 2021, Vol. 41
2021, Vol. 41文章信息
- 胡文浩, 张晓婧, 陈雅杰, 宇振荣, 段美春
- HU Wenhao, ZHANG Xiaojing, CHEN Yajie, YU Zhenrong, DUAN Meichun
- 坝上地区不同年代退耕还林生境的草本层植物多样性及影响因子
- Herb diversity and its impact factors in different periods from 1970s to 2000s of the returning farmland to forest project in the Bashang Area of North China
- 生态学报. 2021, 41(3): 1116-1126
- Acta Ecologica Sinica. 2021, 41(3): 1116-1126
- http://dx.doi.org/10.5846/stxb201912042631
-
文章历史
- 收稿日期: 2019-12-04
- 网络出版日期: 2020-12-14





2. 西南大学农学与生物科技学院, 重庆 400715
2. College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China
坝上地区地处华北西北部、地势复杂, 是首都乃至整个华北地区的重要生态屏障, 但由于开垦过度、土地沙化等原因, 出现了水土流失与地质灾害等问题, 严重威胁华北地区生态安全[1]。我国在不同时期于坝上地区实施了退耕还林工程(Returning farmland to forest project, RFTF)[2-3], 旨在通过栽植人工林, 将产量不稳、土地贫瘠、水土流失严重的坡耕地逐步改造成为以人工林为主的半自然生境, 这对坝上地区的水土保持与水源涵养[4]、降低地质灾害、防止土地沙化[5]等做出了重要贡献。
生物多样性不但反映生态系统的弹性水平, 同样也维系着生态系统服务功能与人类福祉, 是评价生境质量的重要指标[6-8]。近年来, 人工林的现有建设模式对生物多样性的影响是学界的关注热点, 有学者认为, 人工林生境, 尤其是高密度的人工林生境, 不能保有较高的生物多样性水平, 这与人工林的较高种植密度与单一化林分的建设思路有关[9-10];也有学者认为, 人工林内较低的生物多样性水平与其建设过程较强的人为干扰有关, 而随着人为干扰逐渐降低与演替不断进行, 人工林的生物多样性水平会逐步提升[11]。退耕还林是我国人工林建设的一种重要工程模式, 但目前仍不清楚其生物多样性的现状、演替规律及环境影响因子。我国学者对退耕还林的相关研究主要集中在其水土保持[12-13]、社会经济效益[14-16]、营造技术模式[17-18]、土壤改善作用[19]等方面, 针对退耕还林生境的生物多样性水平的监测与评价仍较少。在退耕还林工程已实施了20周年后, 应该对坝上地区不同历史时期的退耕还林生境下的生物多样性水平进行监测与评价, 以更好了解退耕还林工程的生物多样性影响, 并从生物多样性保护角度提出退耕还林工程建设模式的改善建议。
草本层是森林生态系统的重要组分, 有重要生态价值。草本层植物生长周期较短, 对环境变量较敏感, 能较好体现生物群落对环境变化的响应, 并可为多类生物提供适宜生境与食物来源, 是衡量人工林生物多样性水平重要的生态指标[20]。通过选取20世纪70年代至21世纪初共计四个年代的退耕还林样地, 调查草本层植物多样性, 并与弃耕生境和自然林生境对比评价, 确定潜在土壤影响因子。为此, 本文提出三个假设:(1)不同年代的退耕还林生境的草本层植物多样性可能存在一定差异;(2)退耕还林生境的草本层植物多样性水平可能低于弃耕生境;(3)退耕还林生境的土壤理化性质及乔灌层多样性可能与草本层多样性有密切关系。
1 材料与方法 1.1 研究区域研究地点在河北省张家口市崇礼区(40°47′—41°17′N, 114°17′—115°34′E), 这里地处张家口东北部, 是连接内蒙古高原与华北平原的重要地点, 区内80%以上面积为山地;夏季均温19℃, 年均温约3.7℃;森林覆盖率约53%, 属典型的农牧交错带, 具有丰富的植物种质资源。
崇礼当地的人工造林工程主要发生在两个时期, 20世纪70—90年代为停耕造林工程, 21世纪初为退耕还林工程, 均为在坡耕地上种植人工林, 均以纯林为主, 主要栽植樟子松(Pinus sylvestris)与华北落叶松(Larix principis-rupprechtii), 林分密度较高;这两项工程在样地选择、建设模式、后期管护等方面均无显著差异。为论述方便, 下文统称退耕还林。
1.2 试验设计在崇礼区西湾子镇、白旗乡、红旗营镇与四台嘴乡, 共设计18个样地, 其中实验组为退耕还林生境, 跨越20世纪70年代至21世纪初的4个年代, 每个年代3个重复, 共计12个样地;对照组为弃耕生境及自然林生境, 每种生境3个重复, 共计6个样地。
为确保造林年限的准确性, 同当地林业部门、乡镇政府、当地村民多次核实, 并结合卫星历史图像进一步确认;其次, 不同样地之间保持超过1 km的距离, 同一年代重复样地的距离均超过5 km, 以避免受空间自相关性影响;样地筛选还要考虑坡向及坡度, 结合实际情况, 均为阴坡, 坡度介于6°—15°之间。样地海拔分布在1361.81—1712.80 m。生境具体情况详见表 1。
| 代码 Code |
生境类型 Habitat type |
样地数量 Plots No. |
植被描述 Description of vegetation |
| Q | 弃耕生境 | 3 | 没有乔木与灌木草本层: 茵陈蒿(Artemisia capillaris) + 山牛蒡(Synurus deltoides) + 委陵菜(Potentilla chinensis) +鼠掌老鹳草(Geranium sibiricum) +棉团铁线莲(Dendranthema chanetii) +小红菊+刺儿菜(Cirsium setosum) |
| T2000s | 退耕还林(2000年) | 3 | 乔木层以华北落叶松、樟子松为主要建群种。灌木层包括胡枝子(Lespedeza bicolor)、土庄绣线菊(Spiraea pubescens)、沙棘(Hippophae rhamnoides)、美蔷薇(Rosa bella)、锦鸡儿(Caragana sinica)、虎榛子(Ostryopsis davidiana) 等。草本层: 小红菊(Chrysanthemum chanetii) +披碱草(Elymus dahuricus) +硬质早熟禾(Poa sphondylodes) +细叶苔草(Carex rigescens) |
