 2021, Vol. 41
2021, Vol. 41文章信息
- 晏妮, 张腾, 熊康宁, 陈倩, 郭丁力, 刘之威
- YAN Ni, ZHANG Teng, XIONG Kangning, CHEN Qian, GUO Dingli, LIU Zhiwei
- 世界自然遗产地陆生藻类物种多样性比较研究
- Terrestrial algae diversity of the two world natural heritage sites in China
- 生态学报. 2021, 41(24): 9593-9603
- Acta Ecologica Sinica. 2021, 41(24): 9593-9603
- http://dx.doi.org/10.5846/stxb202103080628
-
文章历史
- 收稿日期: 2021-03-08
- 修订日期: 2021-08-17





2. 贵州师范大学生命科学学院, 贵阳 550001;
3. 中山大学生命科学学院, 广州 510275;
4. 华南理工大学环境与能源学院, 广州 510641
2. School of Life Sciences, Guizhou Normal University, Guiyang 550001, China;
3. School of Life Sciences, Sun Yat-sen University, Guangzhou 510275, China;
4. College of Environment and Energy, South China University of Technology, Guangzhou 510641, China
藻类是地球早期生命形式, 并且由藻类植物经水生向陆生发展产生了早期陆地植物[1], 藻类不仅是重建早期陆生植物的系统演化、记录地球地质历史变迁的重要研究对象及关键证据, 亦是地球上最重要的初级生产者, 对水生及陆生生态系统的稳定和更新都有着极其重要的作用。Knoll和Barghoorn基于现代蓝藻的降解作用成功地解释了前寒武纪蓝藻化石的多样性, 通过对时间上相距约10—12亿年之间的化石藻Palaeopleurocapsa wopfnerii与现代蓝藻Pleurocapsa的比较揭示了科水平上的亲缘关系, 并指出很可能是同属[2]。茅绍智等对早中新世的沟鞭藻植物地理作了初步研究, 指出沟鞭藻的分布主要受控于纬度(温度)和环境, 并且推演揭示中新世的气候、植被、海陆分布格局等[3]。烟晓玲通过对太行山中段古元古代东冶组藻礁的研究, 推测在古元古代华北地台已基本成型[4]。王教元等基于硅藻功能群重建200年海西海生态环境演变历史[5]。吴亚生等对湖南慈利晚二叠世藻类化石中藻类群落演替的研究, 揭示了当地的历史水位的变化[6]。穆西南通过对贵州西部晚二叠世地层丰富的钙藻研究推演贵州当时所处的低纬度热带近岸浅海环境[7]。许多钙藻化石残体与现存相关藻类实体常一致而采用现代钙藻的分类原则命名及推断亲缘关系[8]。利用现代藻类细胞外部形态、分裂方式、遗传信息等去分析已经获得的古生物化石标本、推演古地理、古气候等将今论古的方式, 已成为了解生命起源、生物进化、地球演化史的重要手段之一。
施秉云台山白云岩喀斯特、赤水丹霞两世界自然遗产地分别位于贵州东南部与北部。施秉云台山白云岩喀斯特是古生代寒武纪中寒武世白云岩经溶蚀及相应构造作用形成的除石灰岩外的另一种喀斯特地貌, 代表了典型的白云岩喀斯特青年期地貌演化的特征[9]。赤水丹霞代表着丹霞地貌青年期[10], 是中新生代陆相碎屑岩经流水侵蚀、重力崩塌和风化剥落等多种地质作用的产物[11]。两世界自然遗产地经历了漫长的地球历史演变, 具有不可再生性, 满足世界自然遗产的第Ⅶ、Ⅷ两条标准, 反映了白云岩喀斯特地貌、红层地貌不同地球演化主要阶段, 具有各自独特的生态系统和复杂生态过程。目前对于两世界遗产地的研究主要包括以下几个平行并重的方向:美学价值深度挖掘、地貌特征与发育演化、遗产地的监测保护与管理、生态系统结构与功能、生物多样性与分布格局[9, 12], 但基于生物地理学思想, 融合地学的宏进化思维与生物微进化观点, 将生物多样性、生物生态过程与地质地貌协同演化相结合的研究仍较欠缺, 现有相关报道也多局限于对两世界自然遗产地生物分布的环境因素分析[13], 高等植物区系特征探讨[14]等。与高等植物相比, 低等的藻类具有进化时间长、类群分歧时期较早及进化方向多元化特点。从单细胞到不定形群体再到定形群体, 从无分化丝状结构到逐步分化的异丝体、膜状体等系统与个体演化过程中, 包含了从原核光合生物、真核生物、多细胞真核生物三个生物进化史中最为关键的阶段。因此, 在化石记录缺乏的现实下, 基于现存藻类开展其系统演化研究, 对还原早期生命演化历史、揭示蕴藏在藻类进化史中的地质变迁信息都具有重要作用。本研究以两个不同地质地貌的世界自然遗产地现存的陆生藻类的物种多样性、种组特征等内容为切入点, 从地质年代、地层岩性等方面讨论了陆生藻类与地质地貌的协同演化关系, 为未来围绕着陆生藻类的形态学及繁殖生物学对其适应不同的陆生环境条件深入开展适应性研究, 基于微进化角度, 将“陆生藻类分子钟”研究与已有化石证据结合, 重演两世界自然遗产地的地球演变史及生态过程提供了可行的研究对象及本底数据。
