 2021, Vol. 41
2021, Vol. 41文章信息
- 郝建锋, 余飞燕, 姚小兰, 陈聪琳, 喻静, 向琳, 齐锦秋
- HAO Jianfeng, YU Feiyan, YAO Xiaolan, CHEN Conglin, YU Jing, XIANG Lin, QI Jinqiu
- 放猪干扰对四川夹金山植物群落特征和生物量的影响
- Effects of grazing disturbance on vegetation community characteristics and biomass in Jiajin Mountain, Sichuan Province
- 生态学报. 2021, 41(24): 9645-9654
- Acta Ecologica Sinica. 2021, 41(24): 9645-9654
- http://dx.doi.org/10.5846/stxb202104241083
-
文章历史
- 收稿日期: 2021-04-24





2. 四川省水土保持与荒漠化防治重点实验室, 成都 611130;
3. 海南大学生态与环境学院, 环南海陆域生物多样性研究中心, 海口 570228;
4. 海南省热带生态环境修复工程研究中心, 海口 570228;
5. 四川省木材工业与家具工程重点实验室, 成都 611130
2. Key Laboratory for Soil and Water Conservation and Desertification Control of Sichuan, Chengdu 611130, China;
3. Center for Terrestrial Biodiversity of the South China Sea, College of Ecology and Environment, Hainan University, Haikou 570228, China;
4. Center for Eco-Environmental Restoration Engineering of Hainan Province, Haikou 570228, China;
5. Key Laboratory of wood industry and furniture engineering of Sichuan, Chengdu 611130, China
放牧压力从牲畜聚落中心到边缘在空间上是不均衡分布的[1]。随放牧干扰梯度的变化, 等补偿、超补偿和欠补偿效应都可能出现, 其中在适度的放牧干扰下, 牲畜的翻拱践踏可促进空间上的均匀化养分循环[2-3], 且更利于当地植物向耐牧型进化。但在过度放牧影响下, 植被构型格局、生物量分配比值、植被功能群和生活型等均会发生改变[4-6], 造成森林生态系统局部区域过多暴露在外界。目前相关研究主要通过人为划分干扰梯度覆盖范围[7]、测量距饲养场远近程度[8]、根据生物量计算理论承载牛羊数[9]、佩戴式定位器追踪牲畜轨迹[1]等方式来量化放牧干扰梯度研究。二代野猪是纯种野猪与家猪的后代, 食性杂, 通常在土壤中掘根取食植物种子、根茎、脊椎动物和无脊椎动物[10], 时有人为补喂玉米, 多栖身于林下与灌丛, 采食速度更快、时间更长。此类动物性情凶猛, 活动范围更广, 对植被的破坏性更强, 在国内国外研究中的放牧对象主要包括羊、牛、马等或者混合放牧, 涉及二代野猪放牧的研究较少。
物种多样性是衡量一个群落内物种分布的数量和均匀程度的综合指标[7]。生物量是森林生态系统结构优劣和功能高低的最直接的表现, 植物各部分生物量分配格局反映了植物对放牧干扰的生态适应对策[11]。有关物种多样性与生物量之间的关系及其作用机制的研究是植被生态学领域的研究热点, 物种多样性水平的高低与植物群落地上地下生物量分配密切相关, 许多前人研究结果显示物种多样性水平与生物量间呈正相关[12-13]。但薛睿等[2]发现, 在森林群落林窗及强烈的非对称性竞争中, 物种多样性与地上生物量更可能呈现负相关关系。在放牧压力下, 优势种优势度降低, 杂类草比例的增加会导致生境异质性增加, 促进物种多样性指数升高, 而生产力总体下降[6], 因此也会出现负相关情况。相关研究多见于高寒高原草甸[14-15]、荒漠草原[16]等生态系统, 且大多探讨当地植被特征变化对气候变化的响应[17-18], 有关夹金山高山山地的植被研究极少。
作为中国西部生态屏障建设的重要环节, 夹金山针阔混交林内的二代野猪放牧在近年来兴起, 但夹金山植物群落生长受到严重啃食、翻拱、踩踏。因此本文在四川夹金山二代野猪不同干扰强度的研究区开展试验研究, 旨在探讨: (1)夹金山植物群落总体及各功能群(一年生禾草、多年生禾草、一年生杂类草、多年生杂类草及多年生豆科)物种多样性水平;(2)不同植被层次的物种多样性与地上地下生物量的关系;(3)不同植被层次的物种多样性与地上地下生物量对不同等级放猪干扰的响应。最终为有效评估高山地植物群落在空间尺度上的植物多样性变化和建立合理的放牧制度提供科学依据。
