物群落数量分类和排序应用最普遍的方法有双向指示种分析(TWINSPAN, Two-way indicators species anailsis)法、除趋势对应分析(DCA, Detrended correspondence analysis)法、除趋势典范对应分析(DCCA, Detrended canonical correspondence analysis)法、典范对应分析(CCA, Canonical correspondence analysis)法等, 这些方法不仅能够客观反映植物群落的生态关系, 还能准确的反映群落分布与环境之间的生态关系, 已被广泛应用于森林、草地、荒漠、干旱河谷、湿地等植被研究[15-18]。目前有关盐碱地植物群落特征已有较多的研究, 但都集中于研究盐碱地植物区系组成[5-6, 9]、盐碱地植物的耐盐性[10-11]、盐碱地植物开发利用[19], 有关大尺度区域下盐碱地植物群落数量分类和多样性特征的报道较少。因此, 本研究应用TWINSPAN、DCA、CCA和方差分析, 对北疆盐碱地植物群落进行数量分类与排序, 探讨盐碱地植物群落分布与环境因子间的关系及群落物种多样性变化, 以期为绿洲-荒漠复合生态系统的平衡和维持提供数据。
1 研究区概况新疆地形特点是山脉与盆地相间排列, 盆地被高山环抱, 北为阿尔泰山, 南为昆仑山, 天山横亘中部[20]。研究区位于阿尔泰山与天山之间(图 1), 是一个略呈三角形的封闭式内陆盆地, 东西长700 km, 南北宽370 km, 面积为13.4×104 km2, 盆地腹部是中国第二大沙漠-古尔班通古特沙漠, 盆地南部为冲积扇平原, 盆地边缘为山麓绿洲[21]。研究区地属温带气候, 东部为寒潮通道, 冬季为我国同纬度最冷之地, 南部年均温为6—10℃, 无霜期150—190 d;北部、西部年均温3—5℃, 无霜期140 d。

|
| 图 1 研究区及调查样点分布 Fig. 1 Study area and survey sample distribution |
沿古尔班通古特沙漠边缘的荒漠-绿洲交错带进行采样且采样区域均为盐碱地, 主要调查区域(表 1)包括克拉玛依市、2个自治州(昌吉回族自治州, 博尔塔拉蒙古自治州)、2个地区(阿勒泰地区, 塔城地区)总共27个县市。
| 序号 Number |
调查区域 Survey area |
经纬度范围 Latitude and longitude range |
海拔范围/m Altitude range |
植被类型 Vegetation types |
所在行政区 Administrative district |
样点数量 Number of samples |
| 1 | 新疆阜康市 | N44°15′48.1"E87°52′24.3″—N44°20′25.1”E88°43′34.3″ | 441—574 | 荒漠 | 昌吉州 | 22 |
| 2 | 新疆吉木萨尔县 | N44°21′40.8″E88°51′46″—N44°09′22.7″E89°12′25.5″ | 532—612 | 荒漠 | 昌吉州 | 6 |
| 3 | 新疆奇台县 | N44°13′02.4″E89°19′20.7″—N44°05′43.3″E89°51′26.9″ | 597—765 | 荒漠 | 昌吉州 | 5 |
| 4 | 新疆木垒县 | N44°07′17.6″E90°11′22.6″—N44°10′14.4″E90°53′54.5″ | 801—1280 | 荒漠 | 昌吉州 | 12 |
| 5 | 新疆昌吉市 | N44°10′12.9″E87°42′39.4″—N44°27′30.4″E87°05′19.1″ | 406—429 | 荒漠 | 昌吉州 | 4 |
| 6 | 新疆沙湾县 | N44°28′55″E85°52′34.8″ | 375 | 荒漠 | 塔城地区 | 1 |
| 7 | 新疆博乐市 | N44°53′28.6″E82°07′52.6″—N44°08′21.3″E82°39′50.7″ | 194—496 | 荒漠 | 博尔塔拉州 | 5 |
| 8 | 新疆精河县 | N44°17′34.5″E82°47′22″—N44°34′51.7″E83°48′41.5″ | 200—369 | 荒漠 | 塔城地区 | 7 |
| 9 | 新疆乌苏县 | N44°40′35.5″E83°51′46.2″—N44°45′44.9″E84°54′06.3″ | 314—400 | 荒漠 | 塔城地区 | 7 |
| 10 | 新疆青河县 | N45°11′50.7″E90°06′50.7″—N45°36′20.6″E90°12′07.8″ | 1139—1222 | 荒漠 | 阿勒泰地区 | 3 |
| 11 | 新疆富蕴县 | N46°48′35.5″E89°34′07″—N47°09′04.3″ E88°45′26″ | 729—1001 | 荒漠 | 阿勒泰地区 | 4 |
| 12 | 新疆塔城市 | N46°39′41.1″E82°54′22.4″—N46°21′40.1″E83°46′39.9″ | 394—562 | 荒漠 | 塔城地区 | 5 |
| 13 | 新疆克拉玛依市 | N45°48′40.6″E85°20′49.7″—N45°11′01″E85°05′13.1″ | 247—269 | 荒漠 | 克拉玛依市 | 3 |
分别在每个调查样点设置一个10 m×10 m的样方, 进行乔木、灌木植物的调查, 在每个10 m×10 m样方内按梅花点采样法设置5个1 m×1 m的小样方, 用于草本植物的调查, 记录所有样方内的物种组成, 测定每种植物的高度、盖度、频度等指标, 再调查每个样点的同时用GPS记录样点的经纬度。测定植被的同时, 在各样地采用“S”形五点法采集土样。采集时除去凋落物, 用土钻按0—20 cm土层取样, 3次重复, 将采集到的土样装在自封袋带回实验室分析。