1 材料与方法 1.1 研究区概况施秉云台山世界自然遗产地位于贵州省东南部黔东南州施秉县北部(108°01′—108°10′E, 27°13′—27°04′N), 由云台山、外营台、轿顶山及大田垴等群峰组成, 地势西北高, 东南低, 海拔486—1869m, 属亚热带季风湿润气候区, 水热条件优越, 年均温16℃, 年均降水1220mm, 白云岩分布广泛, 以寒武纪纯白云岩风化而来的薄层石灰土为主。区域内植物物种丰富, 种子植物140科1087种[14], 蕨类植物25科127种, 苔藓植物50科286种[15]。森林植被主要类型为常绿阔叶林[16]。赤水丹霞世界自然遗产地位于贵州省西北部的赤水市(105°42′—106°10′E, 28°18′—28°44′N), 包括西区和东区两个核心片区, 覆盖1个国家级自然保护区、2个国家森林公园和1个国家级风景名胜区, 海拔240—1730m, 区域内沉积形成有紫红色泥岩、粉砂质岩及砂岩等, 代表性土壤为紫色土, 属于亚热带季风湿润气候, 年均降水1292mm, 气候适宜, 年均温18℃。据不完全统计, 区域内种子植物171科1715种, 蕨类植物41科249种[17], 苔藓植物52科318种[18]。植被组成类型为常绿阔叶林、针阔混交林以及竹林, 同时是现今亚洲最大的桫椤分布区。
1.2 样品采集与鉴定施秉云台山世界自然遗产地的采样区布设于云台山(108°01′36″—108°10′52″E, 27°13′52″—27°04′51″N), 地球同纬度上最古老、发育最完好的白云岩喀斯特所在地。从云台关、沿樱桃湾、排云关等设置16个采样位点。赤水丹霞采样区设于该世界自然遗产地西区的十丈洞(105°43′48″—105°45′51″E, 28°15′23″—28°22′47″N), 包含蝴蝶桥、孔明泉、大瀑布等10个采样位点和东区的桫椤自然保护区(E105°57′54″—106°07′07″, N28°20′19″—28°28′40″)两岔河、大水沟、甘沟等6个采样位点, 两地红层地貌发育且保存了完好的亚热带森林生态环境, 是赤水丹霞地貌的典范(采样位点见表 1)。本研究在上述代表性区域内开展点线面结合样品广泛普采, 针对不同基质(石壁、树干、土表等)表面划取不同形态、不同颜色藻类群落, 置于盛有甘油-甲醛-水溶液(1 ∶ 1 ∶ 8)固定剂的标本瓶中, 并详细记录包括标本编号、采集日期、生态条件、群落形态特征、颜色和采集人等信息。每个基质采样后用70%酒精擦拭采样工具以防交叉污染。从2014年至今2020年, 共收集标本547号, 其中施秉云台山275号, 赤水丹霞272号, 标本保存于贵州师范大学生命科学学院藻类研究室。通过OLYMPAS-CX31三目光学显微镜进行物种分类鉴定、优势种记录及显微拍照。
| 云台山位点 Sample sites Shibing Yuntai Mountain (SYM) |
经纬度 Latitude and Longitude |
海拔/m Elevation |
赤水丹霞位点 Sample sites of Chishui Danxia(CD) |
经纬度 Latitude and Longitude |
海拔/m Elevation |
|
| 云台关 | 108°06′58″, 27°06′47″ | 834 | 空谷佳人 | 105°44′34″, 28°17′16″ | 617 | |
| 樱桃湾 | 108°06′30″, 27°06′29″ | 843 | 奇兵古道 | 105°44′23″, 28°22′12″ | 613 | |
| 排云关 | 108°06′15″, 27°06′32″ | 891 | 美人梳瀑布 | 105°44′16″, 28°22′08″ | 653 | |
| 野牛洞 | 108°06′12″, 27°06′32″ | 856 | 红雨栈道 | 105°44′31″, 28°21′43″ | 704 | |
| 盘藤道 | 108°06′12″, 27°06′33″ | 874 | 通幽门 | 105°44′02″, 28°21′46″ | 654 | |
| 印斗阁 | 108°06′24″, 27°06′37″ | 902 | 金蟾石 | 105°45′36″, 28°15′39″ | 628 | |
| 坪山垴 | 108°06′25″, 27°06′36″ | 910 | 蝴蝶桥 | 105°44′13″, 28°21′32″ | 667 | |
| 头道灵关 | 108°06′28″, 27°07′06″ | 735 | 孔明泉 | 105°44′20″, 28°21′39″ | 708 | |
| 二道灵关 | 108°06′26″, 27°07′12″ | 727 | 谢安亭 | 105°44′16″, 28°21′35″ | 658 | |
| 滴水观音 | 108°06′22″, 27°07′14″ | 693 | 大瀑布 | 105°44′31″, 28°21′32″ | 740 | |
| 黄州河边 | 108°07′14″, 27°06′33″ | 630 | 两岔河 | 106°00′38″, 28°25′18″ | 711 | |
| 周公殿 | 108°06′20″, 27°07′17″ | 720 | 大水沟 | 106°01′13″, 28°25′52″ | 826 | |
| 会仙桥 | 108°06′26″, 27°07′20″ | 798 | 甘沟 | 106°01′26″, 28°25′26″ | 708 | |
| 徐公殿 | 108°06′27″, 27°07′29″ | 850 | 钟山 | 106°00′07″, 28°25′55″ | 728 | |
| 滴水岩 | 108°06′24″, 27°07′30″ | 882 | 硝岩坡 | 106°01′08″, 28°25′44″ | 452 | |
| 香上 | 108°06′21″, 27°07′26″ | 787 | 狮子岩 | 106°00′38″, 28°25′18″ | 529 |
科多样性指数DF:
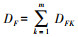
