 2021, Vol. 41
2021, Vol. 41文章信息
- 高明磊, 满秀玲, 段北星
- GAO Minglei, MAN Xiuling, DUAN Beixing
- 林下植被和凋落物对寒温带森林生长季土壤CH4通量的影响
- Effects of understory vegetation and litter on soil CH4 flux during growing season in cold temperate forest in China
- 生态学报. 2021, 41(24): 9886-9897
- Acta Ecologica Sinica. 2021, 41(24): 9886-9897
- http://dx.doi.org/10.5846/stxb202012203232
-
文章历史
- 收稿日期: 2020-12-20
- 网络出版日期: 2021-08-04





2. 东北林业大学森林生态系统可持续经营教育部重点实验室, 哈尔滨 150040
2. Key Laboratory of Sustainable Forest Ecosystem Management of Ministry of Education, Northeast Forestry University, Harbin 150040, China
CH4是大气中主要的温室气体之一, 近几十年来, 其浓度还在逐年增加, 年增长速度达到了0.8%[1]。尽管大气中CH4的浓度远低于CO2的浓度, 但是CH4在大气中停留时间较长, 具有更强的远红外吸收能力[2], 因此在百年尺度上, CH4增温潜势大约是CO2的25倍[3]。森林土壤则是CH4重要的汇, 每年吸收CH4的量约为(9.16±3.84)Tg[4], 这对调节CH4在大气中的含量具有重要意义。此外, 林下植被和凋落物的不同管理方式会影响土壤理化性质[5]、土壤养分[6]和土壤微生物群落[7]等, 从而影响碳通量。枯枝落叶的数量、成分以及分解速率会对林地土壤中微生物的数量和活性、植物养分的供应以及水热条件产生影响, 因此凋落物会对土壤C循环产生影响[8], 但影响程度不尽相同。研究发现北亚热带造林后, 去除凋落物对CH4通量没有显著影响[9], 但在半干旱的科尔沁地区人工杨树林[10]和奥利地山毛榉林[11]研究发现, 去除凋落物后CH4吸收值分别提高33.51%和16.00%。同样, 去除林下植被不仅能改善土壤有机物的输入, 而且能通过改变小气候和土壤理化性质来影响C循环, 从而对森林土壤CH4吸收产生重要影响[12]。在南亚热带研究表明, 剔除林下灌草促进了尾叶桉林[13]和厚荚相思林[12]CH4的吸收;中亚热带也发现相似的结果, 剔除杂草后山核桃林地土壤CH4的年吸收量为116.32 kg hm-2 a-1, 高于留养杂草的71.30 kg hm-2 a-1[14]。去除林下植被后会使土壤湿度降低, 地表土壤的透气性增大, 激活甲烷氧化菌等, 从而提高了林地土壤对CH4的吸收。目前林下植被和凋落物对CH4通量的影响还比较局限, 首先, 只是考虑单一营林措施对其通量的研究, 尚未进一步探索二者交互作用对其通量的影响;再者, 对人工林研究较多, 对天然林的研究相对较少。最后, 国内对这方面研究多集中在亚热带[9, 12-14]和温带[10], 而对于寒温带地区天然林的研究还较少。
大兴安岭林区是我国最大的原始林区, 也是我国唯一的高纬度寒温带林区[15]。作为我国东北平原重要的生态屏障, 大兴安岭森林生态系统在国家生态功能区中发挥着重要的碳平衡功能。该地区也是全球气候变化最敏感的区域之一[16]。研究表明, 我国寒温带地区的森林土壤多表现为CH4的汇[17-19], 然而在森林经营管理过程中不同的营林方式对土壤CH4通量有何影响, 目前尚不清楚。为了更好地研究我国寒温带冻土区森林经营方式对林地土壤CH4通量的影响, 本研究选择白桦林、山杨林、樟子松林和兴安落叶松林4种天然林类型为研究对象, 在林下进行4种营林方式处理(自然状态、去除凋落物、去除林下植被以及林下植被和凋落物都去除), 对其CH4通量特征进行分析探究, 为大兴安岭地区森林生态系统的经营管理和土壤温室气体研究提供科学依据和理论参考。
1 材料与方法 1.1 研究区概况研究区位于大兴安岭地区漠河县黑龙江漠河森林生态系统国家定位观测研究站, 地理坐标为122°06′—122°27′E, 53°17′—53°30′N, 海拔高度209—378 m, 该地区属寒温带大陆性季风气候, 冬季寒冷漫长, 夏季温热多雨, 全年平均气温在-4.9℃, 多年平均降水量在350—500 mm之间, 且降水多集中在7—8月, 全年无霜期80—90 d。该地区也是我国多年冻土的主要分布区, 地带性土壤为棕色针叶林土, 地带性植被是兴安落叶松(Larix gmelinii)林, 此外还有樟子松(Pinus sylvestris var. mongolica)林、白桦(Betula platyphylla)林和山杨(Populus davidiana)林等森林植被类型。林下植被主要有兴安杜鹃(Rhododendron dauricum)、杜香(Ledum palustre)、笃斯越桔(Vaccinium uliginosum)等。
1.2 样地设置与样品采集2019年4月, 经过实地考察后选择白桦林、山杨林、樟子松林和兴安落叶松林(见表 1)4种森林类型为研究对象, 每种林型设置3块20 m×30 m的样地, 每一块样地进行4种处理, 即自然状态、去除凋落物、去除林下植被以及林下植被和凋落物都去除, 每一处理样方面积为1.5 m×1.5 m。林下植被去除是将植被从根部将其剪断, 凋落物去除是将未分解层和半分解层清理掉。五月初第一次完全处理后每周都要清理一次, 且每次取气前3 d完成准备工作。每种处理均设置3个静态箱(包括顶箱和底座两部分), 静态箱底座(50 cm×50 cm×20 cm)于4月份设置, 底座应插入地表 5 cm以下, 并用泥土将底座下端密实且固定。静态箱顶箱(50 cm×50 cm×50 cm)外部有保温板, 箱内装有小风扇和温度计。采集样品时, 底座和顶箱之间的水槽要加水密封。
| 林型 Forest type |
坡向 Slope aspect |
坡度 Gradient/(°) |
林龄 Age/a |
平均胸径 Mean DBH/cm |
平均树高 Height of tree/m |
郁闭度 Canopy density |
主要林下植被 Understory species compositio |
| 白桦林 Betula platyphylla forest |