2.2 土壤、气象、地形因子数据获取将采集的土壤样品用1∶2.5土水比悬液酸度计测定土壤酸碱度(pH), 用1:5土水比浸提液测定土壤电导率, 获得土壤因子数据;通过国家气象数据中心(http://data.cma.cn/)下载新疆各个站点的气象数据, 根据采样点坐标, 从中筛选出54个站点近20年的年均降水量、年均气温、年均蒸发量数据, 利用Microsoft excel 2013软件进行加权平均得到多年平均降水量、多年平均温度、多年平均蒸发量, 再利用ArcGIS 10.4.1软件空间插值分析中的克里金法(Kriging)对气象数据进行插值提取各个采样点的气象因子数据;地形因子中的海拔高度通过GPS测定获取, 坡度和坡向利用ArcGIS 10.4.1中的3D Analyst—slope/aspect工具对新疆DEM影像进行处理并获取各个采样点的数据。
2.3 物种多样性测度与方法群落的物种多样性可反映群落中植物种类的丰富程度, 而群落物种多样性研究的测度常采用α-多样性和β-多样性体现[22]。α-多样性不仅能反映群落中物种丰富度和群落内物种组成还可反映物种在群落中分布的均匀程度。β-多样性反映2个群落或样地中共有种与总物种数之间的比例关系, 比值越大, 则说明2个群落或样地物种间的竞争越小, 群落越稳定。本研究采用α-多样性测度中的Margalef丰富度指数(DMa)、Simpson多样性指数(D)、Shannon-Wiener多样性指数(H′)、Pielou均匀度指数(J)[23-26], 比较各群落多样性的变化特征, 采用Sorenson′s相似性指数(So)和Jaccard′s相似性指数(Ja)测度群落或生境的β-多样性[27, 28], 比较各群落间相似性特点, 其计算公式如下:
Margalf丰富度指数(DMa):
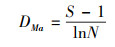
|
(1) |
Simpson优势度指数(D):
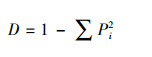
|
(2) |
Shannon-Wiener指数(H′):

|
(3) |
Pielou均匀度指数(J):
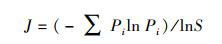
|
(4) |
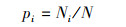
|
(5) |
式中, S为样方中的总物种数;N为样方中的总个体数;Ni为第i种植物的个体数;Pi为i种植物个体数占总个体数比例。
Sorenson′s相似性指数(So):
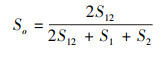
|
(6) |
Jaccard′s相似性指数(Ja):
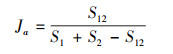
|
(7) |
式中, S12为2个群落都出现的物种数;S1和S2分别为群落1和群落2中的物种数。
2.4 数据分析北疆盐碱地植物群落数量分类和排序采用物种多度与样地组成的矩阵[29]作为基础数据进行计算, 利用Microsoft excel 2016软件对野外调查数据进行处理, 得到84个样点的物种×样地50×84和8个环境因素的环境×样地8×84矩阵数据, 采用WinTWINS 2.3软件[30]对物种-样地矩阵进行双向指示种分析(TWINSPAN)法, 对北疆盐碱地植物群落进行划分, 在划分过程中, 先将所有样地和种类分成0和1两大类, 然后这两大类再各自划分成两类, 以此类推, 直至达到所要求的划分水平为止。物种选用的假种5级分别为:0—2、2—5、5—10、10—20和>20, 选择最多划分水平为6, 每次划分最多分类数为2[31];采用CANOCO 5.0软件[32]中的除趋势对应分析(DCA)法和典范对应分析(CCA)法分析植物群落分布于环境因素间关系;群落多样性变化特征采用SPSS 20.0软件对其进行单因素方差分析(One-way ANOVA)[33]。
3 结果 3.1 群落数量分类利用TWINSPAN等级分类, 将84个样点划分为12个组分, 结合野外调查的实际情况和群落生境特征,采用第4级群丛水平的划分标准[34], 最终将其归并为11个群丛类型(图 2)。

|
| 图 2 盐碱地植物群落84个样点的TWINSPAN树状分类图 Fig. 2 TWINSPAN tree classification diagram of 84 sample points of saline-alkali plant communities E代表特征值, E stands for feature value; N代表样点数量, N stands for the number of samples; D代表分类次数, D stands for the number of classifications; “级数”表示分类水平数, ″level″ indicates the number of classification levels; 罗马数字代表群丛类型, Roman numerals indicate the type of cluster |
通过综合优势比大小确定各层优势种, 依据《中国植被》[34]的分类原则和系统, 对所划分的群丛进行命名, 各群丛类型主要特征及分布情况(表 2)如下:
| 群丛类型 Cluster type |
群丛名称 Cluster name |
所含样点 Included samples |
分布区域 Distribution area |
| I | 胡杨-盐生白刺-刺毛碱蓬群丛 | 60—64, 72 | 精河县、乌苏县、青河县 |
| II | 梭梭+囊果碱蓬+红砂群丛 | 1—3、5—12 | 阜康市 |
| III | 梭梭+红砂-散枝猪毛菜群丛 | 14, 15, 29, 41, 47—49, 53—56, 58, 59, 65—70, 82—84 | 克拉玛依市、昌吉市、阜康市、博乐市、精河县、奇台县、木垒县、乌苏县 |