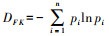
属多样性指数DG:
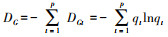
标准化指数DG-F: DG-F=1-DG/DF, 规定:记录中若所有的科都是单种科, 即DF=0时, 则该地区的G-F指数为零, 非单种科越多, G-F指数越高[19]。
1.3.2 Jaccard相似性系数SJ(%)=a×100%/(a+b+c), 其中, SJ为两比较区对应分类单元相似性系数, a为两共有的分类单元数, b、c分别为仅出现于一地的分类单元数[20]。
1.3.3 地理分布区类型及Bray-Curtis聚类参照吴征镒等《中国自然地理-植物地理》[21], 对两世界遗产地的陆生藻类地理分布区类型进行划分, 并对种类的存在与否进行赋值(存在赋值1, 不存在赋值0), 采用组间平均聚类法进行分层聚类分析, 构建Bray-Curtis相似矩阵。
2 结果与分析 2.1 两世界自然遗产地陆生藻类的种类组成经显微观察鉴定统计(表 2), 施秉云台山共发现陆生藻类3门6纲18目26科67属238种, 其中蓝藻种数为177种, 占总种数的74.37%, 绿藻和硅藻分别为39种、22种, 依次占总种数的16.39%和9.24%;优势藻科为色球藻科Chroococcaceae、颤藻科Oscillatoriaceae、伪枝藻科Scytonemataceae、念珠藻科Nostocaceae, 分别包含13属、4属、5属、7属, 共计29属, 占总属数的43.28%, 物种总数为162种, 占总种数的68.07%, 优势属集中在色球藻科的粘球藻属Gloeocapsa、色球藻属Chroococcus, 种数分别为16种、17种, 在总种数中的占比为13.87%。
| 分类数 Classification |
蓝藻Cyanophyta | 绿藻Chlorophyta | 硅藻Bacillariophyta | |||||
| 施秉云台山 (SYM) |
赤水 (CD) |
施秉云台山 (SYM) |
赤水 (CD) |
施秉云台山 (SYM) |
赤水 (CD) |
|||
| 科数Family | 9 | 7 | 12 | 10 | 5 | 7 | ||
| 占总科数比例% of the total | 34.62% | 26.92% | 46.15% | 38.46% | 19.23% | 26.92% | ||
| 属数Genus | 40 | 28 | 18 | 17 | 9 | 9 | ||
| 占总属数比例% of the total | 59.70% | 52.83% | 26.87% | 32.08% | 13.43% | 16.98% | ||
| 种数Species | 177 | 146 | 39 | 31 | 22 | 24 | ||
| 占总种数比例% of the total | 74.37% | 72.64% | 16.39% | 15.42% | 9.24% | 11.94% | ||
赤水丹霞共记录陆生藻类3门6纲19目24科54属201种, 其中蓝藻种数为146, 占总种数的72.64%, 余下为绿藻门31种、硅藻门24种, 分别占15.42%, 11.94%;赤水丹霞优势科为色球藻科Chroococcaceae、颤藻科Oscillatoriaceae, 属数分别为10属、5属, 共计15属, 占总属数的27.78%, 物种总数为116种, 占总种数的57.71%, 优势属表现为粘球藻属Gloeocapsa, 包含25种, 在总种数的占比为12.44%。
2.2 两世界自然遗产地陆生藻类多样性施秉云台山与赤水丹霞两地陆生藻类G-F多样性指数见表 3, 两地的G-F多样性标准化指数值均大于0.7, 处于较高水平, 反映两地均具有丰富的陆生藻类物种多样性。从两地各自三门藻类纵向比较上, 科属多样性指数都表现出相同的规律:蓝藻>绿藻>硅藻, 蓝藻的DG、DF值高于绿、硅藻的值, 两研究区存在明显的蓝藻优势科属现象, 绿硅藻科属多样性指数相差不大;从两地三门藻类横向比较上, 施秉云台山蓝藻科的多样性指数DF值为9.37, 明显高于赤水丹霞的蓝藻科多样性指数6.71, 施秉云台山蓝藻更为丰富, 除两地共有优势科色球藻科Chroococcaceae和颤藻科Oscillatoriaceae外, 施秉云台山白云岩喀斯特还有伪枝藻科Scytonemataceae、念珠藻科Nostocaceae丝状固氮蓝藻优势。施秉云台山绿藻物种数量虽高于赤水丹霞, 但DG-F值却小于赤水丹霞, 这与调查单种科较多结果一致, 非单种科越多, G-F指数越高。同样硅藻也是赤水丹霞的单种科高于施秉云台山, 因此G-F指数较低。
| 分类Classification | DG | DF | DG-F | |||||
| SYM(DGY) | CD (DGC) | SYM (DFY) | CD (DFC) | SYM (DG-F(Y)) | CD (DG-F(C)) | |||
| 蓝藻Cyanophyta | 2.65 | 2.32 | 9.37 | 6.71 | 0.72 | 0.66 | ||
| 绿藻Chlorophyta | 0.75 | 0.70 | 3.25 | 3.99 | 0.77 | 0.83 | ||
| 硅藻Bacillariophyta | 0.41 | 0.49 | 2.10 | 1.85 | 0.80 | 0.73 | ||
| 全部藻类All the algae | 3.81 | 3.51 | 14.72 | 12.55 | 0.74 | 0.72 | ||
| DG: 属的多样性指数Diversity index of genus; DF: 科的多样性指数Diversity index of family; DG-F: 标准化多样性指数Standardized diversity index; | ||||||||