| T1990s | 退耕还林(20世纪90年代) | 3 | |
| T1980s | 退耕还林(20世纪80年代) | 3 | |
| T1970s | 退耕还林(20世纪70年代) | 3 | |
| N | 自然林 Natural forest | 3 | 天然混交林, 乔木层以白桦(Betula platyphylla)为建群种, 灌木层包括美蔷薇、土庄绣线菊、蒙古栎(Quercus mongolica)幼苗等, 半灌木包括悬钩子(Rosa rubus)等草本层: 龙芽草(Agrimonia pilosa) + 山牛蒡(Synurus deltoides) + 鸡腿堇菜(Viola acuminate) +歪头菜(Vicia unijuga) |
植物调查于2018年6月及9月进行, 其中, 乔灌木层植物调查在2018年6月进行, 草本层植物调查于2018年6月及9月进行。
在距离生境边缘不少于20 m的地点随机选取20 m×20 m的正方形样地, 分别测量乔木层与灌木层的植物名称、密度、平均高度、枝下高、胸径、冠幅盖度及乔木层总盖度(林分郁闭度), 并鉴定至物种水平;平均高度、枝下高、胸径测量和胸径高度通过皮尺直接测量;不同植物的盖度及总盖度通过估计植物冠层在地表的投影面积的百分比得出。
将样地分为4个10 m×10 m的网格, 并从每个网格中分别随机选取一个1 m×1 m的样方, 分别统计每个样方内草本层植物的名称、盖度与总盖度。
1.3.2 土壤调查土壤取样于2018年9月进行。采用直径50 mm的螺旋钻, 取地表以下20 cm土层内的土壤样品, 每个样地随机选取5点取样后混合均匀, 送至北京市农林科学院植保所测定总氮(Total nitrogen, TN)、总碳(Total carbon, TC)、有机质(Organic, OM)、土壤酸碱度(pH)和碱解氮(Alkali-hydrolyzale nitrogen, AHN), 并根据总氮和总碳含量计算土壤的碳氮比(C/N)。
1.4 数据分析本文所有数据分析均通过R 3.6.3实现。
首先计算了物种丰富度、盖度、Simpson多样性与均匀度指标, 用来指示α多样性的分布, 计算公式如下:
Simpson多样性:
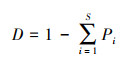
|
均匀度:
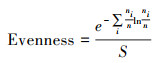
|
式中, S为物种丰富度, n为多度, ni为第i个物种的多度, Pi为第i个物种多度占总体多度的比例。以上指数均由vegan包计算得出。
利用单因素方差分析(One-way ANOVA)比较不同年代退耕还林生境的草本层植被α多样性差异[21], 利用shapiro法检验其正态性, 利用Bartlett法检验其方差齐性;当P>0.05时认为满足要求。当模型P < 0.05, 即不同组别存在显著性差异时, 利用Turkey HSD法进行多重比较[22]。此外, 采用单因素方差分析(One-way ANOVA)比较不同生境α多样性差异, 并采用Bonferroni法进行多重比较。以上分析通过Agricolae包及stats包进行[23]。
采用一般线性模型(general linear model, LM)结合模型筛选与平均(model selection and averaging) 的方法, 分析土壤及植被因子对退耕还林生境草本层植被多样性的影响。为避免出现共线性问题, 首先对环境与景观因子分别进行了相关性检验, 并剔除r2>0.49且P < 0.05的环境因子(见表 2)。在构建总体模型后, 对所有可能存在的子模型分别计算AICc值并进行排序, 最终将所有AICc < 4的模型进行模型平均(model averaging), 并计算不同环境因子的相对重要性(Relative importance)[19]。以上分析通过MuMIn包及stats包进行[24-25]。
| CS | RS | HS | DBH | CT | CTT | DT | HT | RT | TN | Org | TC | AHN | pH | |
| DS | 0.818** | 0.776** | 0.034 | 0.487 | -0.223 | -0.394 | -0.473 | 0.583* | -0.335 | 0.21 | 0.198 | 0.282 | -0.066 | 0.212 |
| CS | 0.833*** | 0.183 | 0.415 | -0.106 | -0.476 | -0.271 | 0.27 | -0.128 | 0.124 | 0.213 | 0.214 | 0.166 | 0.209 | |
| RS | 0.047 | 0.533 | -0.254 | -0.262 | -0.311 | 0.569 | 0 | 0.39 | 0.461 | 0.461 | -0.096 | -0.052 | ||
| HS | -0.037 | 0.157 | -0.031 | -0.033 | -0.143 | 0.129 | 0.277 | 0.245 | 0.214 | 0.133 | -0.234 | |||
| DBH | -0.595* | 0.004 | 0.085 | 0.537 | -0.05 | -0.076 | 0.095 | 0.084 | -0.085 | 0.104 | ||||
| CT | 0.291 | 0.355 | -0.224 | 0.377 | -0.065 | -0.077 | -0.17 | -0.288 | -0.225 | |||||
| CTT | 0.753** | 0.114 | 0.352 | 0.075 | 0.147 | 0.054 | -0.531 | -0.513 | ||||||
| DT | -0.264 | 0.388 | -0.442 | -0.267 | -0.414 | -0.36 | -0.134 | |||||||
| HT | -0.095 | 0.347 | 0.334 | 0.375 | -0.231 | -0.104 | ||||||||