1 材料与方法 1.1 研究区域概况夹金山(102°24′56″—102°38′38″E, 30°38′14″—30°54′22″N)位于宝兴县西北、阿坝州懋功县(现小金县)以南, 为邛崃山脉南部高山, 是省级风景名胜区、红军长征文化遗迹地、国家级森林公园。因受高空西风环流和印度洋西南季风的影响, 明显具有高原型季风气候, 海拔4140 m, 年降水量776.5 mm, 年日照时数1400 h[19]。夹金山土壤类型有山地黄壤、山地黄棕壤、山地暗棕壤、亚高山草甸土等。
夹金山有丰富的野生动物资源, 如金丝猴(Rhinopithecus roxellana)、马鹿(Cervus elaphus)等国家级野生保护动物。夹金山森林植被占53%, 亚高山与高山灌丛草甸植被占36%, 裸岩与雪线以上的冰雪区占8%, 农业用地占4%[20]。经实地踏查, 研究区内植物以川滇长尾槭、冷杉(Abies fabri)、箭竹、秀丽莓(Rubus amabilis)、黄花鼠尾草(Salvia flava)等为主。
1.2 样地设置在全面踏查的基础上, 根据二代野猪活动情况, 牧道数量、面积以及植被破坏程度[21], 结合王晓[22]对山地放牧干扰程度的划分, 分别设置二代野猪干扰区和对照区。经实地踏查发现, 以养猪场为中心, 沿河逆流而上(海拔起伏不大, 表 1)而干扰程度逐步降低, 因此在针阔混交林内每隔1000 m左右按照与河流平行的方向划分样地, 用“Ⅰ—Ⅳ级”干扰程度表示干扰程度逐步降低, 每个类型在沿着同一方向划分4个20 m×30 m的样地, 同时选取立地条件基本一致, 未受到野猪干扰的样地作为对照(CK), 能够反映该区域的长期放牧效应, 总计20个样地(表 1)。
| 于扰强度 Diturbance intensity |
样地 Plot |
经纬度 Longtitude and latitude |
坡度 Slope/(°) |
坡向 Aspect/(°) |
海拔 Altitude/m |
样地情况 Plot situation/% |
| I | p1—p4 | 102°42′45.35″N, 30°51′43.60″E |
24.5 | NE53 | 2908 | 约110头野猪(幼猪50头左右)几乎全天性在该范围内活动, 牧道数量及面积较大, 乔木生长受抑, 灌木草本啃食现象严重 |
| II | p5—p8 | 102°42′55.06″N, 30°52′12.82″E |
24 | NE61 | 2987 | 约70头野猪(幼猪10头左右), 集群活动开始时间为上午九点左右, 牧道数量和面积相对较少, 灌木草本植物遭啃食 |
| III | p9—p12 | 102°42′37.74″N, 30°52′41.65″E |
26 | NE71 | 3007 | 约50头野猪, 集群活动开始时间基本在中午, 牧道数量少, 面积小, 灌木草本植物受破坏小 |
| IV | p13—16 | 102°42′24.59″N, 30°53′15.41″E |
22 | NE65 | 3018 | 大概40头野猪下午时间在此范围内活动, 干扰频次小, 部分植物得以正常生长 |
| CK | p17—p20 | 102°42′30.68″N, 30°53′45.16″E |
25 | NE63 | 3021 | 无野猪放牧干扰 |
| I: 重度干扰Heavy disturbance;Ⅱ: 较重度干扰Little heavy disturbance;Ⅲ: 中度干扰Middle disturbance;Ⅳ: 轻度干扰Slight disturbance;CK: 对照区Control area;NE: 东北Northeast | ||||||
采用“相邻格子法”将每个样地等分为6个10 m×10 m的乔木样方, 并在每块样地中沿对角线选取6个5 m×5 m的灌木样方和12个1 m×1 m的草本样方进行调查。分别记录所有树高≥3 m的乔木树种名称、数量、高度、冠幅和胸径, 以及灌木物种(树高<3 m的木本个体)和草本植物的物种名称、数量、高度和盖度, 共计样方480个, 其中乔木层、灌木层、草本层样方总数分别为120个、120个和240个。同时使用全球定位系统(GPS, GARMIN公司生产)记录经度、纬度、海拔, 运用DQL-1B型森林罗盘仪(哈尔滨光学仪器厂)测定坡度、坡向。
2.2 生物量测定在每个样地中, 随机选取3个点对枯落物量进行收集, 同时随机选取3个草本样方, 采取“全株收获法”, 带回实验室后将其地上、地下部分剪开(茎基部以上为地上部分, 其余部分为地下部分), 同时洗去地下部分所带的残余土渣, 然后各装入相应编号的纸袋, 并置于65℃烘箱中烘干至恒重[23], 记录好各部分生物量;灌木生物量则参照杨昆和管东生[24]的模型建立方法, 在各类型每个样地内选取样木, 测定基径、株高、盖度和冠幅直径等指标, 选用与林下灌木生物量相关性较好的冠幅直径×株高作为回归模型的自变量, 对其进行拟合, 再选择最优模型, 计算其地上生物量、地下生物量, 得到整个样地林下灌木生物量。
2.3 数据计算 2.3.1 物种多样性计算本文选取4个多样性指数来分析夹金山群落物种多样性水平[25], 相对密度=某种的株数/所有种的总株数;相对频度=某种在样方中出现的次数/所有种出现的总次数;相对盖度=某种的盖度/所有种盖度之和;灌木层、草本层重要值: IV=(相对密度+相对盖度+相对频度)/3