两世界自然遗产地总的科、属、种的相似性系数分别为:56.25%、44.05%、37.62%。科水平上, 相似性系值高于50%, 达到中度相似水平, 两地共分布18个相同科, 蓝藻、硅藻、绿藻三门相同科数分别为6、4、8(表 4)。
| 分类Phylum | 共有科Shared families | 非共有科Families appear at different sites | |
| 施秉云台山(SYM) | 赤水(CD) | ||
| 蓝藻门Cyanophyta | 色球藻科Chroococcaceae | 蓝柄藻科Cyanostylonacaea | 真枝藻科Stigonemataceae |
| 石囊藻科Entophysalidaceae | 皮果藻科Dermocarpaceae | ||
| 颤藻科Oscillatoriaceae | 微毛藻科Microchaetaceae | ||
| 伪枝藻科Scytonemataceae | |||
| 胶须藻科Rivulariaceae | |||
| 念珠藻科Nostocaceae | |||
| 硅藻门Bacillariophyta | 圆筛藻科coscinodiscaceae | 菱形藻科NNitzschiaceae | 异极藻科Gomphonemaceae |
| 脆杆藻科Fragilariaceae | 曲壳藻科Achnanthaceae | ||
| 舟形藻科Naviculaceae | 短缝藻科Eunotiaceae | ||
| 桥弯藻科Cymbellaceae | |||
| 绿藻门Chlorophyta | 绿球藻科Chlorococcaceae | 卵囊藻科Oocystaceae | 空星藻科Coelastruaceae |
| 小球藻科Chlorellaceae | 盘星藻科Pediastraceae | 溪菜科Prasiolaceae | |
| 丝藻科Ulotrichaceae | 栅藻科Scenedesmaceae | ||
| 胶毛藻科Chaetophoraceae | 鞘藻科Oedogoniaceae | ||
| 橘色藻科Trentepohliaceae | |||
| 刚毛藻科Cladophoraceae | |||
| 双星藻科Zygnemataceae | |||
| 鼓藻科Desmidiaceae | |||
属水平上, 两地仅为一般相似, 其差异性主要表现在施秉云台山的球状和丝状蓝藻属均较赤水丹霞丰富, 其多有的属为具明显胶鞘的单细胞或单细胞群体类, 包括聚球藻属Synechococcus、拟指球藻属Dactylococcopsis、平裂藻属Merismopedia、蓝柄藻属Cyanostylon、绿胶藻属Chlorogloea、皮果藻属Dermocarpa、裂胞藻属Clastidium(图 1)。两地硅藻均包含9属, 共有4属为脆杆藻属Fragilaria、舟形藻属Navicula、双眉藻属Amphora、桥弯藻属Cymbella, 非共有各5属(图 2)。绿藻中, 施秉云台山具有的如绿节藻属Chlorotylum、费氏藻属Fritschiella、刚毛藻属Cladophora暂未在赤水丹霞采集样品中检出。从三门藻类种的水平来看, 蓝藻种的相似性系数为43.56%, 绿硅藻虽科相似性较高, 但种间相似性却较低, 分别22.81%、24.32%, 也反映了更为进化的真核藻类较原核蓝藻生态适应性弱, 不同藻类对环境要求不同。

|
| 图 1 施秉云台山与赤水丹霞两世界自然遗产地陆生蓝藻属及物种数比较 Fig. 1 Differences at genus and species levels of the Cyanophyta between Shibing Yuntai Mountain (SYM) and Chishui Danxia (CD) |

|
| 图 2 施秉云台山与赤水丹霞两世界自然遗产地硅藻、绿藻属及物种数比较 Fig. 2 Differences at genus and species levels of the Chlorophyta and Bacillariophyta between Shibing Yuntai Mountain and Chishui Danxia |
基于维管植物区系类型与藻类文献记录, 将施秉云台山和赤水丹霞两世界自然遗产地陆生藻类地理分布类型划分为:世界分布、泛热带(热带亚热带)分布、温带分布、青藏高原分布、中国特有成份五个类型, 其中两地均呈现明显泛热带(热带亚热带)性质, 该分布类型占比分别为66.09%、59.21%(表 5)。在施秉云台山泛热带成分中, 蓝藻有107种, 占该地总泛热带分布陆生藻类物种数的93.04%, 对应赤水丹霞该数据分别为81种、90.00%。从藻类组成上, 球状细胞蓝藻中的色球藻属Chroococcus、粘球藻属Gloeocapsa、隐球藻属Aphanocapsa, 丝状蓝藻中的颤藻属Oscillatoria、伪枝藻属Scytonema、鞘丝藻属Lyngbya中绝大多数种类是热带与亚热带地区的常见种。在气候干旱的陆生环境中, 硅藻种类和数量通常较少, 两地调查的共有及非共有硅藻几乎都是世界分布, 仅少数如羽纹藻属Pinnularia北方羽纹藻P. borealis, 短缝藻属Eunotia柔弱短缝藻E. tenella、弧形短缝藻E. arcus等具温带性质;两地绿藻中种类相对较多的属, 如盘星藻属Pediastrum、栅藻属Scenedesmus、转板藻属Mougeotia、水绵属Spirogyra中种类也为世界普生性种类。中国特有成份均属蓝藻门的星球藻属Asterocapsa和肾胞藻属Nephrococcus, 施秉云台山的星球属物种较赤水丹霞丰富。总体上, 两地陆生绿硅藻物种数量少且普生种类居多, 从绿硅藻组成上无法反映两地地理分布类型的特点, 但陆生蓝藻组成较好地表征了两地的地理位置所处纬度范围。