| RT | 0.103 | 0.344 | 0.154 | -0.221 | -0.702* | |||||||||
| TN | 0.936*** | 0.973*** | 0.132 | -0.701* | ||||||||||
| Org | 0.975*** | 0.131 | -0.813** | |||||||||||
| TC | 0.182 | -0.718** | ||||||||||||
| AHN | 0.137 | |||||||||||||
| DS:灌木层密度 Shrub density;CS:灌木层总盖度 Total coverage of shrub layer;RS:灌木层丰富度 Shrub layer richness;HS:灌木层平均高度 Average height of shrub layer;DBH:乔木层胸径 Arbor breast diameter;CT:乔木层冠幅 Tree canopy width;CTT:乔木层总盖度 Tree layer total coverage;DT:乔木层密度 Tree layer density;HT:乔木层平均高度 Average height of arbor layer;RT:Tree layer richness;TN:全氮 Total nitrogen;Org:有机质Organic;TC:全碳 Total carbon;AHN:碱解氮 Alkali-hydrolyzale nitrogen; *:P < 0.05; **: P < 0.01; ***: P < 0.001 | ||||||||||||||
不同年代退耕还林生境的土壤及植被因子详见表 3。此外, 还测定了所有样地的土壤机械组成, 显示质地一致, 均为沙壤土。不同样地的土壤与植被因子相关性见表 2。
| 土壤与植被因子 | 退耕还林年代 Plantation decades of RFTF habitats | |||
| 1970s | 1980s | 1990s | 2000s | |
| 灌木层 Shrub layer | ||||
| 灌木层密度 Shrub density | 27.67 ± 18.717 | 21.67 ± 10.214 | 5.33 ± 4.509 | 8.33 ± 7.506 |
| 灌木层总盖度 Shrub coverage/% | 55 ± 21.794 | 40 ± 8.66 | 20 ± 18.028 | 28.33 ± 11.547 |
| 灌木层丰富度 Shrub richness | 4 ± 1 | 3 ± 1 | 2 ± 1.732 | 2 ± 0 |
| 灌木层平均高度 Shrub height/m | 1.022 ± 0.2037 | 1.086 ± 0.3118 | 1.267 ± 0.8737 | 1.1 ± 0.1 |
| 乔木层 Tree layer | ||||
| 乔木层胸径 Tree Arbor breast diameter/cm | 19.3333 ± 5.1316 | 11.6667 ± 2.88675 | 11.6667 ± 2.88675 | 10.058 ± 1.6835 |
| 乔木层冠幅 Tree coverage/m | 2.27 ± 0.643 | 2.6 ± 0.854 | 2.83 ± 0.961 | 2.07 ± 0.532 |
| 乔木层总盖度 Tree total coverage/% | 76.67 ± 15.275 | 70 ± 10 | 78.33 ± 2.887 | 78 ± 2.861 |
| 乔木层密度 Tree density | 36.33 ± 15.822 | 21.67 ± 2.887 | 35 ± 1 | 35 ± 8.66 |
| 乔木层平均高度 Tree height/m | 8.067 ± 1.1015 | 8.667 ± 4.1633 | 5 ± 1 | 5.1 ± 0.6557 |
| 乔木层丰富度 Tree richness | 1.33 ± 0.577 | 1 ± 0 | 1.33 ± 0.577 | 1.67 ± 0.577 |
| 土壤 Soil | ||||
| 全氮 Total Nitrogen/(g/kg) | 1.6433 ± 0.16442 | 2.6533 ± 1.58812 | 2.0567 ± 0.70784 | 1.94 ± 0.40951 |
| 有机质 Organic/(g/kg) | 25.1 ± 7.602 | 36.667 ± 29.4924 | 26.933 ± 17.9692 | 24.7 ± 13.1651 |
| 全碳 Total carbon/(g/kg) | 14.333 ± 3.5852 | 23.067 ± 16.3967 | 16.2 ± 7.9793 | 13.917 ± 5.2441 |
| 碱解氮 Alkali-hydrolyzale nitrogen/(mg/kg) | 113.33 ± 29.738 | 963.33 ± 671.563 | 183.67 ± 61.37 | 191.67 ± 80.928 |
| 土壤酸碱度 pH-value | 7.7767 ± 0.56889 | 7.6233 ± 0.72418 | 7.4533 ± 0.4373 | 7.39 ± 0.43486 |
方差分析显示, 不同年代退耕还林生境的草本层物种丰富度、总盖度、Simpson多样性指数及均匀度指数均差异不显著(表 4), 这说明, 本研究所选取的不同建设年代退耕还林生境, 其草本层植物多样性并未随建设年代的不同而呈现显著性差异(图 1)。
| 多样性指数 Diversity |
df | 平方和 Sum Sq |
平均平方和 Mean Sq |
F | P |
| 物种丰富度 Species richness | 3 | 77.58 | 25.86 | 2.483 | 0.135 |
| 总盖度 Total coverage | 3 | 373.9 | 124.6 | 1.205 | 0.368 |
| Simpson多样性指数 Simpson diversity index | 3 | 0.001289 | 0.0004297 | 0.995 | 0.443 |
| 均匀度 Evenness | 3 | 0.003591 | 0.001197 | 0.38 | 0.77 |