| IV | 多枝柽柳-粗枝猪毛菜+叉毛蓬群丛 | 4, 23, 46 | 昌吉市、阜康市、吉木萨尔县 |
| V | 囊果碱蓬+红砂-散枝猪毛菜群丛 | 13, 16, 17, 24—26, 28 | 阜康市和吉木萨尔县 |
| VI | 红砂+高枝假木贼-叉毛蓬群丛 | 18, 19, 22, 27, 31—33, 37, 51, 52 | 阜康市、博乐市、吉木萨尔县、木垒县、奇台县 |
| VII | 盐穗木+里海盐爪爪-刺毛碱蓬群丛 | 20, 21, 50, 77 | 阜康市、塔城市、沙湾县 |
| VIII | 高枝假木贼-草原绢蒿+叉毛蓬群丛 | 34, 35, 38—40, 43 | 木垒县 |
| IX | 草原绢蒿群丛 | 36, 42, 73, 78—80 | 塔城市、木垒县、富蕴县 |
| X | 同齿樟味藜-草原绢蒿+角果藜群丛 | 44, 74—76, 81 | 塔城市、木垒县、富蕴县 |
| XI | 盐节木+高枝假木贼群丛 | 30, 45, 57, 71 | 精河县、木垒县、奇台县、青河县 |
类型I:胡杨-盐生白刺-刺毛碱蓬群丛。含样点60—64、72, 分布于精河县、乌苏县、青河县, 海拔高度332.70—368.80 m, 土壤呈碱性(pH>8.5), 具有灰漠土和棕钙土两种类型土壤, 土壤电导率为0.17—10.23 ms/cm, 多年平均降水量为220.47—221.84 mm, 多年平均蒸发量为1341.94—1827.95 mm。乔木层优势种为胡杨(Populus euphratica), 平均高度为560.00 cm;灌木层优势种为盐生白刺(Nitraria sphaerocarpa), 平均高度为38.75 cm;草本层优势种为刺毛碱蓬(Suaeda acuminata), 平均高度为5.93 cm。群丛总盖度相差较大, 范围为20%—90%, 平均盖度61%。
类型II:梭梭+囊果碱蓬+红砂群丛。含样点1—3、5—12, 分布于阜康市, 海拔高度441.00—546.00 m, 土壤呈强碱性(8.5<pH<9.5), 具有灰漠土和风沙土两种类型土壤, 土壤电导率为1.48—5.19 ms/cm, 多年平均降水量为209.01—254.08 mm, 多年平均蒸发量为1542.41—1735.26 mm。灌木层优势种较多, 有梭梭、囊果碱蓬(S.physophora)、红砂(Reaumuria songarica)、盐爪爪(Kalidium foliatum), 平均高度分别为164.00、38.50、27.66、40.66 cm。群丛总盖度相差较大, 范围为30%—90%, 平均盖度65%。
类型III:梭梭+红砂-散枝猪毛菜群丛。含样点14, 15、29、41、47—49、53—56、58, 59、65—70、82—84, 分布于克拉玛依市、昌吉市、阜康市、博乐市及精河县、奇台县、木垒县、乌苏县, 海拔高度193.60—1222.00 m, 土壤呈强碱性(8.5<pH<9.5), 具有灰漠土、灰棕漠土、风沙土、棕钙土4种类型土壤, 土壤电导率为0.17—10.68 ms/cm, 多年平均降水量为134.87—223.73 mm, 多年平均蒸发量为1055.24—5733.24 mm。灌木层的优势种为梭梭和红砂, 平均高度分别为89.65、30.44 cm;草本层优势种为散枝猪毛菜(Salsola brachiata)和粗枝猪毛菜(S.subcrassa), 平均高度分别为27.33、8.56 cm。群丛总盖度相差较大, 范围为10%—90%, 平均盖度43%。
类型IV:多枝柽柳-粗枝猪毛菜+叉毛蓬群丛。含样点4、23、46, 分布于昌吉市、阜康市、吉木萨尔县, 海拔高度429.00—532.00 m, 土壤呈极强碱性(pH>9.5), 具有灰漠土一种类型土壤, 土壤电导率为3.48—6.80 ms/cm, 多年平均降水量为205.51—237.04 mm, 多年平均蒸发量为1699.44—1806.49 mm。灌木层的优势种为多枝柽柳(Tamarix ramosissima), 平均高度为126.00 cm;草本层优势种为粗枝猪毛菜和叉毛蓬(Petrosimonia sibirica), 平均高度分别为8.26 cm、6.53 cm。群丛总盖度较大, 为60%—80%, 平均盖度70%。
类型V:囊果碱蓬+红砂-散枝猪毛菜群丛。含样点13、16, 17、24—26、28, 分布于阜康市和吉木萨尔县, 海拔高度526.00—604.00 m, 土壤呈强碱性(8.5<pH<9.5), 其土壤类型为灰漠土, 土壤电导率为1.08—4.75 ms/cm, 多年平均降水量为192.64—241.69 mm, 多年平均蒸发量为1733.13—6342.27 mm。灌木层的优势种为囊果碱蓬和红砂, 平均高度分别为55.50、25.66 cm;草本层优势种为散枝猪毛菜, 平均高度为6.50 cm。群丛总盖度, 为30%—70%, 平均盖度47%。
类型VI:红砂+高枝假木贼-叉毛蓬群丛。含样点18, 19、22、27、31—33、37、51, 52, 分布于阜康市、博乐市、吉木萨尔县、木垒县、奇台县, 海拔高度374.00—942.00 m, 土壤呈强碱性(8.5<pH<9.5), 具有灰漠土和灰棕漠土两种类型土壤, 土壤电导率为0.20—8.03 ms/cm, 多年平均降水量为130.45—279.55 mm, 多年平均蒸发量为1382.13—6547.47 mm。灌木层的优势种为红砂、高枝假木贼(Anabasis elatior)、梭梭, 平均高度分别为33.00、16.00、220.00 cm;草本层优势种为叉毛蓬, 平均高度为6.94 cm。群丛总盖度较大, 为50%—90%, 平均盖度71%。