| 分布区类型Areal types | 施秉云台山(SYM) | 赤水丹霞(CD) | |||
| S/S1 | P%/P1% | S/S1 | P%/P1% | ||
| 世界分布Cosmopolitan | 52/27 | - | 44/21 | - | |
| 泛热带分布Pantropic | 115/107 | 66.09%/93.04% | 90/81 | 59.21%/90.00% | |
| 温带分布Temperate | 45/34 | 25.86%/75.56% | 49/31 | 32.24%/63.26% | |
| 青藏高原分布Tibetan plateau | 1/1 | 0.57%/100% | 8/8 | 3.29%/100% | |
| 中国特有成份Endemic to China | 13/13 | 7.47%/100% | 5/5 | 5.26%/100% | |
| 分布不祥Unknown distribution | 12/6 | - | 5/5 | - | |
| S: 总物种数Total species number; S1: 蓝藻物种数Species number of the Cyanophyta; P%: 某分布区类型占总分布区类型百分比Percentage of the total types given a specific area; P1%: 某分布区类型蓝藻物种数占该分布区总物种数百分比Percentage of the whole Cyanophyta species of the areas analyzed; 世界分布种类由于遍布世界各大洲无明显分布中心而无法反映两地地理分布特点, 在进行地理分布区统计时扣除不计 | |||||
为进一步探讨处于贵州同一纬度范围的不同地理位置、不同岩性背景下的陆生藻类的相似性, 选择位于贵州东南部的板岩为主的雷公山、贵州西南部的白云岩喀斯特马岭河峡谷及贵州中部贵阳的石灰岩喀斯特的陆生藻类, 与施秉云台山、赤水丹霞两地的陆生藻类物种进行聚类分析(图 3), 结果显示:第一类为贵州西南的马岭河峡谷与贵州东南的雷公山自然保护区;第二类为贵州东南的施秉云台山和贵州北的赤水丹霞;第三类为贵州中部的贵阳。从地理位置上, 施秉云台山并未首先与同处于贵州东南部, 纬度位置更为接近的雷公山聚为一类;从地貌岩性上, 也未与更为相近的马岭河峡谷聚为一类, 而与位置距离较远, 岩性截然不同的赤水丹霞关系更近。

|
| 图 3 五个地区藻类种间相似性聚类分析 Fig. 3 Dendrogram among the algae species from five different areas analyzed |
生命起源和早期进化发生于海洋, 最早生命证据来自于距今37亿年的海洋沉积物[22]。寒武纪时期, 海洋浮游藻类作为海洋食物链中的基本组成部分, 其丰度和分异度的升高被认为“寒武纪生命大爆发”的触发机制, 并且对后生生物的分异产生了重要影响[23]。郑书粲对寒武纪微体生物群落研究发现主要以单球藻和多球藻为主, 包括如光面球藻Leiosphaeridia sp.、色球藻Chrococcaceae sp.、粘球藻Gloeocapsa sp.等, 以及发现了可以比拟现代颤藻的长线体蓝藻类Oscillatoriopsis sp., 具异型胞的念珠藻Nostoc sp.等[24]。谯文浪等的研究揭示施秉白云岩的沉积环境为局限碳酸盐台地, 海水不流畅, 水体较浅, 温度较高, 适合于个体微小的浮游藻类的生长与繁衍[25]。本研究施秉云台山现代陆生藻类组成及优势属特征与报道的古地理时期组成基本一致。杨瑞东等对贵州黔东南的寒武纪宏观藻群研究中发现了包含绿藻、红藻、褐藻以及分类地位不明的宏观藻类化石共20属, 出现了分异度高的绿藻、褐藻、红藻等分枝形态的藻类, 并且肯定钙质藻类出现[26]。寒武纪早期, 藻类生物群落已经相当发达, 这也就解释了施秉云台山具有丰富藻类的重要原因。但随着贵州由东向北海浸退却, 黔东南地区最先出露转变为陆地环境, 一些分异度高的丝状海洋褐藻、红藻相较于原始的单细胞蓝藻类难于适应陆地生活而消失。
丹霞地貌为一种红层地貌, 与施秉云台山的地质演化历史相比, 赤水丹霞地貌发育较晚, 是中生代至新生代的产物, 又因陆相沉积而相异于施秉云台山的海相沉积[27]。研究表明, 中国的红层形成于三叠纪至第三纪的漫长地质历史时期[28], 但有报道指出赤水丹霞初始发育于第四纪[29]。王振等又通过轮藻化石揭示贵州中部两个盆地的红层沉积时代全部归于上白垩纪[30]。目前丹霞地貌岩石基础的时代和岩相、丹霞地貌发育控制因素和演化模式等尚存争议[11]。前人依据藻体简单到复杂及化石出现的年代, 将现存的蓝藻分为了三个纲, 即:色球藻纲、藻殖段纲、真枝藻纲。前两纲化石藻均来源于寒武纪时期, 真枝藻纲的化石是距今4亿年左右的泥盆纪地层中才肯定其存在[31]。从施秉云台山的现代藻类的组成中, 色球藻纲和藻殖段纲丰富, 尤其个体微小的球状蓝藻类占比高达54.23%, 亦进一步佐证了白云岩喀斯特是中寒武世古老产物。伍孟银等报道指出黔东南寒武纪地层中发现的具有水生植物向陆生植物过渡的初期阶段的结构特征的平行藻属Parallelphyton, 类似现生的被认为可能是高等植物的祖先的费氏藻属Fritschiella[32]。因此, 施秉云台山现存的费氏藻等藻类可能蕴藏着从寒武纪至今地质历史演化的痕迹, 而赤水丹霞现存藻类, 如真枝藻属物种与追溯到了泥盆纪的化石种的亲缘关系的探讨亦将可能成为明确赤水丹霞地貌发育起始时期的重要证据, 此外, 两地不同沉积相下现代藻类的研究对探明藻类演化及早期后生植物分类、分布和演化的研究具有重要意义。