|
| 图 1 不同年代退耕还林生境草本层生物多样性 Fig. 1 Herb diversities in different plantation decades of returning farmland to forest (RFTF) habitats 相同字母表示组间差异不显著 |
方差分析显示, 不同生境类型的草本层物种丰富度、总盖度、Simpson多样性指数差异显著(表 5), 进一步的多重分析结果显示, 弃耕生境的物种丰富度、总盖度与Simpson多样性均显著高于其余两种生境, 而退耕还林生境的生物多样性则相对最低, 自然林介于弃耕生境与退耕还林生境之间(图 2)。
| 多样性指数 Diversity |
df | 平方和 Sum Sq |
平均平方和 Mean Sq |
F | P |
| 物种丰富度 Species richness | 2 | 148.9 | 74.46 | 4.174 | 0.0362 |
| 总盖度 Total coverage | 2 | 1066 | 532.9 | 6.593 | 0.00882 |
| Simpson多样性指数 Simpson diversity index | 2 | 0.004646 | 0.0023228 | 6.045 | 0.0119 |
| 均匀度 Evenness | 2 | 0.01494 | 0.007469 | 3.172 | 0.071 |

|
| 图 2 不同生境草本层植物多样性 Fig. 2 Herb diversities in different habitat types |
为排除共线性并确定最终纳入模型的土壤与植被变量, 首先对土壤及植被因子进行了相关分析, 具体结果见表 2。结合前文论述的共线性判定标准, 灌木层密度、灌木层丰富度、乔木层密度、碱解氮、土壤有机质含量等因子被纳入最终的模型。
分别对4个不同指标构建响应模型, 最终显示, 仅有物种丰富度与Simpson多样性指数对土壤及环境因子具有显著响应。其中, 物种丰富度模型共平均了4个AICc值介于57.3至60.52的线性模型, 平均模型中环境因子的相对重要性为:碱解氮>灌木层密度>灌木层丰富度>乔木层丰富度。最终的平均模型结果显示, 物种丰富度对灌木层密度、碱解氮及灌木层丰富度均有显著正响应, 而乔木层密度可能对物种丰富度具有潜在的负效应, 但这种趋势并不显著(表 6)。
| 环境因子 Environmental Factor |
AICc范围 AICc Range |
预估值 Estimate |
调整标准误 Adjusted SE |
Z | P | 相对重要性 Relative importance |
| 物种丰富度 Species richness | ||||||
| 灌木层密度 Shrub density |
[57.3, 60.52] | 0.157604 | 0.046227 | 3.409 | 0.000651 | 0.73 |
| AlW(mg/kg) | 0.004936 | 0.001169 | 4.222 | 2.42×10-5 | 1 | |
| 灌木层丰富度 Shrub richness |
1.447156 | 0.530991 | 2.725 | 0.006423 | 0.27 | |
| 乔木层密度 Tree density |
-0.112972 | 0.070796 | 1.596 | 0.110546 | 0.26 | |
| Simpson多样性 Simpson diversity | ||||||
| 乔木层密度 Tree density |
[-58.32, -54.64] | -1.335×10-3 | 5.752×10-4 | 2.322 | 0.0203 | 0.75 |
| AlW(mg/kg) | 1.973×10-5 | 1.049×10-5 | 1.880 | 0.0601 | 0.48 | |
| 乔木层平均高度 Tree height/ m |
-3.155×10-3 | 2.024×10-3 | 1.559 | 0.1190 | 0.18 | |
| 灌木层密度 Shrub density |
4.982×10-4 | 4.094×10-4 | 1.217 | 0.2237 | 0.05 | |
Simpson多样性模型共平均了6个AICc值介于-58.32与-54.64之间的线性模型。其中环境因子的相对重要性为乔木层密度>碱解氮>乔木层平均高度>灌木层密度。模型结果显示, 乔木层密度对草本层植物Simpson多样性呈显著负相关效应, 而碱解氮则与Simpson多样性指数存在潜在的正相关趋势(表 6)。
3 讨论退耕还林的林下草本层植物修复进程研究结果不一, 这可能与多种因素有关。澳大利亚的研究认为, 持续20—40年的退耕所形成的人工林的林下植物多样性未出现显著差异[26]。而Ahmada等在中国西北进行的研究显示, 林下植物多样性水平同人工林种植年限缺乏直接关系, 但可能存在以林分密度为中介变量的间接关系, 林下草本层的植物多样性指数可能随林分密度和林冠郁闭度增加而下降[27];崔宁洁等在四川低山丘陵区的研究显示, 5—39年生的马尾松人工林林下草本层物种多样性未发现显著差异[28];而罗洁贤等在广东省调查发现, 3年与5年生桉树林的林下草本层物种多样性出现了显著下降[29];在陕西黄土高原的研究表明, 不同林龄刺槐林的林下草本植物多样性指数总体呈现单峰效应, 在30年左右达到最大[30]。这个不一致首先可能与树种有关, 如Barbier等研究表明, 栎类与草本植物的关系较弱, 而松类与草本植物的关系可能较强[31-32];桉树为速生树种, 且具有较强化感作用, 对林下草本植物具有一定抑制性, 其动态变化可能快于预期。其次, 林下植被生物多样性随林龄变化的过程可能与多种因素有关, 林分主要通过调控土壤含水量、光照强度及凋落物与土壤菌群[33]等影响林下植被。第三, 这一过程可能与林地所处演替周期有关, 有学者提出人工林林下植被演替过程更有可能发生在幼林期, 在成熟林期很难产生显著性差异, 期间的改变则与环境条件有关[34]。对本研究而言, 退耕还林最低年限也在20年左右, 均已离开幼林期, 可能因此未发现林下草本层植物多样性的显著差异。最后, 很少学者关注林下草本层植物演替过程与其他生物的交互作用, 但大量研究都提示植被演替过程与授粉生物的关系密不可分[35-37]。本研究中林下草本多样性都普遍偏低, 这可能因林下植物很难得到有效授粉, 因此就很难形成草本层开花植物的有效迁移与交流。