NE | 6 | 44±5.6 | 11.5±5.5 | 13.1±4.1 | 0.9 | 1、3、4、6 |
| 山杨林 Populus davidiana forest |
NE | 7 | 33±4.3 | 16.2±6.3 | 16.5±6.8 | 0.7 | 6、7、8 |
| 樟子松林 Pinus sylvestris var. mongolica forest |
NE | 5 | 96±7.2 | 27.3±5.4 | 21.6±4.5 | 0.6 | 1、3、5 |
| 兴安落叶松林 Larix gmelini forest |
NE | 6 | 78±5.9 | 14.1±6.2 | 19.2±5.6 | 0.8 | 1、2、3、4 |
| 1: 兴安杜鹃(Rhododendron dauricum);2: 杜香(Ledum palustre.);3: 越桔(Vaccinium vitis-idaea);4: 笃斯越桔(Vacciniumu liginosum.);5: 羽节蕨(Gymnocarpium jessoense);6: 红花鹿蹄草(Pyrolaincarnata);7: 舞鹤草(Maianthemum bifolium);8: 东方草莓(Fragaria orientalis);NE: 东北方向; DBH: 胸径Diameter at breast height表中林龄、树高和胸径为平均值±标准差 | |||||||
从2019年5月到9月, 选择晴天上午的9:00—12:00(国际通用时间)采用压力取气法采集气体样品, 并以此时段内的排放速率代表全天平均通量[20-21]。取气是每间隔15 min采集一次, 共4次, 总时间45 min。每次取样50mL左右分别注入大连德霖气体包装公司生产的铝箔采样袋。每次取样的同时记录抽气时间、箱内外温度、5 cm和10 cm的土壤温度和湿度。根据天气情况, 每20 d左右取1次气体样品。所有气体样品尽快带回实验室, 并于一周内用气相色谱仪(赛默飞trace1300)分析测定CH4气体浓度。
每次取气的同时在静态箱底座外围进行土样的采取(避开取气区), 随机选3个点, 分别取0—5 cm和5—10 cm的土壤等质量混合, 带回实验室然后挑去草根、石块, 然后采用国家标准方法对土壤理化性质测定。土壤pH采用玻璃电极法(土水比1 ∶ 2.5)测定;土壤有机碳通过元素分析仪(Vario-TOC, Germany-Elementar)测定。土壤全氮、铵态氮和硝态氮使用连续流动分析仪(AA3, Germany-SEAL)测定, 自然状态土壤基本理化性质见表 2。
| 林型 Forest type |
土层深度 Soil layer/cm |
pH | 有机碳 Total soil organic carbon/(g/kg) |
全氮 Total soil nitrogen/(g/kg) |
铵态氮 NH4+-N/(mg/kg) |
硝态氮 NO3--H/(mg/kg) |
| 白桦林 | 0 —5 | 5.34±0.15Aab | 70.99±1.45Aa | 2.97±0.14Ab | 2.97±0.54Aa | 0.37±0.08Aa |
| Betula platyphylla forest | 5—10 | 5.89±0.21Aa | 29.45±0.73Ba | 1.68±0.09Bb | 1.79±0.22Ba | 0.30±0.06Bb |
| 山杨林 | 0—5 | 5.21±0.18Aab | 57.76±1.98Ab | 3.78±0.33Aa | 1.42±0.33Ac | 0.36±0.07Aa |
| Populus davidiana forest | 5—10 | 5.45±0.31Aab | 34.15±1.01Ba | 2.22±0.12Ba | 1.37±0.30Ab | 0.34±0.06Aa |
| 樟子松林 | 0—5 | 5.66±0.27Aa | 46.71±1.12Ac | 2.24±0.19Ac | 1.67±0.51Abc | 0.38±0.10Aa |
| Pinus sylvestris var mongolica forest | 5—10 | 5.71±0.33Aab | 20.11±0.98Bb | 1.02±0.10Bc | 1.12±0.27Bc | 0.35±0.03Aa |
| 兴安落叶松林 | 0—5 | 4.92±0.19Ab | 40.34±1.10Ac | 2.52±0.27Ac | 1.91±0.48Ab | 0.38±0.05Aa |
| Larix gmelini forest | 5—10 | 5.11±0.21Ab | 11.52±0.34Bc | 0.91±0.07Bc | 1.65±0.34Ba | 0.35±0.05Aa |
| 表中数值为平均值±标准差;不同大写字母表示相同林型不同土层间各指标差异达到显著水平(P<0.05);不同小写字母表示相同土层不同林型间各指标差异达到显著水平(P<0.05) | ||||||
利用以下公式进行CH4通量计算:
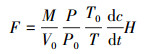
|
式中:F为气体通量(μg m-2 h-1), M为气体的摩尔质量(g/mol), V0、P0和T0分别为标准状况下气体摩尔体积(m3/mol)、大气压(Pa)和温度(K), P和T分别为取样时的实际气压(Pa)和温度(K), dc/dt为气体浓度随时间变化的回归曲线斜率(μmol mol-1h-1), H为箱内部地表到箱顶的高度(m)。当F为正值时表示排放, F为负值时表示吸收。
1.4 数据处理采用SPSS 25.0和Excel 2010对数据进行分析, 采用Duncan多种比较分析不同林型不同处理的CH4通量均值差异, 用Pearson法对CH4通量与环境因子进行相关性分析, 用Origin 2018进行作图。图中和表中的数据为平均值±标准差。
2 结果与分析 2.1 4种林型不同处理土壤温湿度特征在整个观测期内, 4种林型不同处理土壤5 cm和10 cm温湿度变化规律基本一致, 白桦林、山杨林和樟子松林的土壤温湿度在不同处理后差异性不显著(P>0.05), 但兴安落叶松林的土壤温湿度不同处理间差异性显著(P<0.05), 同一处理不同月份之间土壤温湿度组间差异性显著(P<0.05)。4种林型不同处理土壤5 cm和10 cm温湿度详见表 3。
| 林型 Forest type |
对比处理 Treatment |
5 cm Ts/℃ | 10 cm Ts/℃ | 5 cm VWC/% | 10 cm VWC/% | |||||||