3.2 两世界自然遗产地的地层岩性与陆生藻类种组特征岩性作为地貌发育、演化的物质基础, 在各种地貌形态成因分析时起着重要的作用[33]。岩性反映岩石特征属性, 包括成分、结构、特殊矿物等, 直接或间接地影响土壤的成分及其上植被类型与分布。大地构造位置上, 施秉云台山区域属扬子陆块范畴, 发育一套以薄层白云岩为主体的碳酸盐岩组合的岩溶构造[25], 独特的地上地下二元空间结构, 地表储水困难, 生境具有石漠化、土壤贫瘠、含钙量高等特点, 植被因此具有旱生性、岩生性等不同于其它地区植被的特点。从施秉云台山的陆生藻类组成上, 充分体现了对这一岩性特征所赋予的石生干旱贫瘠等环境的适应。主要表现在蓝藻的绝对优势上, 囊括色球藻纲两大目, 其中最为原始的色球藻科是其第一大优势科, 该科物种数占蓝藻门总记录物种数的一半以上。色球藻科物种的球状单细胞或多细胞胶群体, 结构简单、具有不同程度的胶鞘是其适应干旱的重要机制。另报道指出白云岩的δ13C值除与循环环境条件有关外, 主要与藻类相关[25]。白云岩孔裂隙结构特性与岩面分布植物含N量具相关性[34]。施秉云台山以较进化的具异形胞的丝状固氮蓝藻伪枝藻科、念珠藻科的优势, 反映了对碳氮营养缺乏的贫瘠生境的适应, 同时改善其殖居微生境并与岩性成分、结构等形成明显响应。郑云普等[35]对念珠藻属研究结果显示pH值、Ca2+浓度、Mg2+浓度对其生长影响极显著。袁蕾等[36]对贵州产念珠藻的矿质元素测定, 揭示了念珠藻的喜钙性。念珠藻科优势较好地指示了白云岩喀斯特的富钙、镁的岩性[33]。此外, 集胞藻属(Synechocystis)和伪枝藻属(Scytonema)对碳酸盐沉积也具有某种程度的特殊性[37]。白云岩土壤的微碱性[38], 促使了喜中偏碱性环境的蓝藻的发育和生长。施秉云台山丰富的陆生藻类的种组反映了地层岩性。
赤水丹霞研究区属扬子区四川盆地分区, 产状近水平的红色厚层状长石石英砂岩层被普遍认为是赤水地区丹霞地貌发育最主要的物质基础。岩体内的盐类物质成分、含量与岩体强度等都会受到岩面生物影响[39], 如Büdel等研究指出砂岩的硅酸与其定殖蓝藻代谢产生的生物碱发生反应加速岩表破坏, 主要为粘杆藻属、粘球藻属种类[40]。由此推测, 本研究中赤水丹霞包含该两属并且以粘球藻属为优势的原因可能与某些矿物成分有关, 而不同其在白云岩“喀斯特干旱”生存对策及适应机制。虽赤水地区年降水量与施秉云台山相近, 但由于砂岩、泥岩的透水性差, 地表水系极为发达, 因此具胶鞘的陆生蓝藻类少于施秉云台山, 且胶鞘也因水分较多而相较稀薄。赤水红层中的CaO及CaCO3的平均含量较少, 远低于7.05%、9.33%[41], 不到施秉云台山CaO平均含量(30.87%)[42]的1/4, 从岩石钙含量差异上解释了本研究中赤水丹霞的喜钙藻类如念珠藻属的种类少于施秉云台山的现象。在赤水炎热潮湿的气候条件下, 大量铁、铝、硅质在酸性水溶液中发生胶合形成块状红土, 尤以赤铁矿居多[43], Fe含量丰富可达6.8%左右[44], 而施秉云台山较低, 仅为0.09%—0.36%[42]。许多实验发现铁、锰等对藻类的生长表现出“低促高抑”现象, 低质量浓度元素更易于藻类生长[45], 这亦可能是施秉云台山藻类较赤水丰富的原因之一。此外, 通常为高pH指示生物的刚毛藻属在施秉云台山有分布而在赤水丹霞中未调查到, 也反映了两地岩性不同造就的生境的酸碱性差异。因此, 岩石微量元素的成分组成差异、pH值的高低都直接或间接影响着藻细胞内部生理生化反应, 成为了影响藻类物种组成及分布原因之一。
3.3 两世界自然遗产地的地理位置与陆生藻类物种相似性从全球的陆生藻类分布来看, 不同地区藻属的相似性介于40%—60%之间[46], 其中有研究指出, 澳大利亚与非洲和南美洲共有的藻类种类比世界其他任何地方都要多, 从藻类上佐证古冈瓦纳关系。杨瑞东等对布于北美、西伯利亚、欧洲和中国贵州、云南等地早、中寒武世地层中的宏观藻群化石分析并建立的古生物地理分区模式, 指出相同的古纬度范围内的藻类组合特征是相同的, 较好地解释不同大陆现代藻类的相似性[47]。贵州地史也表明其早期阶段与中国西北地区甚至大洋洲可能有许多亲缘关系[48]。胡春香、张丙昌等报道的西北地区的优势藻主要为色球藻属、颤藻属物种[49-50], 与本研究结果一致, 支持了这一观点。施秉云台山和赤水丹霞两世界自然遗产地虽分属贵州北部与东南部, 但处于同一纬度带, 且两地的海拔、温度、年均降水量相似, 因此, 陆生藻类属级水平的相似性达到44%左右。进一步从种水平上的聚类结果分析贵州东南部的施秉云台山与北部的赤水丹霞聚为一类, 贵州东南部的雷公山与西南的马岭河聚为一类的原因, 是由于雷公山为苗岭主峰, 追溯苗岭形成的地质历史时间为距今约248万年更新世, 晚于施秉云台山白云岩喀斯特及赤水丹霞地貌的形成时间, 并且有证据证明苗岭的形成是由于长江和珠江水系的向西溯源侵蚀和切割所致[51], 雷公山成为了贵州境内长江和珠江水系的分水岭高地, 以北为长江水系, 以南为珠江水系。施秉云台山和赤水丹霞两世界自然遗产地位于苗岭以北长江流域, 这就解释了聚类分析的结果, 但还有待从水生藻类到陆生藻类演变的角度寻找更多的支撑证据。
4 结论(1) 在环境因子相似情况下, 两世界自然遗产地陆生藻类均表现为蓝藻为主的蓝-绿-硅的组成结构, 具不同程度胶鞘的色球藻科和自由运动特性的颤藻科作为共同的优势科, 是对两地干旱、干热环境的形态和生理性响应;施秉云台山物种丰富度高于赤水丹霞, 与白云岩土壤的微碱性促藻、红层土壤的富铁锰性抑藻效应有关;施秉云台山具有伪枝藻科、念珠藻科固氮蓝藻的优势则进一步反映了白云岩喀斯特的富钙镁贫营养的环境特性;赤水丹霞粘球藻属物种更为丰富, 除与干热气候有关外, 还与砂岩的硅酸矿物有关。两地物种组成上的差异, 主要是由于地质历史构造所造成的喀斯特白云岩和丹霞地貌的石英砂岩岩性差异所致。