类型VII:盐穗木+里海盐爪爪-刺毛碱蓬群丛。含样点20, 21、50、77, 分布于阜康市、塔城市、沙湾县, 海拔高度375.00—468.00 m, 土壤呈强碱性(8.5<pH<9.5), 具有灰漠土和棕钙土两种类型土壤, 土壤电导率为3.21—5.35 ms/cm, 多年平均降水量为188.78—266.95 mm, 多年平均蒸发量为1080.08—5100.59 mm。灌木层的优势种为盐穗木(Halostachys caspica)、里海盐爪爪(K.caspicum), 平均高度分别为80.00、70.00 cm;草本层优势种为刺毛碱蓬和狗牙根(Cynodon dactylon), 平均高度分别为51.46、13.20 cm。群丛总盖度相差较大, 为30%—90%, 平均盖度65%。
类型VIII:高枝假木贼-草原绢蒿+叉毛蓬群丛。含样点34, 35、38—40、43, 分布于木垒县, 海拔高度801.00—1112.00 m, 土壤呈强碱性(8.5<pH<9.5), 其土壤类型为灰漠土, 土壤电导率为0.50—2.18 ms/cm, 多年平均降水量为235.88—275.74 mm, 多年平均蒸发量为1893.37—1941.13 mm。灌木层的优势种为高枝假木贼, 平均高度为19.33 cm;草本层优势种为草原绢蒿(Seriphidium schrenkianum)和叉毛蓬, 平均高度分别为6.00、2.64 cm。群丛总盖度较大, 为60%—80%, 平均盖度70%。
类型IX:草原绢蒿群丛。含样点36、42、73、78—80, 分布于塔城市、木垒县、富蕴县, 海拔高度393.50—1280 m, 土壤呈碱性(7.5<pH<8.5), 具有灰漠土和棕钙土两种类型土壤, 土壤电导率为0.43—5.13 ms/cm, 多年平均降水量为188.99—278.37 mm, 多年平均蒸发量为1117.62—1945.43 mm。优势种为草本层的草原绢蒿, 平均高度为30.83 cm。群丛总盖度相差较大, 为30%—90%, 平均盖度58%。
类型X:同齿樟味藜-草原绢蒿+角果藜群丛。该群丛含样点44、74—76、81, 分布于塔城市、木垒县、富蕴县, 海拔高度562.20—1037.00 m, 土壤呈强碱性(8.5<pH<9.5), 具有灰漠土和棕钙土两种类型土壤,土壤电导率为0.51—2.53 ms/cm, 多年平均降水量为142.92—207.21 mm, 多年平均蒸发量为544.37—8099.05 mm。灌木层优势种为同齿樟味藜(Camphorosma monspeliaca), 平均高度为4.40 cm;草本层的优势种为草原绢蒿和角果藜(Ceratocarpus arenarius), 平均高度分别为22.60 cm、3.20 cm。群丛总盖度较大, 为40%—70%, 平均盖度56%。
类型XI:盐节木+高枝假木贼群丛。含样点30、45、57、71, 分布于精河县、木垒县、奇台县、青河县, 海拔高度204.00—1197.00 m, 土壤呈强碱性(8.5<pH<9.5), 土壤电导率为0.20—1.38 ms/cm, 具有灰漠土、灰棕漠土和棕钙土三种类型土壤, 多年平均降水量为187.99—215.42 mm, 多年平均蒸发量为1353.54—6151.62 mm。优势种为灌木层的盐节木(Halocnemum strobilaceum)和高枝假木贼, 平均高度分别为18.66、16.33 cm。群丛总盖度相差较大, 为13%—60%, 平均盖度43%。
3.2 群落多样性变化特征通过TWINSPAN聚类所划分11个植物群丛的α-物种多样性变化分析结果见图 3。由图可知, 各群丛类型Margalef丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数、Pielou均匀度指数整体变化一致, 均表现为下降趋势。各群丛Simpson多样性指数变化表现为, 群丛VII>群丛I>群丛V>群丛III>群丛II>群丛IV>群丛VIII>群丛XI>群丛VI>群丛IX>群丛X, 而Shannon-Wiener多样性指数变化则表现为群丛VII>群丛I>群丛V>群丛VIII>群丛III>群丛II>群丛IV>群丛VI>群丛IX>群丛XI>群丛X, 其中群丛VII的多样性最大, Simpson多样性指数和Shannon-Wiener多样性指数分别为0.595、1.182;其次是群丛I, 指数分别为0.582、1.159;群丛X的多样性最小, 指数分别为0.213、0.375, 群丛VII的Simpson多样性指数和Shannon-Wiener多样性指数比群丛X分别高出47.27%和51.81%, 群丛VII和群丛I多样性显著高于群丛X(P<0.05);各群丛Pielou均匀度指数变化表现为, 群丛VII>群丛I>群丛V>群丛III>群丛IV>群丛II>群丛VIII>群丛IX>群丛VI>群丛XI>群丛X, 物种在群丛VII和群丛I的分布比较均匀, 指数分别为0.686、0.681, 群丛VII和群丛I均匀度显著高于群丛X(P<0.05);各群丛类型的Margalef丰富度指数变化表现为, 群丛I>群丛V>群丛VII>群丛III>群丛IV>群丛VIII>群丛II>群丛VI>群丛XI>群丛IX>群丛X, 群丛I丰富度显著高于群丛X(P<0.05)。

|
| 图 3 11个盐碱地植物群丛Simpson指数、Shannon-Wiener指数、Pielou均匀度指数、Margalef丰富度指数的变化特征 Fig. 3 Sampson index, Shannon-Wiener index, Pielou evenness index, Margalef richness of 11 saline-alkali flora Index change characteristics 类型I:胡杨-盐生白刺-刺毛碱蓬群丛; 类型II:梭梭+囊果碱蓬+琵琶柴群丛; 类型III:梭梭+琵琶柴-散枝猪毛菜群丛; 类型IV:多枝柽柳-粗枝猪毛菜+叉毛蓬群丛; 类型V:囊果碱蓬+琵琶柴-散枝猪毛菜群丛; 类型VI:琵琶柴+高枝假木贼-叉毛蓬群丛; 类型VII:盐穗木+里海盐爪爪-刺毛碱蓬群丛; 类型VIII:高枝假木贼-草原娟蒿+叉毛蓬群丛; 类型IX:草原娟蒿群丛; 类型X:同齿樟味藜-草原娟蒿+角果藜群丛;类型XI:盐节木+高枝假木贼群丛 |