本研究表明, 弃耕生境的草本层多样性水平较高, 这提示其自然恢复过程可能对恢复生物多样性有益。近年来, 弃耕生境作为一种特殊的恢复生境, 其生物多样性保护价值正日益得到重视。欧洲的研究表明, 弃耕生境是生物多样性的热点地区, 可有效提升区域范围内的生物多样性水平[38]。目前, 国内针对内蒙古鄂尔多斯、重庆巫山、河南豫西、陕西咸阳等不同区域的弃耕生境植被演替规律的研究结果显示, 弃耕生境在演替中能产生较高的草本植物多样性, 并随演替时间加强而逐步提升, 在进入灌丛与乔木群落阶段, 仍能保有较高的草本植物多样性水平[39-42]。这表明, 弃耕生境在全国范围内均可有益于保护生物多样性、促进生态系统服务功能。弃耕生境的生物多样性形成可能存在综合驱动因素, 如Lomba等对地中海山地农田弃耕的植物多样性影响的长期观测认为, 景观异质性是弃耕生境较高植物多样性的主要原因[43];也有研究认为, 弃耕地土壤的细菌群落动态与土壤理化性质是影响弃耕生境中植物多样性的主要因子之一[44]。此外, 弃耕生境恢复过程中人为干扰较少, 而人为干扰被认为是影响草本植物分布的重要因素[45], 在较低水平的干扰活动下, 土壤中可能保留更丰富的种子库, 且很少面临大型乔灌木的竞争作用, 因此更易保有较高的生物多样性[46]。
本文显示, 退耕还林生境的生物多样性对土壤及植被因子均有显著响应, 降低乔木层密度、增加灌木层密度及丰富度、提升土壤中的碱解氮水平, 都利于提升退耕还林生境的草本层植物多样性。草本的根系较浅, 更易受土壤理化性质影响, 因此0—20 cm土层的土壤养分与草本层物种多样性关系密切[47]。碱解氮是植物直接吸收利用的土壤营养, 人工林土壤中的碱解氮主要来源包括有机质凋落物的分解、人为活动, 固氮效应等, 低强度放牧可能导致土壤碱解氮含量提升[48]。本文显示土壤碱解氮含量是影响草本层植物多样性的重要潜在因子, 这与草本直接利用碱解氮促进其生长有关, 一些对水肥条件要求较高的草本植物也有可能更倾向于在碱解氮含量较高的生境分布。
林木密度对林下植被具有负面效应已渐成共识[49-50], 本文结果进一步佐证了这一结论。首先, 林木密度提高可能影响土壤理化性质(如全氮、含水量、有机质等), 从而进一步影响林下植物的养分来源[51];其次, 林木密度与林冠层郁闭度(覆盖度)呈显著正相关, 在华北落叶松林生境中, 林木密度高易造成松针累积覆盖, 并通过物理阻碍与化感作用影响植物萌发[52];第三, 林冠覆盖度高会降低林地总辐射, 从而改变林地植被组成[53-54]。
国内外对人工林林下植物多样性研究虽然较多[55-57], 但较少有研究涉及林下植物的交互作用, 尤其灌木层对草本层的影响。本文显示, 灌木层密度与丰富度是影响林下草本层植物多样性的关键要素, 草本层植物的物种丰富度对较高的灌木层密度与物种丰富度具有显著的正响应。灌木层对草本层植物的影响机制目前尚不明确, 但较丰富的林地垂直空间结构能容纳更高的生物多样性水平[58-59]。调查发现, 灌木层树种主要为乡土种, 其中包含了较多的豆科、蔷薇科植物, 较高的灌木层多样性可能受益于更合理完善的乔木冠层结构, 并有自身构成了林下灌木层, 还可能在一定程度上提高其他机制抵消乔木层的负面影响, 如调查中发现的土庄绣线菊、美蔷薇等是授粉生物喜爱的寄主植物与访花植物, 这对授粉生物构成溢出效应, 增加对林下草本层植物的访问, 这对草本层的构建具有积极影响[60]。
坝上地区即将开始迎来新一轮退耕还林建设, 其生物多样性保护与生态系统服务功能关乎区域生态效益, 需要未来进一步优化坝上地区的退耕还林工程模式, 以提高林下草本层植物多样性水平与森林的生物多样性保护功能。本文结果显示, 应尽量避免营造树种单一、结构单一的过密林分, 重视退耕还草及自然恢复的重要生态价值, 因地制宜, 宜林则林, 宜草则草, 同时关注恢复具有高生物多样性与高生态价值的草地。可适当降低退耕林密度, 适当补入乡土灌木, 营造结构完善的林分。在现有退耕林的生境管理上, 可考虑适当间伐, 降低林木密度, 补植适宜林下生长的乡土野花及其组合, 如北柴胡(Bupleurum chinense)、林地风毛菊(Saussurea hylophila)、驴欺口(Echinops davuricus)等, 进一步增强退耕还林后的生物多样性水平。
4 结论坝上地区不同年代退耕还林后的林下草本层植物多样性的差异不显著, 且均低于弃耕地生境与自然林生境;林下草本层植物多样性对乔灌层植被特征及土壤特征均有显著响应, 林木密度是制约林下草本层植物多样性的关键因子, 增强对林下灌木层的塑造与维护和增加土壤碱解氮含量以利于提升林下草本层的物种多样性。在未来国土绿化及生态修复中, 需因地制宜, 重视自然恢复, 积极改善人工林建设模式, 优化林分空间结构, 以满足多样化的生态系统服务功能需求。
| [1] |
王彦芳, 裴宏伟. 1980-2015年河北坝上地区生态环境状况评价与对策研究. 生态经济, 2018, 34(1): 186-190, 236-236. |
| [2] |
许团斌. 柳河县停耕造林工作体系机制探讨. 绿色科技, 2014(8): 98-99. DOI:10.3969/j.issn.1674-9944.2014.08.041 |
| [3] |
李世东. 中外退耕还林还草之比较及其启示. 世界林业研究, 2002, 15(2): 22-27. DOI:10.3969/j.issn.1001-4241.2002.02.004 |
| [4] |
杨波, 王全九. 退耕还林后榆林市土壤侵蚀和养分流失功效研究. 水土保持学报, 2016, 30(4): 57-63. |
| [5] |
李建明, 王志刚, 王爱娟, 王家乐, 王可, 刘晨曦, 崔豪, 张平仓. 退耕还林恢复年限对岩溶槽谷区石漠化土壤物理性质的影响. 农业工程学报, 2020, 36(1): 99-108. |
| [6] |