(2) 现存藻类的差异与对应化石藻的发现为两地地貌发育时间提供了佐证;施秉云台山现存陆生藻类与对古地理时期的海洋沉积微体藻类研究报道的科属组成相似, 很可能解释说明藻类从水环境到陆生环境的演变规律, 但需进一步从分子系统学开展研究。
(3) 两世界自然遗产地陆生藻类属的相似性系数为44.05%, 与全球其他地区报道结果属同一水平范围, 说明了藻类从水生到陆生的趋同进化;物种相似性不高, 反映了藻类对地理位置空间分异造成的不同陆生环境的适应性, 需从比较形态学、生理学及基因组学等方面进一步探讨其适应性机制。
致谢: 感谢西澳大学Pan Ruliang教授对写作的帮助。| [1] |
舒江平, 刘莉, 沈慧, 戴锡玲, 王全喜, 严岳鸿. 基于系统基因组学分析揭示早期陆生植物的复杂网状进化关系. 生物多样性, 2017, 25(6): 675-682. |
| [2] |
Knoll A H, Barghoorn E S, Golubić S. Paleopleurocapsa Wopfnerii gen. et sp. nov.: a late Precambrian alga and its modern counterpart. Proceedings of the National Academy of Sciences of the United States of America, 1975, 72(7): 2488-2492. DOI:10.1073/pnas.72.7.2488 |
| [3] |
茅绍智, 黄奇瑜, 雷作淇. 早中新世沟鞭藻植物地理区系. 现代地质, 2000, 14(3): 273-280. DOI:10.3969/j.issn.1000-8527.2000.03.006 |
| [4] |
烟晓玲. 太行山中段古元古代东冶组藻礁特征及其意义[D]. 石家庄: 河北地质大学, 2019.
|
| [5] |
王教元, 陈光杰, 康文刚, 胡葵, 陈小林, 吴飞红, 朱庆生, 冯钟. 基于硅藻功能群特征揭示海西海近现代生态环境变化. 应用生态学报, 2018, 29(9): 3120-3130. |
| [6] |
吴亚生, 蒋子文, 姜红霞. 湖南慈利晚二叠世松藻类化石古生态. 古生物学报, 2020, 59(2): 198-214. |
| [7] |
穆西南. 贵州西部上二叠统的钙藻化石. 古生物学报, 1981, 20(1): 33-49, 104-107. |
| [8] |
刘志礼. 化石藻类学导论. 北京: 高等教育出版社, 1990.
|
| [9] |
陈武, 肖加飞, 谯文浪, 李艳桃, 唐佐其, 刘凌云. 贵州施秉喀斯特世界自然遗产地地质遗迹类型和特征. 贵州科学, 2020, 38(1): 62-70. DOI:10.3969/j.issn.1003-6563.2020.01.012 |
| [10] |
陈奕彤, 熊康宁, 肖时珍, 肖华. 中国丹霞赤水世界自然遗产景观格局时空演变及驱动因素分析. 水土保持研究, 2018, 25(6): 314-321. |
| [11] |
郭福生, 陈留勤, 严兆彬, 刘富军, 潘志新, 张炜强, 胡海平. 丹霞地貌定义、分类及丹霞作用研究. 地质学报, 2020, 94(2): 361-374. DOI:10.3969/j.issn.0001-5717.2020.02.002 |
| [12] |
肖时珍, 容丽, 肖华, 蓝家程, 罗俊杰, 方俊, 陈奕彤. 中国丹霞赤水世界自然遗产地监测体系构建. 贵州师范大学学报: 自然科学版, 2019, 37(6): 8-19. |
| [13] |
李芹, 容丽, 王敏. 地形对喀斯特山地植物物种多样性及分布格局的影响. 水土保持通报, 2019, 39(6): 27-34. |
| [14] |
汤晓辛, 张潮, 商传禹. 世界遗产地施秉云台山白云岩喀斯特野生种子植物的区系. 分子植物育种, 2017, 15(5): 1986-1993. |
| [15] |
李晓娜, 熊康宁, 陈浒, 容丽, 向刚, 郭治友, 周江. 黔东南施秉白云岩喀斯特地区生物多样性与世界遗产价值. 贵州师范大学学报: 自然科学版, 2010, 28(3): 13-18. DOI:10.3969/j.issn.1004-5570.2010.03.004 |
| [16] |
蔡艳, 罗小菊, 王晓双, 王雁青, 王帅, 刘映良. 黔东南州白云岩喀斯特区植物群落演化特征. 西北植物学报, 2019, 39(11): 2064-2071. |
| [17] |
杨菊, 熊康宁, 肖时珍. 赤水丹霞地貌区植被覆盖度时空变化分析. 人民长江, 2018, 49(18): 38-44. |
| [18] |
彭涛, 李飞, 梁盛, 何琴琴, 张廷跃, 张潮, 麻俊虎. 贵州赤水桫椤国家级自然保护区藓类植物区系分析. 分子植物育种, 2018, 16(22): 7541-7549. |
| [19] |
蒋志刚, 纪力强. 鸟兽物种多样性测度的G-F指数方法. 生物多样性, 1999, 7(3): 220-225. DOI:10.3321/j.issn:1005-0094.1999.03.010 |
| [20] |
王荷生. 植物区系地理. 北京: 科学出版社, 1992.
|
| [21] |
吴征镒, 王荷生. 中国自然地理-植物地理(上). 北京: 科学出版社, 1983.