丰富度指数: D=S
Simpson指数(优势度指数): 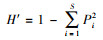
Shannon-Wiener多样性指数: 
Pielou均匀度指数: 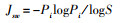
式中, Pi为第i种的株数(ni)占所有种总株数(n)的比例, 即Pi=ni/n(i=1, 2, 3, …… S, S为物种数)。
2.3.2 林下更新能力本研究根据郭其强等[26]研究选取指标, 参照高润梅等[27]对优势种幼苗幼树的划分, 即胸径胸径(DBH) < 2.5 cm的个体为幼苗, 2.5 cm≤DBH < 7.5 cm的为幼树;再统计其比值, 以各类型平均值作为评价群落更新潜力的指标。
2.3.3 植物群落物种多样性和生物量的RDA分析先用species-sample数据资料做DCA分析得到分析结果, Lengths of gradient的第一轴大小值为0.3, 因此适于RDA分析。选用各层次生物量作响应变量, 物种多样性作为解释变量, 对5个干扰等级的20个样地进行RDA二维分析排序。
2.4 数据处理采用Excel 2016和DPS 19.0对数据进行统计处理。采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)研究不同放猪梯度群落物种多样性和生物量差异(P=0.05)。采用Origin 9.0、Excel 2016和Canoco 5.0软件制图。
3 结果和分析 3.1 物种组成夹金山针阔混交林群落共有植物55科117属172种, 其中乔木层9科10属13种、灌木层20科27属57种、草本层38科89属128种。据图 1, 乔木层以槭树科(Aceraceae)、松科(Pinaceae)等为主, 且乔木层科、属、种数随干扰等级变化未明显波动, 在Ⅳ级干扰下物种组成最丰富(8科9属12种)。灌木层以蔷薇科(Rosaceae)、虎耳草科(Saxifragaceae)等为主, 在I级干扰下植被种类最少(11科12属15种), 在受Ⅳ级干扰下物种组成最丰富(15科20属37种)。草本层以唇形科(Lamiaceae)、紫草科(Boraginaceae)等为主, Ⅳ级干扰下物种组成丰富程度(31科57属77种)略高于对照(29科46属70种), 物种组成变化趋势与灌木层一致, 即随着干扰梯度增加而减小。乔灌草三层均在Ⅳ级干扰下物种多样性水平最高, I级干扰下物种数最贫乏。

|
| 图 1 不同放牧干扰下夹金山针阔混交林群落物种组成 Fig. 1 The species composition of coniferous-broad-leaved mixed forest under different grazing disturbance levels in Jiajin Mountain T: 乔木层Tree layer;S: 灌木层Shrub layer;H: 草本层Herb layer;I: 重度干扰Heavy disturbance;Ⅱ: 较重度干扰Little heavy disturbance;Ⅲ: 中度干扰Middle disturbance;Ⅳ: 轻度干扰Slight disturbance;CK: 对照区Control area |
不同程度放牧干扰下, 夹金山针阔混交林群落各层次优势种不同(表 2)。乔木层、灌木层在各程度干扰下优势种变动较小。随着干扰程度递减, 乔木层中冷杉逐渐占据研究区首要地位(从0.165逐渐上升到0.245), 川滇长尾槭则几乎未受影响(重要值始终高居0.211—0.327之间), 抗放牧干扰能力最强。就灌木层而言, 箭竹重要值明显大于其他物种而成为群落优势种, 秀丽莓的重要值在干扰梯度增加的过程中从0.092跃升为0.270。且结合对照(CK)来看, 夹金山灌木层中蔷薇科贡献了较高重要值(0.290—0.378)。草本层优势种波动较大, 在不同干扰梯度下均有不同物种组成, 黄花鼠尾草、酸模(Rumex acetosa)在放牧干扰程度加大的过程中分别由0.136、0.144剧减为0.035、0.004, 抗干扰能力差。各干扰梯度下均有多枝、蔓生或攀缘状草本植物出现(重要值≥0.029), 如原拉拉藤(Galium aparine)、茜草(Rubia cordifolia)等。
| 层次 layer |
干扰梯度 Disturbance intensity |
优势种 Dominant species |
重要值之和 Total important value |
| 乔木层 Tree layer |
I | 川滇长尾槭+糙皮桦+冷杉+山梅花+川西樱桃 Acer caudatum var. prattii+Betula utilis+Abies fabri+Philadelphus incanus+Cerasus trichostoma |
0.972 |