各群丛类型β-多样性分析结果(表 3和表 4), 由表可知, 群丛V和群丛VII的相似性最高, Sorenson′s指数和Jaccard′s指数分别为0.368、0.412, 其次为群丛V和群丛VI, 指数分别为0.364、0.400, 其原因为群丛V与群丛VII中共有植物种类较多, 两个群丛中分布的植物种类比较相似, 海拔高度、土壤酸碱度、多年平均降水量之间差异不大, 说明群丛V与群丛VII物种间的竞争较小, 群丛稳定性高于其他群丛。群丛VII和群丛X的相似性最低, 指数均为0, 主要原因为群丛VII与群丛X中没有出现共有物种, 海拔高度、土壤酸碱度、多年平均降水量、多年平均温度、多年平均蒸发量之间差异较大, 说明两群丛物种间存在较大竞争, 群丛稳定性低且与其他群丛之间不具有相似性。
| Sorenson′s相似性 Sorenson′s similarity |
群丛I Assoc.I |
群丛II Assoc.II |
群丛III Assoc.III |
群丛IV Assoc.IV |
群丛V Assoc.V |
群丛VI Assoc.VI |
群丛VII Assoc.VII |
群丛VIII Assoc.VIII |
群丛IX Assoc.IX |
群丛X Assoc.X |
| 群丛II Assoc.II | 0.350 | |||||||||
| 群丛III Assoc.III | 0.291 | 0.235 | ||||||||
| 群丛IV Assoc.IV | 0.240 | 0.345 | 0.238 | |||||||
| 群丛V Assoc.V | 0.316 | 0.316 | 0.291 | 0.296 | ||||||
| 群丛VI Assoc.VI | 0.300 | 0.300 | 0.349 | 0.276 | 0.364 | |||||
| 群丛VII Assoc.VII | 0.333 | 0.333 | 0.245 | 0.261 | 0.368 | 0.316 | ||||
| 群丛VIII Assoc.VIII | 0.125 | 0.176 | 0.255 | 0.222 | 0.263 | 0.286 | 0.133 | |||
| 群丛IX Assoc.IX | 0.154 | 0.214 | 0.186 | 0.211 | 0.267 | 0.294 | 0.167 | 0.294 | ||
| 群丛X Assoc.X | 0.091 | 0.167 | 0.154 | 0.133 | 0.231 | 0.154 | 0.000 | 0.214 | 0.333 | |
| 群丛XI Assoc.XI | 0.154 | 0.083 | 0.255 | 0.118 | 0.214 | 0.143 | 0.091 | 0.200 | 0.182 | 0.200 |
| Jaccard′s相似性 Jaccard′s similarity |
群丛I Assoc.I |
群丛II Assoc.II |
群丛III Assoc.III |
群丛IV Assoc.IV |
群丛V Assoc.V |
群丛VI Assoc.VI |
群丛VII Assoc.VII |
群丛VIII Assoc.VIII |
群丛IX Assoc.IX |
群丛X Assoc.X |
| 群丛II Assoc.II | 0.368 | |||||||||
| 群丛III Assoc.III | 0.258 | 0.182 | ||||||||
| 群丛IV Assoc.IV | 0.188 | 0.357 | 0.185 | |||||||
| 群丛V Assoc.V | 0.300 | 0.300 | 0.258 | 0.267 | ||||||
| 群丛VI Assoc.VI | 0.273 | 0.273 | 0.367 | 0.235 | 0.400 | |||||
| 群丛VII Assoc.VII | 0.333 | 0.333 | 0.194 | 0.214 | 0.412 | 0.300 | ||||
| 群丛VIII Assoc.VIII | 0.077 | 0.120 | 0.206 | 0.167 | 0.217 | 0.250 | 0.083 | |||
| 群丛IX Assoc.IX | 0.100 | 0.158 | 0.129 | 0.154 | 0.222 | 0.263 | 0.111 | 0.263 | ||
| 群丛X Assoc.X | 0.053 | 0.111 | 0.100 | 0.083 | 0.176 | 0.100 | 0.000 | 0.158 | 0.333 | |
| 群丛XI Assoc.XI | 0.100 | 0.048 | 0.207 | 0.071 | 0.158 | 0.091 | 0.053 | 0.143 | 0.125 | 0.143 |
采用物种-样地矩阵数据对TWINSPAN所划分的11种群丛进行DCA排序, 发现第1轴的梯度长度为3.85<4.0(当第1轴长度的值>4.0时, 应选单峰模型;在3.0—4.0之间, 选单峰和线性模型均可;若小于3.0, 则选线性模型)满足单峰模型, 故本研究选用单峰模型对84个样点及50种盐碱地植物进行DCA排序分析。结果显示, 4个排序轴的特征值分别为0.5475、0.5157、0.3816、0.2967, 其中第1排序轴特征值最大, 第2排序轴次之, 因此采用前两个排序轴做二维排序图。由图 4可知, DCA排序结果与TWINSPAN分类所产生的各群丛类型基本吻合且在二维排序图中能较好地显示出来,排序结果与植物群丛分类结果有很大的相似性,说明DCA排序能较好的反映出实际观测情况。