| 范围 | 均值 | 范围 | 均值 | 范围 | 均值 | 范围 | 均值 | |||||
| 白桦林 | CK | 2.1—13.1 | 9.8 | 1.2—11.6 | 8.4 | 5.1—15.8 | 9.0 | 7.0—24.8 | 14.3 | |||
| Betula platyphylla forest | DL | 3.9—15.0 | 10.6 | 2.3—12.0 | 9.0 | 4.5—13.4 | 8.3 | 6.8—21.5 | 13.1 | |||
| DP | 2.6—13.8 | 10.4 | 2.2—12.3 | 9.2 | 4.5—13.1 | 8.5 | 6.2—24.3 | 13.7 | ||||
| DD | 3.4—13.3 | 10.7 | 2.4—12.5 | 9.2 | 4.4—13.3 | 8.6 | 6.0—26.9 | 13.7 | ||||
| 山杨林 | CK | 3.4—14.4 | 10.4 | 1.3—11.3 | 8.3 | 5.3—15.7 | 11.2 | 9.1—26.6 | 19.3 | |||
| Populus davidiana forest | DL | 4.4—13.9 | 10.6 | 2.6—11.6 | 8.9 | 6.2—14.2 | 10.6 | 10.1—23.6 | 18.5 | |||
| DP | 4.3—14.5 | 11.0 | 2.3—11.5 | 8.6 | 6.2—15.1 | 10.6 | 9.8—22.0 | 17.4 | ||||
| DD | 6.1—14.6 | 11.5 | 3.8—12.3 | 9.5 | 5.6—13.1 | 9.5 | 9.6—22.0 | 16.2 | ||||
| 樟子松林 | CK | 4.7—15.3 | 11.3 | 3.1—14.2 | 10.1 | 5.0—15.7 | 9.6 | 8.4—23.6 | 15.4 | |||
| Pinus sylvestris var.mongolica forest | DL | 3.9—17.2 | 11.9 | 3.3—14.4 | 10.4 | 5.6—13.3 | 8.9 | 7.3—21.6 | 14.7 | |||
| DP | 5.3—15.6 | 11.6 | 3.0—14.8 | 10.4 | 4.4—15.1 | 8.9 | 8.2—22.0 | 14.9 | ||||
| DD | 3.6—17.4 | 11.8 | 3.6—15.1 | 10.8 | 5.1—12.7 | 9.2 | 8.6—19.6 | 14.6 | ||||
| 兴安落叶松林 | CK | 3.0—9.4 | 7.4 | 0.9—8.2 | 5.8 | 3.1—15.7 | 8.0 | 4.8—26.6 | 14.2 | |||
| Larix gmelini forest | DL | 3.8—10.8 | 8.6 | 1.9—9.4 | 7.0 | 4.6—10.2 | 7.1 | 7.2—15.3 | 12.8 | |||
| DP | 5.4—13.7 | 10.0 | 3.0—10.7 | 8.2 | 4.3—10.1 | 6.7 | 8.7—22.0 | 12.2 | ||||
| DD | 6.0—12.7 | 10.0 | 3.9—10.7 | 8.1 | 3.4—10.1 | 6.5 | 6.1—18.0 | 11.8 | ||||
| Ts: 土壤温度Soil temperature;VWC: 土壤湿度Soil moisture; CK: 自然状态Natural state;DL: 去除凋落物Removing litter; DP: 去除林下植被Removing understory vegetation;DD: 去除林下植被和凋落物Removing understory vegetation and litter | ||||||||||||
由图 1可知, 寒温带4种天然林土壤CH4通量均为负值, 说明这4种林型均表现为CH4的汇。4种林型土壤CH4通量在5—9月均表现出明显的单峰曲线变化趋势, 峰值出现在7月下旬或8月中旬, 且兴安落叶松林土壤CH4吸收通量峰值(-77.17±21.61)μg m-2 h-1显著小于其他三种林型(P<0.05)。兴安落叶松林的CH4吸收通量从生长季初期的最小值(-39.42±8.28)μg m-2 h-1上升至吸收峰值(-77.17±21.61)μg m-2 h-1, 升幅为95.77%, 然后从吸收峰值下降到生长季末期(-20.13±4.23)μg m-2 h-1, 降幅为73.92%。白桦林则是和兴安落叶松林有着较为相似的变化趋势, 且在整个监测期内, 兴安落叶松林的CH4吸收通量显著小于白桦林(P<0.05)。樟子松林相比于其它三种林型变化幅度比较大, 其土壤CH4吸收通量从生长季初期的最小值(-10.54±2.21)μg m-2 h-1上升到吸收峰值(-110.75±32.18)μg m-2 h-1, 然后从吸收峰值下降到生长季末期(-30.63±6.12)μg m-2 h-1。另外生长季初期, 樟子松林的土壤CH4吸收通量显著小于其他三种林型(P<0.05);而生长季末期, 阔叶林的土壤CH4吸收通量显著大于针叶林(P<0.05)。4种林型CH4平均吸收通量表现为白桦林(-79.23±14.92)μg m-2 h-1>山杨林(-64.27±9.60)μg m-2 h-1>樟子松林(-62.54±15.48)μg m-2 h-1>兴安落叶松林(-48.73±12.26)μg m-2 h-1。兴安落叶松林土壤CH4平均吸收通量显著小于其他三种林型(P<0.05), 且总体而言, 阔叶林吸收通量大于针叶林。

|
| 图 1 4种林型生长季不同处理土壤CH4通量动态特征 Fig. 1 Dynamic characteristics of soil CH4 flux under different treatments in growth seasons of four forest types |