Bennett E M, Cramer W, Begossi A, Cundill G, Díaz S, Egoh B N, Geijzendorffer I R, Krug C B, Lavorel S, Lazos E, Lebel L, Martín-López B, Meyfroidt P, Mooney H A, Nel J L, Pascual U, Payet K, Harguindeguy N, Peterson G D, Prieur-Richard A H, Reyers B, Roebeling P, Seppelt R, Solan M, Tschakert P, Tscharntke T, Turner Ⅱ B L, Verburg P H, Viglizzo E F, White P C, Woodward G. Linking biodiversity, ecosystem services, and human well-being: three challenges for designing research for sustainability. Current Opinion in Environmental Sustainability, 2015, 14: 76-85. DOI:10.1016/j.cosust.2015.03.007 |
| [7] |
Mace G M, Norris K, Fitter A H. Biodiversity and ecosystem services: a multilayered relationship. Trends in Ecology & Evolution, 2012, 27(1): 19-26. |
| [8] |
Tscharntke T, Klein A M, Kruess A, Steffan-Dewenter I, Thies C. Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecology Letters, 2005, 8(8): 857-874. DOI:10.1111/j.1461-0248.2005.00782.x |
| [9] |
Horák J, Brestovanská T, Mladenović S, Kout J, Bogusch P, Halda J P, Zasadil P. Green desert?: biodiversity patterns in forest plantations. Forest Ecology and Management, 2019, 433: 343-348. DOI:10.1016/j.foreco.2018.11.019 |
| [10] |
Moen J, Gardfjell H, Oksanen L, Ericson L, Ekerholm P. Grazing by food-limited microtine rodents on a productive experimental plant community: does the "green desert" exist?. Oikos, 1993, 68(3): 401-413. DOI:10.2307/3544908 |
| [11] |
Mansourian S, Vallauri D. Forest Restoration in Landscapes: Beyond Planting Trees. New York: Springer Science & Business Media, 2005.
|
| [12] |
杨光, 丁国栋, 赵廷宁, 孙保平. 黄土丘陵沟壑区退耕还林的水土保持效益研究——以陕西省吴旗县为例. 水土保持通报, 2006, 26(2): 88-90, 99-99. DOI:10.3969/j.issn.1000-288X.2006.02.022 |
| [13] |
罗海波, 钱晓刚, 刘方, 何腾兵, 宋光煜. 喀斯特山区退耕还林(草)保持水土生态效益研究. 水土保持学报, 2003, 17(4): 31-34, 41-41. DOI:10.3321/j.issn:1009-2242.2003.04.008 |
| [14] |
王小龙. 退耕还林: 私人承包与政府规制. 经济研究, 2004(4): 107-116. |
| [15] |
李蕾, 刘黎明, 唐伟. 退耕还林还草对农民收入及农村经济的影响. 农村经济, 2004(3): 50-51. |
| [16] |
黄富祥, 康慕谊, 张新时. 退耕还林还草过程中的经济补偿问题探讨. 生态学报, 2002, 22(4): 471-478. |
| [17] |
王继军, 谢永生, 卢宗凡, 权松安. 退耕还林还草下生态农业发展模式初探. 水土保持学报, 2004, 18(1): 134-137. DOI:10.3321/j.issn:1009-2242.2004.01.035 |
| [18] |
李世东, 刘霞. 东北山地沙地区和京津周围沙地平原区退耕还林区域模式研究. 防护林科技, 2004(2): 1-6, 12-12. DOI:10.3969/j.issn.1005-5215.2004.02.001 |
| [19] |
鹿士杨, 彭晚霞, 宋同清, 曾馥平, 杜虎, 王克林. 喀斯特峰丛洼地不同退耕还林还草模式的土壤微生物特性. 生态学报, 2012, 32(8): 2390-2399. |
| [20] |
Vockenhuber E A, Scherber C, Langenbruch C, Meißner M, Seidel D, Tscharntke T. Tree diversity and environmental context predict herb species richness and cover in Germany's largest connected deciduous forest. Perspectives in Plant Ecology, Evolution and Systematics, 2011, 13(2): 111-119. DOI:10.1016/j.ppees.2011.02.004 |
| [21] |
Chambers J M, Freeny A E, Heiberger R M. Analysis of variance; designed experiments//Chambers J M, Hastie T J, eds. Statistical Models in S. Pacific Grove, California: Wadsworth & Brooks/Cole, 2017: 145-193.