| Ⅱ | 川滇长尾槭+冷杉+山梅花+糙皮桦+花楸 Acer caudlatum var. prattii+Abies fabriPhiladelphus incanus+Betula utilis+Sorbus pohuashanensis |
0.873 | |
| Ⅲ | 川滇长尾槭+冷杉+糙皮桦+山梅花+红桦 Acer caudlatum var. prattii+Abies fabri+Betula utilis+Philadelphus incanus+Betula albosinensis |
0.897 | |
| Ⅳ | 冷杉+川滇长尾槭+糙皮桦+山梅花+红桦 Abies fabri+Acer caudatum var. prattii+Betula utilis+Philadelphus incanus+Betula albosinensis |
0.790 | |
| CK | 川滇长尾槭+冷杉+糙皮桦+花楸+山梅花 Acer caudatum var. prattii+Abies fabri+Betula utilis+Sorbus pohuashanensis+Philadelphus incanus |
0.783 | |
| 灌木层 Shrub layer |
I | 秀丽莓+箭竹+唐古特忍冬+高粱泡+猫儿刺 Rubus amabilis+Fargesia spathacea+Lonicera szechuanica+Rubus lambertianus+Ilex pernyi |
0.710 |
| Ⅱ | 箭竹+秀丽莓+山梅花+唐古特忍冬+花楸 Fargesia spathacea+Rubus amabilis+Philadelphus incanus+ Lonicera szechuanica+Sorbus pohuashanensis |
0.803 | |
| Ⅲ | 箭竹+插田泡+川滇长尾槭+秀丽莓+山梅花 Fargesia spathacea+Rubus coreanus+Acer caudatum var. prattii+Philadelphus incanus+Acer caudatum var. prattii |
0.666 | |
| Ⅳ | 箭竹+插田泡+川滇长尾槭+秀丽莓+山梅花 Fargesia spathacea+Rubus amabilis+Acer caudatum var. prattii+Rubus amabilis+Philadelphus incanus |
0.576 | |
| CK | 箭竹+插田泡+秀丽莓+山梅花+糙皮桦 Fargesia spathacea+Rubus coreanus+Rubus amabilis+Philadelphus incanus+Betula utilis |
0.660 | |
| 草本层 Herb layer |
I | 宽翅香青+蕨+碎米莎草+夜香牛+茜草 Anaphalis latialata+Pteridium aquilinum var. latiusculum+Cyperus iria+Vernonia cinerea+Rubia cordifolia |
0.338 |
| Ⅱ | 琉璃草+黄花鼠尾草+酸模+西南草莓+路边青 Cynoglossum zeylanicum+Salvia flava+Rumex acetosa+Fragaria moupinensis+Geum aleppicum |
0.516 | |
| Ⅲ | 黄花鼠尾草+蟹甲草+原拉拉藤+酸模+琉璃草 Salvia flava+Parasenecio forrestii+Galium aparine+Rumex acetosa+Cynoglossum zeylanicum |
0.427 | |
| Ⅳ | 酸模+黄花鼠尾草+琉璃草+蟹甲草+原拉拉藤 Rumex acetosa+Salvia flava+Cynoglossum zeylanicum+Parasenecio forrestii+Galium aparine |
0.429 | |
| CK | 柳叶菜+蟹甲草+阴地苎麻+原拉拉藤+暗花金挖耳 Epilobium hirsutum+Parasenecio forrestii+Boehmeria umbrosa+Galium aparine+Carpesium triste |
0.367 |
不同生活型功能群对不同干扰梯度响应不同(图 2)。一年生杂类草在各干扰梯度下分布较均匀(重要值在0.279—0.406), 如琉璃草(Cynoglossum furcatum)、原拉拉藤等。多年生杂类草在各种梯度干扰下占绝大优势, 如黄花鼠尾草、蟹甲草(Parasenecio forrestii)等, 且多年生杂类草在Ⅳ级干扰下重要值最高(0.879)。一年生禾草(如画眉草(Eragrostis pilosa)等)、多年生禾草(如小花剪股颖(Agrostis micrantha)等)均为适口性较强的植物, 重要值均随干扰强度减小而逐渐增大。多年生豆科植物的重要值处于波动中, 且在Ⅳ级干扰下达到最大, 在对照(CK)中为0。

|