|
| 图 4 84个样点DCA和CCA的二维排序图 Fig. 4 Two-dimensional sorting map of 84 samples DCA and CCA DCA: 除趋势对应分析;CCA典范对应分析 |
利用环-样地矩阵数据对北疆盐碱地84个采样点进行CCA排序, 分析多年平均温度和多年平均降水量、多年平均蒸发量、土壤酸碱度、土壤电导率、海拔高度、坡度、坡向等8个环境因子与11个盐碱地植物群丛分布相关关系。CCA排序分析中, 箭头代表各个环境因子, 箭头所处象限代表环境因子与排序轴间的正负相关性, 可根据箭头连线长短判别群丛分布与该环境因子相关性的大小, 具体表现为连线越长, 其相关性越大, 连线越短, 其相关性越小;此外, 两个箭头之间的夹角大小也反映出两个环境因子间相关性的大小, 表现为夹角越小, 相关性越大[35]。
由图 4和表 5可以看出, 蒙特卡洛(Monte Carlo)检验结果显示所有典范轴的显著性均达到显著水平(P=0.0020)。海拔高度与CCA第一排序轴呈极显著正相关(P<0.001), 相关系数为0.4513, 土壤电导率和多年平均温度、多年平均蒸发量与第一排序轴呈极显著负相关(P<0.001), 相关系数分别为-0.3868、-0.4659、-0.7632, 第一排序轴主要反映了多年平均蒸发量、海拔高度、土壤电导率、多年平均温度的变化, 其中多年平均蒸发量对各群丛分布具有较高的相关性;海拔高度和多年平均降水量与CCA第二排序轴呈极显著正相关(P<0.001), 相关系数分别为0.5826、0.3785, 多年平均蒸发量与CCA第二排序轴呈显著正相关(P<0.05), 相关系数为0.2127, 多年平均温度与CCA第二排序轴呈显著负相关(P<0.01), 相关系数为-0.3679, 第二排序轴反映了海拔高度、多年平均降水量、多年平均蒸发量、多年平均温度的变化, 其中海拔高度与各群丛分布的相关性较大;土壤酸碱度与CCA第三排序轴呈显著正相关(P<0.001), 相关系数为0.5012, 坡向与第三排序轴呈显著正相关(P<0.05), 相关系数为0.2209, 与第三排序轴相关性较大的环境因子是土壤酸碱度;土壤电导率与CCA第四排序轴呈显著正相关(P<0.05), 相关系数为0.2638, 多年平均温度与第四排序轴呈显著负相关(P<0.01), 相关系数为-0.2992, 多年平均蒸发量也与第四排序轴呈显著负相关(P<0.05), 相关系数为-0.2562, 与第四排序轴相关性较大的环境因子是多年平均温度。根据前4个排序轴的相关系数和各个环境因子在图上显示的结果, 可知, 各环境因子影响群丛分布大小依次为海拔高度>多年平均蒸发量>多年平均温度>多年平均降水量>土壤电导率>土壤酸碱度>坡向>坡度, 其中主要影响各群丛分布的环境因子是海拔高度、多年平均温度、多年平均降水量和蒸发量、土壤电导率, 8个环境因子中坡度与前4个排序轴无显著关系, 对群丛分布的影响不大。
| 环境因子 Environmental Factors |
第一轴 Aixs 1 |
第二轴 Aixs 2 |
第三轴 Aixs 3 |
第四轴 Aixs 4 |
| 酸碱度pH | 0.0436 | 0.1463 | 0.5012*** | 0.0450 |
| 土壤电导率Soil electrical conductivity | -0.3868*** | -0.0838 | 0.0705 | 0.2638* |
| 海拔高度Altitude | 0.4513*** | 0.5826*** | -0.1127 | 0.1102 |
| 坡度Slope | -0.0594 | -0.0768 | 0.0154 | -0.0850 |
| 坡向Aspect | 0.0179 | -0.1678 | 0.2209* | 0.0433 |
| 多年平均温度Annual average temperature | -0.4659*** | -0.3679*** | 0.1712 | -0.2992** |
| 多年平均降水量Annual average precipitation | 0.2175* | 0.3785*** | -0.1280 | -0.1709 |
| 多年平均蒸发量Annual average evaporation | -0.7632*** | 0.2127* | -0.0740 | -0.2562* |
| 特征值Eigenvalues | 0.7000 | 0.4530 | 0.3590 | 0.2860 |
| 物种与环境相关系数Species-environment correlations | 0.8970 | 0.7670 | 0.7340 | 0.6160 |
| 物种与环境关系方差解释的累计比例/% Cumulative proportion of variance interpretation of species- environment relationship |
29.0 | 47.7 | 62.6 | 74.4 |
| 所有典范轴的显著性检验Test of significance of all canonical axes | 0.0020 | |||
| *、**和***分别表示相关性在P<0.05、P<0.01和P<0.001水平具有统计学意义 | ||||
本研究利用数量生态学方法对北疆盐碱地植物群落进行数量分析。通过TWINSPAN等级分类和DCA排序,可以看出,二者结合能客观准确的划分群落类型, 两种方法可互相对比验证;TWINSPAN分类与DCA排序二者结果的一致性也说明了这两种方法对北疆盐碱地84个样点的盐碱地植物群落数量分类的可靠性。CCA排序分析的前2个排序轴中, 物种与环境因子的相关系数分别为0.8970和0.7670, 说明前2个排序轴能更好的说明环境因子与群落分布间的关系。一般的, 前3个排序轴的物种与环境关系方差累积贡献率在40%以上, 则说明排序效果较好[36], 本研究所得出前3个轴的方差累积贡献率达到62.6%, 由此说明CCA排序能较好的反映所选环境因子与群落分布间的关系且排序结果是可靠的。