由图 1可知, 4种林型在去除凋落物后土壤CH4通量同为负值, 除了樟子松林土壤CH4吸收峰值(-108.88±21.78)μg m-2 h-1出现在8月, 其他三种林型的峰值均出现7月末。5月下旬4种林型CH4通量差异不显著(P>0.05), 但随着时间推移, 不同林型之间逐渐产生差异。6月之后, 兴安落叶松林土壤CH4吸收通量显著小于白桦林和樟子松林(P<0.05)。白桦林土壤CH4通量从生长季初期的(-21.36±4.27)μg m-2 h-1达到吸收峰值(-114.77±16.34)μg m-2 h-1, 变化幅度高达4.37倍, 但到生长季末期CH4通量为(-79.00±10.76)μg m-2 h-1, 变幅只有31.16%。樟子松林和山杨林在整个监测期过程中, 除了7月下旬, 其他时间差异性不显著(P>0.05)。4种林型在去除凋落物后土壤CH4平均吸收通量表现为白桦林(-81.58±12.18)μg m-2 h-1>山杨林(-70.58±8.01)μg m-2 h-1>樟子松林(-70.14±11.84)μg m-2 h-1>兴安落叶松林(-49.76±12.87)μg m-2 h-1, 阔叶林大于针叶林。
2.4 生长季去除林下植被4种林型土壤CH4通量特征由图 1可知, 4种林型去除植被后土壤CH4通量均为负值, 林地土壤也表现为CH4的汇。4种林型土壤CH4通量呈现单峰变化趋势, 峰值都是出现在7月, 且兴安落叶松林土壤CH4吸收通量峰值(-73.37±3.66)μg m-2 h-1显著小于其他三种林型(P<0.05)。山杨林和兴安落叶松林均是在7月下旬达到吸收峰值, 且山杨林土壤CH4吸收通量显著大于兴安落叶松林(P<0.05)。而白桦林和樟子松林在到达峰值前后(5—7月), 变化趋势较为一致, 差异性不显著(P>0.05)。4种林型在去除林下植被后CH4平均吸收通量表现为白桦林(-79.90±14.19)μg m-2 h-1>山杨林(-77.48±17.94) μg m-2 h-1>樟子松林(-68.27±13.41) μg m-2 h-1>兴安落叶松林(-51.25±7.30)μg m-2 h-1。分析发现, 兴安落叶松林分别与白桦林和山杨林差异性极显著(P<0.01)(表 4)。
| 林型Forest type | CK | DL | DP | DD |
| 白桦林Betula platyphylla forest | -79.23±14.92Aa | -81.58±12.18Aa | -79.90±14.19Aa | -82.35±8.80Aa |
| 山杨林Populus davidiana forest | -64.27±9.60Cb | -70.58±8.01BCb | -77.48±17.94Ba | -87.58±15.57Aa |
| 樟子松林Pinus sylvestris var. mongolica forest | -62.54±15.48Ab | -70.13±11.84Ab | -68.27±13.41Aab | -69.86±7.26Aab |
| 兴安落叶松林Larix gmelini forest | -48.73±12.26Bc | -49.76±12.87ABc | -51.25±7.30ABb | -55.38±7.83Ab |
| 表中数值为平均值±标准差;不同大写字母表示相同林型不同处理间各指标差异达到显著水平(P<0.05);不同小写字母表示相同处理不同林型间各指标差异达到显著水平(P<0.05) | ||||
由图 1可知, 4种林型林下植被和凋落物都去除后土壤CH4通量呈明显的单峰曲线, 除了兴安落叶松林的吸收通量峰值(-88.12±8.42)μg m-2 h-1出现在8月, 其他3种林型均出现在7月, 且兴安落叶松林的吸收通量显著小于其他3种林型(P<0.05)。在监测过程中, 4种林型土壤CH4平均吸收通量分别为山杨林(-87.58±15.57)μg m-2 h-1>白桦林(-82.35±8.80)μg m-2 h-1>樟子松林(-69.86±7.26)μg m-2 h-1>兴安落叶松林(-55.38±7.83)μg m-2 h-1。分析发现, 兴安落叶松林与白桦林差异性极显著(P<0.01)(表 4)。
2.6 4种林型不同处理间土壤CH4通量特征由图 2和表 4可知, 4种林型不同处理之后, 均呈现CH4通量吸收增加的趋势。白桦林在不同处理之后, 土壤CH4平均吸收通量表现为去除林下植被和凋落物(-82.35±8.80)μg m-2 h-1>去除凋落物(-81.58±12.18)μg m-2 h-1>去除林下植被(-79.90±14.19)μg m-2 h-1>自然状态(-79.23±14.92)μg m-2 h-1, 且不同处理相比于自然状态增加不显著(P>0.05)。山杨林在不同处理之后, 去除凋落物、去除林下植被以及去除林下植被和凋落物相比于自然状态(-64.27±9.60)μg m-2 h-1分别提高了9.81%、20.55%和36.26%, 其中后两者呈显著增加(P<0.05)。樟子松林在不同处理之后, 相比于自然状态(-62.54±15.48)μg m-2 h-1吸收增加最多的是去除凋落物(-70.13±11.84)μg m-2 h-1, 去除林下植被增加最小(-68.27±13.41)μg m-2 h-1, 但变化幅度都没有达到显著水平(P>0.05)。兴安落叶松林在不同处理之后, 土壤CH4平均吸收通量表现为去除林下植被和凋落物(-55.38±7.83)μg m-2 h-1>去除林下植被(-51.25±7.30)μg m-2 h-1>去除凋落物(-49.76±12.87)μg m-2 h-1>自然状态(-48.73±12.26)μg m-2 h-1, 且去除林下植被和凋落物相比于自然状态显著增加(P<0.05)。

|
| 图 2 不同林型不同处理生长季土壤CH4通量动态特征 Fig. 2 Dynamic characteristics of soil CH4 flux in different forest types with different treatments during the growing season |
不同处理方式均对森林土壤CH4通量有一定影响, 且不同林型不同处理之间存在差异。不同处理对白桦林和樟子松林土壤CH4通量影响不显著(P>0.05), 去除林下植被以及同时去除林下植被和凋落物对山杨林土壤CH4通量影响显著(P<0.05), 而同时去除林下植被和凋落物对兴安落叶松林土壤CH4通量影响显著(P<0.05)。由此可见, 凋落物去除与否对土壤CH4通量影响较小, 去除林下植被仅对山杨林土壤CH4通量影响显著, 而同时去除时对山杨林和兴安落叶松林土壤CH4通量影响显著。
2.7 CH4通量与环境因子的相关性由表 5可知, 白桦林的自然状态、去除凋落物以及去除林下植被土壤CH4通量与土壤温度呈极显著负相关(P<0.01), 而同时去除林下植被和凋落物则呈显著负相关(P<0.05);山杨林去除凋落物土壤CH4通量与土壤温度呈现显著负相关(P<0.05), 去除林下植被与土壤5 cm温度和空气温度呈现显著负相关(P<0.05);樟子松林只有去除两者后土壤CH4通量与土壤温度呈极显著负相关(P<0.01);兴安落叶松林的去除凋落物后土壤CH4通量与土壤温度呈极显著负相关(P<0.01), 自然状态和去除凋落物后土壤CH4通量与空气温度呈现显著负相关(P<0.05);4种林型不同处理均与土壤湿度之间相关不显著(P>0.05)。由此可见, 各处理条件下土壤CH4通量与土壤温度相关性密切, 而土壤湿度对土壤CH4通量的影响相对较小。