| 图 2 不同强度干扰下草本层生活型功能群重要值 Fig. 2 The important value of plant community functional groups in herbaceous layer under different levels of disturbance |
研究区植物群落物种多样性随干扰梯度、植物群落退化程度的变化而改变(图 3)。在总体来看, 随干扰梯度增加, Shannon-Wiener指数H和物种丰富度指数D、Simpson指数H′和Pielou均匀度指数Jsw变化趋势一致。就乔木层而言, I级干扰下物种丰富度指数D最低, 显著低于Ⅳ级放牧干扰。除对照(CK), 轻度干扰Ⅳ的Simpson指数H′显著高于其他梯度。各梯度干扰下Pielou均匀度指数Jsw、Shannon-Wiener指数H均不存在显著差异。在Ⅰ—Ⅳ级干扰梯度中, Ⅳ级干扰梯度下的植被丰富度、均匀度、优势度均最高, I级干扰梯度下的物种丰富度虽最低, 但就其物种分布离散程度并非最低。就灌木层而言, I级干扰梯度下各多样性指数均最低, Ⅳ级干扰梯度下各多样性指数均显著最高。就草本层而言, Pielou均匀度指数在各梯度干扰下均无显著差异, 其余指数均在I级干扰梯度下最低, Ⅳ级干扰梯度下最高。由此可见放牧干扰对夹金山森林群落结构、物种多样性等影响较大。

|
| 图 3 不同强度放牧干扰下针阔混交林群落的物种多样性指数 Fig. 3 Species diversity of coniferous-broad-leaved mixed forest under different levels of disturbance D: 物种丰富度指数Richness index;H: Shannon-Wiener指数Shannon-Wiener index;H′: Simpson优势度指数Simpson dominance index;Jsw: Pielou均匀度指数Pielou eveness index; 不同小写字母表示不同生境间的多样性指数差异显著(P<0.05) |
由图 4可得, 除III级干扰群落外, 灌木地上和地下生物量均随着干扰程度的增加呈递减趋势, 且III级干扰梯度是各部分生物量剧烈变化的重要拐点。灌木层、草本层地上生物量均明显大于地下生物量。就灌木层而言, 除II级干扰外, 其余处理下灌木层地上、地下生物量均显著大于I级干扰。就草本层而言, 草本层地上生物量随干扰程度增加呈递减变化, 但地下生物量在III级干扰下又有所上升。相比于对照组, 林下枯落物量随干扰梯度增加分别降低了13.1%、32.04%、44.52%、85.52%, 即干扰越剧烈, 枯落物量越小。随干扰强度增大, 灌木层地下与地上生物量比值变化不明显(由0.375到0.371), 而草本层明显降低(由0.703到0.195), 即草本层植物群落增加了对地上生物量的投入。

|
| 图 4 不同强度放牧干扰下植物生物量分配情况 Fig. 4 Distribution of plant biomass under different grazing intensities GUB: 灌木层地上生物量Aboveground biomass of shrub layer;GDB: 灌木层地下生物量Underground biomass of shrub layer;LIB: 枯落物Litter;CUB: 草本层地上生物量Aboveground biomass of herb layer;CDB: 草本层地下生物量Underground biomass of herb layer; 不同小写字母表示不同生境间生物量的差异显著(P<0.05) |
由图 5可得, 夹金山针阔混交林内的乔木优势种幼苗、幼树的占比随干扰梯度增加呈明显减少趋势, IV级干扰梯度下的幼苗占比显著高于其他干扰梯度(51.0%), 群落更新能力最强。I级干扰梯度下的幼苗占比最低(1.21%), 且林内几乎未发现成活的乔木优势种幼树, 受放牧干扰最为严重。除I级干扰梯度, 其余干扰梯度下的幼树占比与对照(CK)均无显著差异。

|
| 图 5 不同强度放牧干扰下针阔混交林群落更新潜力 Fig. 5 Regeneration potential of coniferous-broad-leaved mixed forest under different levels of disturbance 不同小写字母表示不同生境间的更新能力差异显著(P<0.05) |
选用各层次生物量作响应变量, 物种多样性作为解释变量, 去除贡献小的部分变量, 对5个干扰等级的20个样地进行RDA分析排序(图 6)。结果显示, RDA1、2轴的特征值分别为0.609和0.153, 前两轴能够解释各层次生物量变化的76.2%。沿RDA 1轴从右至左, 依次出现Ⅱ、Ⅰ、Ⅳ、CK、Ⅲ级干扰群落, 乔灌草植被多样性提升, 各部分生物量增加。4个干扰等级群落在RDA排序图上的分布揭示了重要的环境梯度, 其分布格局吻合较好。灌木层地上地下生物量与枯落物量相关度大于草本层, 初步表明生境内枯落物量主要依赖于灌木植物补给。灌、草本层各物种多样指数(除Pielou均匀度指数Jsw外)与各层次生物量呈正相关。乔木层各多样性指数与灌木层地上地下生物量、枯落物量均呈正相关, 且相关度强于草本层。乔木层H、H、Jsw与草本层地上地下生物量呈负相关, 而D与其呈正相关。