|
| [22] |
Hsu J. Multiple Comparisons: Theory and Methods. New York: CRC Press, 1996.
|
| [23] |
de Mendiburu F, de Mendiburu M F. Package 'Agricolae'. R Package Version, 2019, 1-2. |
| [24] |
Carvajal-Rodríguez A. Multi-model inference of non-random mating from an information theoretic approach. Theoretical Population Biology, 2020, 131: 38-53. DOI:10.1016/j.tpb.2019.11.002 |
| [25] |
Barton K, Barton M K. Package 'MuMIn'. R Package Version, 2019, 1(6). |
| [26] |
Wei L P, Archaux F, Hulin F, Bilger I, Gosselin F. Stand attributes or soil micro-environment exert greater influence than management type on understory plant diversity in even-aged oak high forests. Forest Ecology and Management, 2020, 460: 117897. DOI:10.1016/j.foreco.2020.117897 |
| [27] |
Ahmad B, Wang Y H, Hao J, Liu Y H, Bohnett E, Zhang K B. Optimizing stand structure for trade-offs between overstory timber production and understory plant diversity: a case-study of a larch plantation in Northwest China. Land Degradation & Development, 2018, 29(9): 2998-3008. |
| [28] |
崔宁洁, 张丹桔, 刘洋, 张健, 欧江, 张捷, 邓超, 纪托未. 不同林龄马尾松人工林林下植物多样性与土壤理化性质. 生态学杂志, 2014, 33(10): 2610-2617. |
| [29] |
罗洁贤, 俞政民, 聂钰滢, 吕培, 邱权. 不同林龄桉树林下植被多样性分析. 林业与环境科学, 2019, 35(1): 36-42. |
| [30] |
杨丽霞, 任广鑫, 韩新辉, 康迪, 赵发珠, 邓健, 杨改河. 黄土高原退耕区不同林龄刺槐林下草木植物的多样性. 西北农业学报, 2014, 23(7): 172-178. |
| [31] |
Barbier S, Chevalier R, Loussot P, Bergès L, Gosselin F. Improving biodiversity indicators of sustainable forest management: tree genus abundance rather than tree genus richness and dominance for understory vegetation in French lowland oak hornbeam forests. Forest Ecology and Management, 2009, 258(S1): S176-S186. |
| [32] |
Barbier S, Gosselin F, Balandier P. Influence of tree species on understory vegetation diversity and mechanisms involved-a critical review for temperate and boreal forests. Forest Ecology and Management, 2008, 254(1): 1-15. DOI:10.1016/j.foreco.2007.09.038 |
| [33] |
Santonja M, Rancon A, Fromin N, Baldy V, Hättenschwiler S, Fernandez C, Montès N, Mirleau P. Plant litter diversity increases microbial abundance, fungal diversity, and carbon and nitrogen cycling in a Mediterranean shrubland. Soil Biology and Biochemistry, 2017, 111: 124-134. DOI:10.1016/j.soilbio.2017.04.006 |
| [34] |
Swanson M E, Franklin J F, Beschta R L, Crisafulli C M, DellaSala D A, Hutto R L, Lindenmayer D B, Swanson F J. The forgotten stage of forest succession: early-successional ecosystems on forest sites. Frontiers in Ecology and the Environment, 2011, 9(2): 117-125. DOI:10.1890/090157 |
| [35] |
Coulin C, Aizen M A, Garibaldi L A. Contrasting responses of plants and pollinators to woodland disturbance. Austral Ecology, 2019, 44(6): 1040-1051. DOI:10.1111/aec.12771 |
| [36] |
Joshi N K, Otieno M, Rajotte E G, Fleischer S J, Biddinger D J. Proximity to woodland and landscape structure drives pollinator visitation in apple orchard ecosystem. Frontiers in Ecology and Evolution, 2016, 4: 38. |
| [37] |
Williams N M, Winfree R. Local habitat characteristics but not landscape urbanization drive pollinator visitation and native plant pollination in forest remnants. Biological Conservation, 2013, 160: 10-18. DOI:10.1016/j.biocon.2012.12.035 |
| [38] |
Queiroz C, Beilin R, Folke C, Lindborg R. Farmland abandonment: threat or opportunity for biodiversity conservation? A global review. Frontiers in Ecology and the Environment, 2014, 12(5): 288-296. DOI:10.1890/120348 |
| [39] |
石丹, 倪九派, 倪呈圣, 刘锦春. 巫山高山移民迁出区不同弃耕年限对植物物种多样性的影响. 生态学报, 2019, 39(15): 5584-5593. |
| [40] |
赵威, 李亚鸽, 亓琳, 李琳, 王馨. 豫西丘陵坡地弃耕农田植被演替对土壤碳、氮库的影响. 生态学报, 2018, 38(19): 7016-7025. |
| [41] |