| 林型 Forest type |
处理 Treatment |
Ts | VWC | pH | 有机碳 Soil organic content |
全氮 Total nitrogen |
铵态氮 NH4+-N |
硝态氮 NO3--H |
|||||||||||||
| 5 cm | 10 cm | 5 cm | 10 cm | 0—5 cm | 5—10 cm | 0—5 cm | 5—10 cm | 0—5 cm | 5—10 cm | 0—5 cm | 5—10 cm | 0—5 cm | 5—10 cm | ||||||||
| 白桦林 | CK | -0.794** | -0.814** | 0.252 | 0.268 | -0.655** | -0.591** | -0.238 | -0.055 | -0.113 | 0.005 | 0.070 | 0.111 | 0.071 | -0.118 | ||||||
| Betula platyphylla forest | DL | -0.818** | -0.853** | -0.117 | 0.025 | -0.802** | -0.833** | -0.314 | -0.033 | -0.026 | 0.175 | 0.310 | 0.220 | 0.254 | 0.106 | ||||||
| DP | -0.631** | -0.664** | 0.272 | 0.261 | -0.277 | -0.318 | -0.051 | -0.007 | -0.127 | -0.029 | 0.068 | 0.067 | -0.134 | -0.377 | |||||||
| DD | -0.520* | -0.587* | 0.342 | 0.343 | 0.133 | 0.043 | 0.077 | 0.112 | -0.126 | -0.043 | -0.306 | 0.112 | -0.041 | -0.370 | |||||||
| 山杨林 | CK | -0.220 | -0.263 | -0.114 | 0.041 | 0.028 | 0.216 | 0.246 | 0.035 | 0.388 | 0.130 | 0.106 | 0.131 | 0.173 | -0.106 | ||||||
| Populus davidiana forest | DL | -0.609* | -0.602* | -0.448 | -0.475 | 0.074 | 0.242 | 0.530* | 0.223 | 0.675** | 0.192 | -0.054 | -0.115 | 0.060 | -0.487 | ||||||
| DP | -0.546* | -0.384 | 0.344 | 0.236 | 0.230 | 0.019 | 0.561* | 0.543* | 0.585* | 0.328 | -0.258 | -0.570* | -0.282 | -0.695** | |||||||
| DD | -0.566* | -0.524* | -0.009 | -0.020 | 0.390 | 0.337 | 0.275 | 0.255 | 0.579* | 0.359 | -0.045 | -0.414 | -0.227 | -0.554* | |||||||
| 樟子松林 | CK | -0.146 | -0.262 | -0.005 | -0.069 | 0.128 | 0.217 | 0.218 | 0.204 | 0.158 | 0.387 | 0.034 | -0.319 | -0.102 | -0.031 | ||||||
| Pinus sylvestris var. | DL | -0.645 | -0.613 | -0.344 | -0.316 | -0.090 | -0.248 | 0.094 | 0.134 | 0.258 | 0.568 | 0.246 | -0.273 | 0.369 | 0.449 | ||||||
| mongolica forest | DP | -0.317 | -0.204 | 0.461 | 0.377 | -0.036 | 0.029 | -0.543* | -0.180 | -0.499* | 0.093 | -0.340 | -0.296 | 0.036 | 0.174 | ||||||
| DD | -0.821** | -0.738** | 0.424 | 0.451 | -0.067 | -0.050 | -0.414 | 0.038 | -0.525* | 0.593* | -0.094 | -0.320 | 0.284 | 0.110 | |||||||
| 兴安落叶松林 | CK | -0.371 | -0.155 | 0.005 | 0.103 | -0.279 | -0.798** | 0.776** | -0.250 | 0.756** | -0.453 | -0.485 | -0.680** | -0.521 | -0.663** | ||||||
| Larix gmelini forest | DL | -0.883** | -0.855** | -0.594 | -0.451 | -0.419 | -0.726* | 0.338 | -0.112 | 0.411 | -0.114 | 0.268 | -0.631 | -0.539 | -0.804* | ||||||
| DP | -0.330 | -0.258 | -0.128 | -0.314 | -0.180 | 0.227 | 0.254 | 0.120 | 0.373 | -0.064 | -0.179 | -0.439 | -0.002 | -0.639* | |||||||
| DD | -0.338 | -0.428 | 0.202 | 0.245 | -0.424 | -0.469* | 0.263 | -0.083 | -0.060 | -0.300 | -0.287 | 0.101 | -0.061 | 0.149 | |||||||
| *P<0.05;**P<0.01 | |||||||||||||||||||||