通过研究结果发现, 影响北疆盐碱地植物群落分布的主要环境因素是海拔高度,其次是多年平均温度、降水量、蒸发量和土壤电导率, 在所选环境因子中地形因子坡度对盐碱地植物群落的分布影响不大, 其原因是研究区域内某些调查样点存在特殊生境所导致的。根据TWINSPAN分类结果可以看出,各个群丛在空间上表现为不连续性, 一些在地理上相距较远的样点被分成了相同的群丛(例如群丛X所含样点包含新疆塔城地区、阿勒泰地区、昌吉回族自治州), 由此可知, 群丛分布与研究区域内复杂的地形条件和气候有关。研究区位于新疆北疆地区, 海拔和气象因子在很大程度上影响着植物群落的分布[37]。大部分学者认为, 植物群落的形成是地形、气候和土壤因子的综合作用, 但在不同尺度区域下, 影响植物群落分布的主要环境因素是不同的[38]。在大尺度区域内, 影响植物群落分布的主要环境因子为气候因子, 而在小区域尺度内, 则是地形和土壤因子影响着植物群落的分布[39-40]。桂东伟等[41]研究中昆仑山北坡策勒河流域生态因素对植物群落的影响时发现, 群落分布主要受海拔的影响, 其次是受土壤因子的影响;包小婷等[42]对拉萨河流域植物群落的数量分类与排序的研究中得出, 影响植物群落分布的主要因素是年均温度和海拔,其次是年均降水量。上述研究结果与本研究所得出环境因子对群丛分布的结果基本一致。姚帅臣等[14]对拉萨河谷草地群落的研究表明, 影响草地群落分布格局的主要环境因子是海拔和坡向;范得芳等[43]对秦岭锐齿栎群落分布的研究中得出坡度和土壤全磷对群落分布影响最大, 而本研究结果发现地形因子对植物群落分布的影响不显著, 其原因是研究对象和研究尺度的不同, 本研究区域尺度较大且地形、气候较为复杂, 环境条件存在较大差异。从CCA排序轴结果上看, 前两个排序轴仅解释了47.7%, 这可能与研究区某一小尺度区域内土壤理化因子和地理隔离[44]等因素有关。
| [1] |
郗金标, 张福锁, 毛达如, 田长彦, 宋玉民, 刘德玺. 新疆盐渍土分布与盐生植物资源. 土壤通报, 2005, 36(3): 299-303. |
| [2] |
张添佑, 王玲, 罗冲, 彭丽. 玛纳斯河流域土壤盐渍化时空动态变化. 水土保持研究, 2016, 23(1): 228-233. |
| [3] |
胡文杰, 李跃进, 刘洪波, 张昊, 蒙荣, 芦永兴. 土默川平原土壤盐渍化与盐生植物分布及盐分离子特征的研究. 干旱区资源与环境, 2012, 26(4): 127-131. |
| [4] |
张玲. 河北滨海地区白刺及碱蓬属盐生植物营养成分分析[D]. 保定: 河北农业大学, 2013.
|
| [5] |
傅德平, 吕光辉, 杨建军. 克拉玛依植物群落结构和物种多样性研究. 干旱区资源与环境, 2008, 22(6): 152-155. |
| [6] |
王荷生. 新疆主要盐生植物羣落的分布及其与土壤、地下水的关系. 植物生态学与地植物学丛刊, 1964, 2(1): 57-69. |
| [7] |
麦尔哈巴·尼加提, 戴岳, 师庆东, 肖合来提·巴义, 彭丽萍, 尼加提·卡斯木. 准噶尔盆地东南缘荒漠灌丛盐生假木贼周围土壤理化性质研究. 灌溉排水学报, 2019, 38(7): 38-44. |
| [8] |
陈莎莎, 姚世响, 袁军文, 谷丽丽, 油天钰, 兰海燕, 张富春. 新疆荒漠地区盐生植物灰绿藜种子的萌发特性及其对生境的适应性. 植物生理学通讯, 2010, 46(1): 75-79. |
| [9] |
张立运. 准噶尔盆地南缘盐生植被的类型和组成. 干旱区研究, 1985, 2(1): 44-50. |
| [10] |
尹传华, 田长彦, 张锁福, 郗金标. 新疆部分盐生植物元素特征与土壤环境关系//西部地区第二届植物科学与开发学术讨论会论文摘要集. 乌鲁木齐: 中国植物学会, 2001: 139-139.
|
| [11] |
郗金标, 张福锁, 毛达如, 田长彦, 董振成, 王开芳. 新疆盐生植物群落物种多样性及其分布规律的初步研究. 林业科学, 2006, 42(10): 6-12. |
| [12] |
姚帅臣, 王景升, 丁陆彬, 包小婷, 李超, 王彤, 刘文婧, 李妍妍. 拉萨河谷草地群落的数量分类与排序. 生态学报, 2018, 38(13): 4779-4788. |
| [13] |
陈宝瑞, 朱玉霞, 张宏斌, 周磊, 辛晓平. 呼伦贝尔草甸草原植被的数量分类和排序研究. 武汉植物学研究, 2008, 26(5): 476-481. |
| [14] |
王静娅, 王明亮, 张凤华. 干旱区典型盐生植物群落下土壤微生物群落特征. 生态学报, 2016, 36(8): 2363-2372. |
| [15] |
张峰, 张金屯. 历山自然保护区猪尾沟森林群落植被格局及环境解释. 生态学报, 2003, 23(3): 421-427. |
| [16] |
潘晓玲, 张远东, 初雨, 韦如意. 塔里木河流域荒漠河岸林群落的多元分析与环境解释. 西北植物学报, 2001, 21(2): 245-251. |
| [17] |
鲁为华, 于磊, 蒋惠. 新疆昭苏县沙尔套山天然草地植物群落数量分类与排序(简报). 草业学报, 2008, 17(1): 135-139. |
| [18] |
贾希洋, 马红彬, 周瑶, 张蕊, 宿婷婷, 张双乔, 张俊. 不同生态恢复措施下宁夏黄土丘陵区典型草原植物群落数量分类和演替. 草业学报, 2018, 27(2): 15-25. |
| [19] |
胡文象, 张立运, 陈昌笃, 等. 准噶尔盆地南部植被类型、结构、生产力和开发利用考察研究. 中国科学院新疆生态与地理研究所: 2010年以前数据, 1989.