|
| 图 6 植物群落物种多样性和生物量的RDA二维排序 Fig. 6 RDA two-dimensional ordering of species diversity and biomass of plant communities in Jiajin Mountain T-H: 乔木层Shannon-Wiener指数Shannon Wiener index of tree layer;T-H′: 乔木层Simpson指数Simpson index of tree layer;T-D: 乔木层物种丰富度指数Species richness index of tree layer;S-D: 灌木层物种丰富度指数Species richness index of shrub layer;H-H′: 草本层Simpson指数Simpson index of herbaceous layer;H-H: 草本层Shannon-Wiener指数Shannon Wiener index of herbaceous layer;H-D: 草本层物种丰富度指数Species richness index of herbaceous layer |
随着生境压力逐渐加大(由Ⅳ到Ⅰ), 适口性较强的优良草种(如禾本科(Poaceae)、豆科(Fabaceae)等)被采食率升高, 且一年生草本重要值逐渐增大, 多年生草本重要值逐渐降低。这说明一年生草本对环境改变的响应更加灵敏, 更新、适应能力更强;可食性较高的禾草类草本被二代野猪优先采食, 导致适口性较强的禾草重要值、盖度及优势度降低;可食性较低的杂类草本比例增加, 表现为“功能性组分冗余”[7]。虽然研究区内的二代野猪对牧草的选择性采食导致群落内各功能群的再生能力、补偿性生长能力不同, 但禾草和杂类草功能群之间存在补偿效应, 使得群落相对稳定[7, 27]。群落中的优势种的替代和变化, 是群落对干扰强度响应的主要特征, 直接决定着群落的发生与发展的方向[8, 28]。本实验乔木层川滇长尾槭、灌木层蔷薇科、草本层原拉拉藤等均在干扰梯度增加的过程中逐渐或始终占据优势地位, 为抗干扰能力强的物种。乔灌草三层均在Ⅳ级干扰下Shannon-Wiener指数H、物种丰富度指数D、Simpson指数H′、Pielou均匀度指数Jsw最高, 即植被丰富度、均匀度、优势度均最高, 支持中度干扰假说[29]。这是因为野猪的轻度翻拱和采食行为能刺激植被快速生长, 以补偿优良草种的损失, 但当盖度达到一定水平时, 这种功能补偿往往产生部分植物的生长冗余[7]。
随着放牧压力的增加, 物种的表型性状会通过地上、地下不同器官的资源配置模式的变化而发生变化, 最终向种群适应度提高的方向发展[16]。随干扰强度增大, 草本层反倒增加对地上部分的投入, 而灌木层地上地下生物量比值几乎未受影响, 这不符合功能平衡假说[30], (1)因为研究区二代野猪活动范围大, 对土地的严重踩踏翻拱及对植物的频繁采食, 严重扰乱了植物地下与地上光合产物分配比值;(2)因为二代野猪的采食促进的对象仅为杂类草本地上部分;(3)因为重度干扰区灌木层蔷薇科植被呈低矮倒伏或植物体小型化, 几乎未受任何影响。在近似土壤—大气系统的碳循环, 枯落物分解速率和植物向土壤输入养分速率是支配整个夹金山系统碳循环最关键的变量[31]。此研究区干扰越剧烈, 枯落物量越小, 是因为重度干扰等级生境的本底植被条件较差, 植物向土壤输入养分速率降低;也可能是因为野猪不断地取食与践踏使得地表土壤温度迅速提升[32], 又或是放牧减少了植物群落盖度从而可能增加了光降解[3], 加速了表层土壤有机质的分解。
乔、灌、草三层物种多样性与灌、草层生物量均呈正相关, 即物种多样性的适度提高会对生物量的增加有促进作用[33], 这说明放猪干扰造成植被破坏会间接影响到植被生物量高低。乔木优势种幼苗、幼树的占比随干扰梯度增加呈明显减少趋势, 重度干扰后生境内幼苗占比最低甚至几乎无成活的乔木优势种幼树, 可见受放猪干扰最为严重, 这涉及到未来夹金山群落更新能力的提升, 亟待提出可行的解决方案。
5 结论因为可达性不同, 二代野猪对四川省夹金山植物群落的干扰程度随距养猪场的远近而不同。乔灌草三层均在Ⅳ级干扰下物种多样性水平最高, I级干扰下物种数最贫乏。乔灌草三层均在Ⅳ级度干扰下植被丰富度、均匀度、优势度均最高。I级干扰梯度下林内几乎未发现成活的乔木优势种幼树, 受放牧干扰最为严重。因此, 建议将Ⅰ、Ⅱ级重度干扰样地(即距养猪场2000 m范围内)的管理措施设置为禁牧或轮牧, 并且补植山梅花(Philadelphus incanus)、红桦(Betula albosinensis)等乔木树种以维持物种多样性水平, 重点维护川滇长尾槭、糙皮桦幼苗幼树的生长以促进更新。
| [1] |
靳茗茗, 徐增让, 成升魁. 藏北高寒草地植被和土壤对不同放牧强度的响应. 生态学报, 2020, 40(23): 8753-8762. |