张瑞红, 蔡文涛, 来利明, 周继华, 王永吉, 姜联合, 郑元润. 鄂尔多斯高原弃耕农田植物群落演替过程中物种多样性的变化. 干旱区资源与环境, 2018, 32(10): 178-183. |
| [42] |
郝红敏, 刘玉, 王冬, 杨政, 武高林. 典型草原开垦弃耕后不同年限群落植物多样性和空间结构特征. 草地学报, 2016, 24(4): 754-759. |
| [43] |
Lomba A, Gonçalves J, Moreira F, Honrado J. Simulating long-term effects of abandonment on plant diversity in Mediterranean mountain farmland. Plant Biosystems-An International Journal Dealing with all Aspects of Plant Biology, 2013, 147(2): 328-342. DOI:10.1080/11263504.2012.716794 |
| [44] |
Zhang C, Liu G B, Xue S, Wang G L. Soil bacterial community dynamics reflect changes in plant community and soil properties during the secondary succession of abandoned farmland in the Loess Plateau. Soil Biology and Biochemistry, 2016, 97: 40-49. DOI:10.1016/j.soilbio.2016.02.013 |
| [45] |
Hobbs R J, Huenneke L F. Disturbance, diversity, and invasion: implications for conservation. Conservation Biology, 1992, 6(3): 324-337. DOI:10.1046/j.1523-1739.1992.06030324.x |
| [46] |
Bakker J P, Poschlod P, Strykstra R J, Bekker R M, Thompson K. Seed banks and seed dispersal: important topics in restoration ecology. Acta Botanica Neerlandica, 1996, 45(4): 461-490. DOI:10.1111/j.1438-8677.1996.tb00806.x |
| [47] |
Brunet J, Falkengren-Grerup U, Tyler G. Herb layer vegetation of south Swedish beech and oak forests-effects of management and soil acidity during one decade. Forest Ecology and Management, 1996, 88(3): 259-272. DOI:10.1016/S0378-1127(96)03845-5 |
| [48] |
杨立宾, 朱道光, 崔福星, 等. 寒温带兴安落叶松林不同林型土壤微生物群落特征. 东北林业大学学报, 2017, 045(9): 66-72. DOI:10.3969/j.issn.1000-5382.2017.09.014 |
| [49] |
孙千惠, 吴霞, 王媚臻, 等. 林分密度对马尾松林林下物种多样性和土壤理化性质的影响. 应用生态学报, 2018, 29(3): 732-738. |
| [50] |
Chen Y, Cao Y. Response of tree regeneration and understory plant species diversity to stand density in mature Pinus tabulaeformis plantations in the hilly area of the Loess Plateau, China]. Ecological engineering, 2014, 73: 238-245. DOI:10.1016/j.ecoleng.2014.09.055 |
| [51] |
Paoli G D, Curran L M, Slik J W F. Soil nutrients affect spatial patterns of aboveground biomass and emergent tree density in southwestern Borneo. Oecologia, 2008, 155(2): 287-299. DOI:10.1007/s00442-007-0906-9 |
| [52] |
韩芬, 王辉, 边银霞, 李永兵. 华北落叶松枝叶挥发性物质的化学成分及其化感作用. 应用生态学报, 2008, 19(11): 2327-2332. |
| [53] |
Tripathi S, Bhadouria R, Srivastava P, Devi R S, Chaturvedi R, Raghubanshi A S. Effects of light availability on leaf attributes and seedling growth of four tree species in tropical dry forest. Ecological Processes, 2020, 9(1): 2. DOI:10.1186/s13717-019-0206-4 |
| [54] |
Crous-Duran J, Graves A R, Paulo J A, Mirck J, Oliveira T S, Kay S, de Jalón S G, Palma J H N. Modelling tree density effects on provisioning ecosystem services in Europe. Agroforestry Systems, 2019, 93(5): 1985-2007. DOI:10.1007/s10457-018-0297-4 |
| [55] |
贾亚运, 周丽丽, 吴鹏飞, 刘雨晖, 马祥庆. 不同发育阶段杉木人工林林下植被的多样性. 森林与环境学报, 2016, 36(1): 36-41. |
| [56] |
冯颖, 赵媛媛, 郭跃, 杨育红, 李文臣, 马成功, 丁国栋. 不同经营模式对华北落叶松人工林林下植被的影响. 水土保持通报, 2015, 35(2): 64-69, 75-75. |
| [57] |
成向荣, 徐金良, 刘佳, 虞木奎. 间伐对杉木人工林林下植被多样性及其营养元素现存量影响. 生态环境学报, 2014, 23(1): 30-34. DOI:10.3969/j.issn.1674-5906.2014.01.005 |
| [58] |
Machado M A, de Almeida Jr E B. Spatial structure, diversity, and edaphic factors of an area of Amazonian coast vegetation in brazil. The Journal of the Torrey Botanical Society, 2019, 146(1): 58-68. DOI:10.3159/TORREY-D-18-00025.1 |
| [59] |
高保嘉, 张执中. 封山育林对植物群落结构及多样性的影响. 北京林业大学学报, 1992, 014(2): 46-53. |
| [60] |
Isbell F, Adler P R, Eisenhauer N, Fornara D, Kimmel K, Kremen C, Letourneau D K, Liebman M, Polley H W, Quijas S, Scherer-Lorenzen M. Benefits of increasing plant diversity in sustainable agroecosystems. Journal of Ecology, 2017, 105(4): 871-879. DOI:10.1111/1365-2745.12789 |