白桦林自然状态和去除凋落物时土壤CH4通量与土壤pH呈极显著负相关(P<0.01), 其他均不相关。山杨林去除凋落物与5 cm有机碳显著正相关(P<0.05), 与5 cm全氮极显著正相关(P<0.01);去除林下植被与有机碳、5 cm全氮和10 cm铵态氮显著相关(P<0.05), 与10 cm硝态氮极显著负相关(P<0.01);两者都去除与5 cm全氮和10 cm硝态氮显著相关(P<0.05)。樟子松林去除林下植被与5 cm有机碳和全氮显著负相关(P<0.05), 两者都去除与全氮显著相关(P<0.05)。兴安落叶松林与5 cm有机碳和全氮极显著正相关(P<0.01), 与10 cm pH、铵态氮和硝态氮极显著负相关(P<0.01);去除凋落物和两者都去除与10 cm pH显著负相关(P<0.05);去除凋落物和去除林下植被与10 cm硝态氮显著负相关(P<0.05)。
3 讨论CH4通量主要取决于甲烷氧化菌消耗和产生菌生产的平衡作用[22]。森林土壤是大气CH4重要的汇, 而CH4吸收量多少则是受到多个因素的影响, 例如底物有效性、土壤温湿度、土壤pH、土壤中的养分以及植被类型等[23]。本研究结果表明, 4种林型土壤CH4吸收通量具有明显的单峰曲线变化趋势, 峰值出现在7月下旬或8月中旬, 这可能与本时期良好的水热条件有关。此外发现阔叶林土壤的CH4吸收通量高于针叶林, 这一研究结果与本地区[17, 19, 24]及其他地区[25]研究结果一致, 这一方面可能是阔叶林地土壤的通气性强于针叶林, 疏松的土壤有利于氧气输送, 提高了土壤中甲烷氧化菌的活性, 从而提高了土壤对CH4的吸收能力[17], 另一方面可能是针叶林的松针分解的单萜类物质会限制甲烷氧化菌的活性, 从而抑制土壤对CH4的吸收作用[26]。研究发现, 兴安落叶松林土壤温度均值小于其他三种林型(见表 3), 而土壤甲烷氧化菌的活性会随温度升高呈现指数变化趋势[27], 这可能也是兴安落叶松林土壤CH4吸收通量显著小于其他三种林型的主要原因(见表 3)。本研究结果与同纬度带的一些学者研究结果相似[18-19], 但低于华南地区的一些研究[12-14], 这可能与不同地区的气候条件、森林类型、林下植被种类以及凋落物质量和数量有关。
凋落物层是森林土壤的重要组成部分, 在地上和地下之间起到了桥接作用[28], 同时它为森林土壤提供了大量的碳和养分输入[29]。本研究发现, 4种林型在去除凋落物后增强了土壤对CH4的吸收, 研究结果与其他地区的相似[10-11, 30]。去除凋落物后, 土壤部分裸露, 会使得土壤温度升高、湿度降低[31], 可使得甲烷氧化菌活性增强, 从而提高土壤对CH4的吸收。不同地区的林地土壤在去除凋落物后对CH4吸收通量的增加幅度不同, 本地区4种林型在去除凋落物后土壤对CH4的吸收提高了2.12%—12.15%, 这一研究结果小于很多地区的结果[10-11, 30], 这可能与不同地区的水热条件以及凋落物数量和分解速率有关。研究表明, 温度升高会促进凋落物的分解[32], 而本地区年平均气温远低于其他研究地区, 因此造成本地区凋落物分解速率不及其他地区。此外, 地表存在凋落物的时候, 凋落物分解会提高土壤中的N素含量, 使得N/C增大, 从而抑制CH4的吸收[33]。据报道, 凋落物本身不产生或者消耗CH4[34-35], 但是凋落物层会通过控制气体扩散到土壤中来影响土壤对CH4的吸收[36], 因此, 凋落物层可能扮演阻碍CH4扩散到土壤中的屏障角色, 从而当地表保存凋落物后会降低对CH4的吸收。但在本研究中, 只是确定了土壤CH4的净通量(即吸收通量), 没有对扩散量和汇集量分别验证, 所以需要进一步的研究来验证这一假设。
林下植被是森林生态系统的重要组成部分, 在维持生物多样性和土壤养分循环方面发挥着重要作用[37]。而去除林下植被既可以防治火灾发生还能促进幼树的生长[38], 因此, 去除林下植被被认为是提高森林生产力和生物量碳储量的有效途径之一[39-40]。本研究表明, 去除林下植被会提高土壤对CH4的吸收, 但提高幅度不尽相同, 阔叶林提高了0.84%—20.55%, 针叶林提高了5.18%—9.16%。这可能是去除林下植被后会增强地表土壤的通气性, 从而使得土壤温度升高并且加快土壤水分蒸发, 促进了甲烷氧化菌的活性, 从而增加土壤CH4吸收[41], 这与很多研究结果一致[1214]。一些研究表明, 植被在生长过程中, 可能会释放少量的CH4[42], 所以当去除林下植被后, CH4通量在吸收和排放的收支平衡过程中会向着吸收增加方向发展。去除林下植被还会降低枯落物的输入, 减少了凋落物的数量, 可能使土壤的N/C降低[33], 从而提高土壤对CH4的吸收。此外造成不同地区林地去除林下植被后CH4通量差异的原因可能还与气候带、林型以及植被去除方式[14](是否使用除草剂)有关。
林下植被和凋落物都去除后, 土壤没有小灌木、草本以及凋落物层的遮护, 直接暴露于阳光下, 从而加强表层土壤水分蒸发以及温度升高。本研究野外观测发现, 4种林型在去除林下植被和凋落物后与自然状态相比CH4吸收通量呈现不同程度升高, 白桦林、山杨林、樟子松林和兴安落叶松林, CH4吸收通量分别提高了3.94%、36.26%、11.70%和13.65%。这种变化模式类似于单一去除的结果, 但4种林型增加幅度不尽相同。两者都去除和只去除凋落物相比, 白桦林、山杨林和兴安落叶松林土壤CH4吸收通量分别提高了0.95%、24.09%和11.29%, 但樟子松林下降0.40%, 说明这2种处理方式对山杨林土壤CH4通量有显著影响(P<0.05);同样, 两者都去除和只去除林下植被相比, 白桦林、山杨林、樟子松林和兴安落叶松林土壤CH4吸收通量分别提高了3.07%、13.04%、2.33%和8.06%, 且只有山杨林达显著水平(P<0.05)。这说明林下植被和凋落物都去除后, 土壤CH4吸收通量的增加并不等于去除凋落物或去除林下植被土壤CH4吸收通量增加之和, 而是低于二者之和, 这表明林下植被和凋落物对土壤CH4通量的影响存在耦合效应。去除林下植被和凋落物相比单一处理, 土壤温度升高更多, 湿度降低也多(见表 3), 这会使得甲烷氧化菌更加活跃, 从而对CH4的吸收量大于单一处理的量。其次, 林下植被和凋落物去除可能会降低土壤N/C[33]和失去气体扩散的屏障[36], 从而使得CH4吸收通量增加, 但CH4吸收量增加不是累加关系, 这可能还与土壤本身的养分含量有关[41]。