| [2] |
薛睿, 郑淑霞, 白永飞. 不同利用方式和载畜率对内蒙古典型草原群落初级生产力和植物补偿性生长的影响. 生物多样性, 2010, 18(3): 300-311. |
| [3] |
Song X X, Wang L, Zhao X, Liu C, Chang Q, Wang Y, Xu T T, Wang D L. Sheep grazing and local community diversity interact to control litter decomposition of dominant species in grassland ecosystem. Soil Biology and Biochemistry, 2017, 115: 364-370. DOI:10.1016/j.soilbio.2017.09.003 |
| [4] |
温璐, 董世魁, 朱磊, 施建军, 刘德梅, 王彦龙, 马玉寿. 环境因子和干扰强度对高寒草甸植物多样性空间分异的影响. 生态学报, 2011, 31(7): 1844-1854. |
| [5] |
戴黎聪, 柯浔, 曹莹芳, 张法伟, 杜岩功, 李以康, 郭小伟, 李茜, 林丽, 马建军, 曹广民. 青藏高原矮嵩草草甸地下和地上生物量分配格局及其与气象因子的关系. 生态学报, 2019, 39(2): 486-493. |
| [6] |
吴红宝, 高清竹, 干珠扎布, 李钰, 闫玉龙, 胡国铮, 王学霞, 严俊, 何世丞. 放牧和模拟增温对藏北高寒草地植物群落特征及生产力的影响. 植物生态学报, 2019, 43(10): 853-862. |
| [7] |
董全民, 马玉寿, 李青云, 赵新全, 王启基, 施建军. 牦牛放牧率对小嵩草高寒草甸暖季草场植物群落组成和植物多样性的影响. 西北植物学报, 2005, 25(1): 94-102. |
| [8] |
王向涛, 张世虎, 陈懂懂, 谈嫣蓉, 孙大帅, 杜国祯. 不同放牧强度下高寒草甸植被特征和土壤养分变化研究. 草地学报, 2010, 18(4): 510-516. |
| [9] |
牛钰杰, 杨思维, 王贵珍, 刘丽, 杜国祯, 花立民. 放牧干扰下高寒草甸物种、生活型和功能群多样性与生物量的关系. 生态学报, 2018, 38(13): 4733-4743. |
| [10] |
Wirthner S, Frey B, Busse M D, Schütz M, Risch A C. Effects of wild boar (Sus scrofa L.) rooting on the bacterial community structure in mixed-hardwood forest soils in Switzerland. European Journal of Soil Biology, 2011, 47(5): 296-302. DOI:10.1016/j.ejsobi.2011.07.003 |
| [11] |
安钰, 安慧. 宁夏荒漠草原优势植物生长及生物量分配对放牧干扰的响应. 西北植物学报, 2015, 35(2): 373-378. |
| [12] |
Fenetahun Y, Xu X W, You Y, Wang Y D. Effects of vegetation cover, grazing and season on herbage species composition and biomass: a case study of Yabello rangeland, Southern Ethiopia. Journal of Resources and Ecology, 2020, 11(2): 159-170. DOI:10.5814/j.issn.1674-764x.2020.02.004 |
| [13] |
Zuo X A, Zhang J, Lv P, Wang S K, Yang Y, Yue X Y, Zhou X, Li Y L, Chen M, Lian J, Qu H, Liu L X, Ma X J. Effects of plant functional diversity induced by grazing and soil properties on above-and belowground biomass in a semiarid grassland. Ecological Indicators, 2018, 93: 555-561. |
| [14] |
Du Y G, Ke X, Guo X W, Cao G M, Zhou H K. Soil and plant community characteristics under long-term continuous grazing of different intensities in an alpine meadow on the Tibetan plateau. Biochemical Systematics and Ecology, 2019, 85: 72-75. |