|
| [22] |
Ohtomo Y, Kakegawa T, Ishida A, Nagase T, Rosing M T. Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks. Nature Geoscience, 2014, 7(1): 25-28. DOI:10.1038/ngeo2025 |
| [23] |
Butterfield N J. Plankton ecology and the Proterozoic-Phanerozoic transition. Paleobiology, 1997, 23(2): 247-262. DOI:10.1017/S009483730001681X |
| [24] |
郑书粲. 中上扬子寒武纪早期有机质来源与类型及微体古生物组合对有机质保存的影响[D]. 武汉: 中国地质大学, 2019.
|
| [25] |
谯文浪, 肖加飞, 陈武, 李艳桃, 唐佐其, 马义波. 贵州施秉云台山白云岩类型、碳氧同位素组成及沉积环境. 地球与环境, 2014, 42(3): 347-354. |
| [26] |
Yang R D, Mao J R, Zhao Y L. New macroalgal fossils from middle Cambrian Kaili Biota in Guizhou Province, China. Acta Botanica Sinica, 2001, 43(7): 742-749. |
| [27] |
Peng H. China Danxia. Singapore: Springer, 2020.
|
| [28] |
李廷勇, 王建力. 中国的红层及发育的地貌类型. 四川师范大学学报: 自然科学版, 2002, 25(4): 427-431. DOI:10.3969/j.issn.1001-8395.2002.04.029 |
| [29] |
保广普, 刘春娥, 黄广文. 青海丹霞地貌的分布、特征及演化. 西北地质, 2019, 52(3): 199-208. |
| [30] |
王振, 王克勇. 从晚白垩世轮藻化石的发现论黔中红层的划分及相关的构造运动. 古生物学报, 1985, 24(5): 492-502. |
| [31] |
朱浩然. 中国淡水藻志第2卷色球藻纲. 北京: 科学出版社, 1991.
|
| [32] |
伍孟银, 赵元龙, 童金南, 杨瑞东. 贵州凯里生物群中的宏观藻类新材料. 中国科学: 地球科学, 2010, 40(11): 1510-1517. |
| [33] |
张乾柱, 刘子琦, 罗井升, 白雪梅, 叶正明. 岩石地球化学特征对喀斯特峡谷形态与分布影响分析——以贵州施秉喀斯特世界自然遗产预选地为例. 西南大学学报: 自然科学版, 2012, 34(6): 114-120. |
| [34] |
王美权, 黄宗胜. 木本植物对喀斯特石质生境岩石形态结构的适应性. 生态学报, 2018, 38(21): 7749-7761. |
| [35] |
郑云普, 张丙昌, 赵建成, 张元明. 不同理化因子对分离于荒漠生物结皮中念珠藻生长的影响. 河北师范大学学报: 自然科学版, 2009, 33(4): 531-537, 542-542. |
| [36] |
袁蕾, 谭娅, 胡泽刚, 侯云燕, 晏妮. 地木耳的营养成分与铅汞含量分析. 食品安全质量检测学报, 2020, 11(4): 1145-1149. |
| [37] |
张京梅. 环境因子对蓝藻的生长及其钙化速率影响的研究[D]. 贵阳: 贵州师范大学, 2007.
|
| [38] |
邹军, 喻理飞, 李媛媛, 黄宗胜. 喀斯特区不同岩性发育的土壤上植物-枯落物-土壤系统δ13C值变化特征. 水土保持学报, 2020, 34(1): 186-194. |
| [39] |
彭华, 潘志新, 闫罗彬, Simonson S. 国内外红层与丹霞地貌研究述评. 地理学报, 2013, 68(9): 1170-1181. |
| [40] |
Büdel B, Weber B, Kühl M, Pfanz H, Sültemeyer D, Wessels D. Reshaping of sandstone surfaces by cryptoendolithic cyanobacteria: bioalkalization causes chemical weathering in arid landscapes. Geobiology, 2004, 2(4): 261-268. DOI:10.1111/j.1472-4677.2004.00040.x |
| [41] |
黄进. 赤水丹霞地貌. 北京: 科学出版社, 2015.
|
| [42] |
谯文浪, 肖加飞, 陈武, 李艳桃, 刘凌云, 马义波. 贵州施秉云台山白云岩地球化学特征及成因探讨. 沉积与特提斯地质, 2015, 35(3): 94-101. |
| [43] |
刘江龙. 中国东南部丹霞地貌形成机理及其地学效应研究[D]. 长沙: 中南大学, 2009.
|
| [44] |
龚子同, 张甘霖, 陈志诚. 土壤发生与系统分类. 北京: 科学出版社, 2007.
|
| [45] |
孔赟, 邹培, 宋黎明, 王子, 戚姣琴, 朱亮, 徐向阳. 铁对藻类生长及藻毒素合成影响研究进展. 应用生态学报, 2014, 25(5): 1533-1540. |
| [46] |
Belnap J, Lange O L. Biological Soil Crusts: Structure, Function, and Management. Berlin Heidelberg: Springer, 2003.
|
| [47] |
杨瑞东, 毛家仁, 张位华, 姜立君. 全球早、中寒武世藻类生物地理分区初探. 古地理学报, 2002, 4(2): 12-18. DOI:10.3969/j.issn.1671-1505.2002.02.002 |
| [48] |
陈毓遂. 贵州地质演化的基本轮廓. 贵州地质, 1984, 1(1): 65-78. |
| [49] |
胡春香, 张斌才, 马红樱, 刘永定, 张德禄. 兰州北山生物结皮中陆生藻种类组成与群落结构. 西北师范大学学报: 自然科学版, 2003, 39(1): 59-63. |
| [50] |
张丙昌, 张元明, 赵建成, 张茹春. 准噶尔盆地古尔班通古特沙漠生物结皮蓝藻研究. 地理与地理信息科学, 2005, 21(5): 107-109. |
| [51] |
周启永, 陈佩英. 贵州及邻区晚新生代岩相古地理演化. 贵州地质, 1993, 10(3): 201-207. |