土壤温度和湿度是影响CH4通量的2个重要因子, 但不同地区的水热因子对CH4通量影响不尽相同。本研究发现, 4种林型土壤CH4通量普遍与土壤温度呈现负相关关系[10, 19]。然而, 4种林型土壤CH4通量与土壤湿度之间不存在显著相关关系, 这可能是土壤湿度对CH4氧化速率的促进或抑制存在交互作用的影响, 而降水则是造成这种影响的关键因素[43]。一般认为, 土壤湿度对土壤CH4氧化速率的影响呈现抛物线变化, 即土壤湿度很低或很高的条件下, CH4吸收量减少[44]。土壤理化性质对CH4通量也有很大影响, 但不同林型和不同处理有很大差异, 土壤有机碳含量对山杨林和兴安落叶松林部分处理土壤CH4通量呈正相关, 因此土壤有机碳含量是影响CH4通量的主要因素[19]。全氮也是影响CH4吸收的重要因子之一[45], 但本研究发现, 除了白桦林, 其他三种林型的部分处理与全氮呈现一定的相关性, 这可能是由于本地区N素不是影响白桦林土壤CH4通量的关键性因子。白桦林和樟子松林不同处理下土壤CH4通量与铵态氮、硝态氮相关性不显著, 这与广州鹤山[13]的研究结果相一致。pH只对白桦林和兴安落叶松林影响显著, 但对山杨林和樟子松林影响不显著, 这可能是pH对土壤其他养分造成的影响导致的, 这有待进一步探索。
尽管CH4在大气中浓度含量远不及CO2, 但是其产生的增温效应更大[46], 而我国的大兴安岭寒温带林区又是重要的碳汇基地[47], 这对今后的碳汇经营过程中有着重要意义。本研究发现, 不同处理的4种林型土壤均表现为CH4的汇, 且相比于自然状态均表现吸收增加的趋势, 其中白桦林在不同处理后土壤对CH4的吸收增加不显著, 山杨林在去除林下植被和去除两者后显著提高了土壤对CH4的吸收, 樟子松林在去除凋落物后土壤CH4吸收增加最多, 兴安落叶松林在去除两者后土壤对CH4吸收增加显著, 但在这个过程中, 凋落物去除与否对4种林型土壤CH4通量影响相对较小。因此, 从提高林地土壤碳汇功能的角度来看, 在今后本地区森林经营过程中, 可以适当对山杨林和兴安落叶林进行林床清理。值得一提的是, 本结论是林下不同处理后1年内的研究结果, 可能存在短期效应, 而林下植被和凋落物对该地区土壤CH4通量影响的长期效应还需要进一步探究。
4 结论林下植被和凋落物处理对寒温带天然林生长季土壤CH4通量的影响因林型而异, 凋落物去除与否对土壤CH4通量影响没有达到显著水平, 林下植被去除仅对山杨林土壤CH4通量影响显著, 而同时去除林下植被和凋落物时对山杨林和兴安落叶松林土壤CH4通量影响显著。各处理条件下土壤CH4通量与土壤温度相关性密切, 而土壤湿度对土壤CH4通量的影响相对较小。今后还应对本地CH4通量增加取样频率做到长期且连续监测, 且进一步对土壤CH4通量组分进行研究。
| [1] |
Mosier A R, Delgado J A, Keller M. Methane and nitrous oxide fluxes in an acid Oxisol in western Puerto Rico: effects of tillage, liming and fertilization. Soil Biology and Biochemistry, 1998, 30(14): 2087-2098. DOI:10.1016/S0038-0717(98)00085-6 |
| [2] |
胡启武, 吴琴, 李东, 曹广民. 不同土壤水分含量下高寒草地CH4释放的比较研究. 生态学杂志, 2005, 24(2): 118-122. |
| [3] |
白贞智. 热带山地雨林土壤N2O, CH4通量及其对养分添加的响应[D]. 杨凌: 西北农林科技大学, 2014. http://cdmd.cnki.com.cn/article/cdmd-10712-1014430078.htm
|
| [4] |
Yu L J, Huang Y, Zhang W, Li T T, Sun W J. Methane uptake in global forest and grassland soils from 1981 to 2010. Science of the Total Environment, 2017, 607-608: 1163-1172. DOI:10.1016/j.scitotenv.2017.07.082 |
| [5] |
吴亚丛. 林下植被清除对樟树人工林有机碳库的影响[D]. 北京: 中国林业科学研究院, 2014. http://cdmd.cnki.com.cn/article/cdmd-82201-1014331889.htm
|
| [6] |
李媛良, 汪思龙, 颜绍馗. 杉木人工林剔除林下植被对凋落层养分循环的短期影响. 应用生态学报, 2011, 22(10): 2560-2566. |
| [7] |
贺同鑫, 李艳鹏, 张方月, 王清奎. 林下植被剔除对杉木林土壤呼吸和微生物群落结构的影响. 植物生态学报, 2015, 39(8): 797-806. |
| [8] |
Wang Q K, He T X, Wang S L, Liu L. Carbon input manipulation affects soil respiration and microbial community composition in a subtropical coniferous forest. Agricultural and Forest Meteorology, 2013, 178-179: 152-160. DOI:10.1016/j.agrformet.2013.04.021 |
| [9] |
Dou X L, Zhou W, Zhang Q F, Cheng X L. Greenhouse gas (CO2, CH4, N2O) emissions from soils following afforestation in central China. Atmospheric Environment, 2016, 126: 98-106. DOI:10.1016/j.atmosenv.2015.11.054 |
| [10] |
程功, 刘廷玺, 王冠丽, 段利民, 李东方. 降雨和凋落物对人工杨树林土壤温室气体通量的影响. 农业环境科学学报, 2019, 38(6): 1398-1407. |
| [11] |
Leitner S, Sae-Tun O, Kranzinger L, Zechmeister-Boltenstern S, Zimmermann M. Contribution of litter layer to soil greenhouse gas emissions in a temperate beech forest. Plant and Soil, 2016, 403(1/2): 455-469. |
| [12] |