| [15] |
Dai L C, Fu R Y, Guo X W, Du Y G, Lin L, Zhang F W, Li Y K, Cao G M. Long-term grazing exclusion greatly improve carbon and nitrogen store in an alpine meadow on the northern Qinghai-Tibet Plateau. CATENA, 2021, 197: 104955. |
| [16] |
Ye R H, Liu G F, Chang H, Shan Y M, Mu L, Wen C, Te R, Wu N T, Shi L, Liu Y H, Wang H M, Yun X J, Liu G X, Li F. Response of plant traits of Stipa breviflora to grazing intensity and fluctuation in annual precipitation in a desert steppe, northern China. Global Ecology and Conservation, 2020, 24: e01237. |
| [17] |
Dai L C, Guo X W, Du Y G, Ke X, Cao Y F, Li Y K, Cao G M, Zhang F W. Thirteen-year variation in biomass allocation under climate change in an alpine Kobresia meadow, northern Qinghai-Tibetan Plateau. Grass and Forage Science, 2019, 74(3): 476-485. |
| [18] |
苗百岭, 梁存柱, 史亚博, 梁茂伟, 刘钟龄. 降水变化对内蒙古典型草原地上生物量的影响. 植物生态学报, 2019, 43(7): 557-565. |
| [19] |
张荣, 董洪君, 周润惠, 余飞燕, 王敏, 陈聪琳, 喻静, 郝建锋. 四川夹金山灌丛群落根系特征对土壤抗冲性的影响. 生态学杂志, 2020, 39(11): 3558-3566. |
| [20] |
陈富斌, 赵永涛, 兰立波. 论夹金山脉大熊猫栖息地的世界自然遗产价值. 山地学报, 2002, 20(6): 687-694. |
| [21] |
刘金鑫. 放牧干扰对山地森林/干旱河谷交错带植物群落特征的影响[D]. 雅安: 四川农业大学, 2012.
|
| [22] |
王晓. 浑善达克沙地榆树疏林生态系统组成、空间格局分布及其对放牧干扰的响应[D]. 北京: 北京林业大学, 2016.
|
| [23] |
余飞燕, 王坤悦, 叶鑫, 董洪君, 黄凯, 罗志力, 郝建锋. 金马河温江段河岸带不同生境草本群落物种多样性和生物量变化研究. 草地学报, 2020, 28(3): 793-800. |
| [24] |
杨昆, 管东生. 森林林下植被生物量收获的样方选择和模型. 生态学报, 2007, 27(2): 705-714. |
| [25] |
方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. |
| [26] |
郭其强, 张文辉, 曹旭平. 基于模糊综合评判的森林群落稳定性评价体系模型构建——以黄龙山主要森林群落为例. 林业科学, 2009, 45(10): 19-24. |
| [27] |
高润梅, 石晓东, 郭跃东. 山西文峪河上游河岸林群落稳定性评价. 植物生态学报, 2012, 36(6): 491-503. |
| [28] |
王明明, 刘新平, 何玉惠, 张铜会, 魏静, 车力木格, 孙姗姗. 科尔沁沙地封育恢复过程中植物群落特征变化及影响因素. 植物生态学报, 2019, 43(8): 672-684. |
| [29] |
张鲜花, 朱进忠, 吴咏梅. 天山北坡羊茅草原群落结构与数量特征的动态变化. 草地学报, 2012, 20(5): 819-824. |
| [30] |
Grime J P. Vegetation classification by reference to strategies. Nature, 1974, 250(5461): 26-31. |
| [31] |
王敏, 苏永中, 杨荣, 杨晓. 黑河中游荒漠草地地上和地下生物量的分配格局. 植物生态学报, 2013, 37(3): 209-219. |
| [32] |
展鹏飞, 闫鹏飞, 刘振亚, 王行, 张贇, 张勇, 崔海军, 肖德荣. 藏香猪放牧对滇西北高原湿地土壤CO2通量的影响. 生态学报, 2019, 39(9): 3309-3321. |
| [33] |
韩泽民, 李源, 王熊, 菅永峰, 周靖靖, 佃袁勇, 黄光体. 不同演替程度下马尾松人工林生物多样性对生物量的影响. 浙江农林大学学报, 2021, 38(2): 246-252. |