|
| [20] |
李小明. 新疆植被分布规律与水热关系初探. 干旱区研究, 1988, 5(2): 41-46. |
| [21] |
王敏芳, 焦养泉, 任建业, 佟殿君, 徐志诚. 准噶尔盆地侏罗纪沉降特征及其与构造演化的关系. 石油学报, 2007, 28(1): 27-32. |
| [22] |
李军玲, 张金屯. 太行山中段植物群落物种多样性与环境的关系. 应用与环境生物学报, 2006, 12(6): 766-771. |
| [23] |
Magurran A E. Ecological Diversity and Its Measurement. Princeton: Princeton University Press, 1988.
|
| [24] |
方炜, 彭少麟. 鼎湖山马尾松群落演替过程物种变化之研究. 热带亚热带植物学报, 1995, 3(4): 30-37. |
| [25] |
彭少麟, 周厚诚, 陈天杏, 郭少聪. 广东森林群落的组成结构数量特征. 植物生态学与地植物学学报, 1989, 13(1): 10-17. |
| [26] |
马克平, 黄建辉, 于顺利, 陈灵芝. 北京东灵山地区植物群落多样性的研究: Ⅱ丰富度、均匀度和物种多样性指数. 生态学报, 1995, 15(3): 268-277. |
| [27] |
赵志模, 郭依泉. 群落生态学原理与方法. 重庆: 科学技术文献出版社重庆分社, 1990.
|
| [28] |
阳含熙, 卢泽愚. 植物生态学的数量分类方法. 北京: 科学出版社, 1981.
|
| [29] |
王忠欣, 栾兆擎, 刘贵花. 洪河国家级自然保护区浓江河滨河湿地植物对土壤环境因子的响应. 湿地科学, 2013, 11(1): 54-59. |
| [30] |
van Kleef H H, Verberk W C E P, Leuven R S E W, Esselink H, van der Velde G, van Duinen G A. Biological traits successfully predict the effects of restoration management on macroinvertebrates in shallow softwater lakes. Hydrobiologia, 2006, 565(1): 210-216. DOI:10.1007/s10750-005-1914-6 |
| [31] |
张元明, 陈亚宁, 张道远. 塔里木河中游植物群落与环境因子的关系. 地理学报, 2003, 58(1): 109-118. |
| [32] |
El-Bana M, Khedr A H, van Hecke P, Bogaert J. Vegetation composition of a threatened hypersaline lake (Lake Bardawil), North Sinai. Plant Ecology, 2002, 163(1): 63-75. DOI:10.1023/A:1020351704409 |
| [33] |
钟韩珊, 李元哲, 张晓雨, 赵美玉, 赵丽娅. 不同类型沙地植物群落特征及多样性研究. 湖北大学学报: 自然科学版, 2019, 41(4): 349-355. |
| [34] |
吴征镒. 中国植被. 北京: 科学出版社, 1980.
|
| [35] |
张金屯. 数量生态学. 北京: 科学出版社, 2004.
|
| [36] |
Fernández-Aláez C, Fernández-Aláez M, Garcáa-Criado F. Spatial distribution pattern of the riparian vegetation in a basin in the NW spain. Plant Ecology, 2005, 179(1): 31-42. |
| [37] |
赵从举, 康慕谊, 雷加强. 准噶尔盆地典型地段植物群落及其与环境因子的关系. 生态学报, 2011, 31(10): 2669-2677. |
| [38] |
Siefert A, Ravenscroft C, Althoff D, Alvarez-Yépiz J C, Carter B E, Glennon K L, Heberling J M, Jo I S, Pontes A, Sauer A, Willis A, Fridley J D. Scale dependence of vegetation-environment relationships: a meta-analysis of multivariate data. Journal of Vegetation Science, 2012, 23(5): 942-951. |
| [39] |
van Couwenberghe R, Collet C, Lacombe E, Pierrat J C, Gégout J C. Gap partitioning among temperate tree species across a regional soil gradient in windstorm-disturbed forests. Forest Ecology and Management, 2010, 260(1): 146-154. |
| [40] |
段后浪, 赵安, 姚忠. 排序法在植物群落与环境关系研究中的应用述评. 热带亚热带植物学报, 2017, 25(2): 202-208. |
| [41] |
桂东伟, 雷加强, 曾凡江, 江源, 穆桂金, 杨发相. 中昆仑山北坡策勒河流域生态因素对植物群落的影响. 草业学报, 2010, 19(3): 38-46. |
| [42] |
包小婷, 丁陆彬, 姚帅臣, 王景升, 石培礼, 王彤, 李超, 刘文婧. 拉萨河流域植物群落的数量分类与排序. 生态学报, 2019, 39(3): 779-786. |
| [43] |
范得芳, 王得祥, 柴宗政, 胡有宁, 保积存, 王涛. 环境因子对秦岭锐齿栎群落物种分布及多样性的影响. 西北农林科技大学学报: 自然科学版, 2016, 44(10): 59-67. |
| [44] |
沈泽昊, 张新时. 中国亚热带地区植物区系地理成分及其空间析局的数量分析. 植物分类学报, 2000, 38(4): 366-368. |