李海防, 张杏锋. 剔除灌草和添加翅荚决明对厚荚相思林土壤温室气体排放的影响. 应用生态学报, 2010, 21(3): 563-568. |
| [13] |
李海防, 夏汉平, 傅声雷, 张杏锋. 剔除林下灌草和添加翅荚决明对尾叶桉林土壤温室气体排放的影响. 植物生态学报, 2009, 33(6): 1015-1022. DOI:10.3773/j.issn.1005-264x.2009.06.001 |
| [14] |
刘娟, 陈雪双, 吴家森, 姜培坤, 周国模, 李永夫. 剔除杂草对山核桃林地土壤温室气体排放的影响. 应用生态学报, 2015, 26(3): 666-674. |
| [15] |
段北星, 满秀玲, 宋浩, 刘家霖. 大兴安岭北部不同类型兴安落叶松林土壤呼吸及其组分特征. 北京林业大学学报, 2018, 40(2): 40-50. |
| [16] |
孙凤华, 袁健, 路爽. 东北地区近百年气候变化及突变检测. 气候与环境研究, 2006, 11(1): 101-108. |
| [17] |
Wu X W, Zang S Y, Ma D L, Ren J H, Chen Q, Dong X F. Emissions of CO2, CH4, and N2O fluxes from forest soil in permafrost region of Daxing'an Mountains, Northeast China. International Journal of Environmental Research and Public Health, 2019, 16(16): 2999-2999. DOI:10.3390/ijerph16162999 |
| [18] |
马秀枝, 张秋良, 李长生, 陈高娃, 王飞. 寒温带兴安落叶松林土壤温室气体通量的时间变异. 应用生态学报, 2012, 23(8): 2149-2156. |
| [19] |
梁东哲, 赵雨森, 曹杰, 辛颖. 不同恢复方式下大兴安岭重度火烧迹地林地土壤温室气体通量. 生态学报, 2019, 39(21): 7950-7959. |
| [20] |
王博, 段玉玺, 王伟峰, 李晓晶, 刘宗奇, 刘源. 植被恢复对库布齐沙漠非生长季土壤CH4、N2O通量的影响. 生态学报, 2020, 40(6): 2028-2038. |
| [21] |
梁东哲, 赵雨森, 辛颖. 大兴安岭重度火烧迹地天然次生林土壤温室气体通量及其影响因子. 应用生态学报, 2019, 30(3): 777-784. |
| [22] |
Tate K R. Soil methane oxidation and land-use change-from process to mitigation. Soil Biology and Biochemistry, 2015, 80: 260-272. DOI:10.1016/j.soilbio.2014.10.010 |
| [23] |
程淑兰, 方华军, 于贵瑞, 徐敏杰, 张裴雷, 郑娇娇, 高文龙, 王永生. 森林土壤甲烷吸收的主控因子及其对增氮的响应研究进展. 生态学报, 2012, 32(15): 4914-4923. |
| [24] |
吴祥文, 臧淑英, 马大龙, 任建华, 李昊, 赵光影. 大兴安岭多年冻土区森林土壤温室气体通量. 地理学报, 2020, 75(11): 2319-2331. DOI:10.11821/dlxb202011004 |
| [25] |
菊花, 申国珍, 徐文婷, 赵常明, 苏磊, 王杨, 谢宗强, 张秋良. 神农架主要森林土壤CH4、CO2和N2O排放对降水减少的响应. 生态学报, 2016, 36(20): 6397-6408. |
| [26] |
Amaral J A, Knowles R. Inhibition of methane consumption in forest soils by monoterpenes. Journal of Chemical Ecology, 1998, 24(4): 723-734. DOI:10.1023/A:1022398404448 |
| [27] |
李娜. 增温和施氮肥对荒漠草原生态系统土壤温室气体能量的影响[D]. 呼和浩特: 内蒙古农业大学, 2010. http://cdmd.cnki.com.cn/Article/CDMD-10129-2010150021.htm
|
| [28] |
Zhang Z H, Duan J C, Wang S P, Luo C Y, Chang X F, Zhu X X, Xu B B, Wang W Y. Effects of land use and management on ecosystem respiration in alpine meadow on the Tibetan plateau. Soil and Tillage Research, 2012, 124: 161-169. DOI:10.1016/j.still.2012.05.012 |
| [29] |
FAO Forestry Department. Global Forest Resources Assessment 2010: Main Report. Rome: Food and Agriculture Organization of the United Nations, 2010.
|
| [30] |
Fan J L, Luo R Y, McConkey B G, Ziadi N. Effects of nitrogen deposition and litter layer management on soil CO2, N2O, and CH4 emissions in a subtropical pine forestland. Scientific Reports, 2020, 10: 8959-8959. DOI:10.1038/s41598-020-65952-8 |
| [31] |
段北星, 蔡体久, 宋浩, 肖瑞晗. 寒温带兴安落叶松林凋落物层对土壤呼吸的影响. 生态学报, 2020, 40(4): 1357-1366. |
| [32] |
Singh K P, Singh P K, Tripathi S K. Litterfall, litter decomposition and nutrient release patterns in four native tree species raised on coal mine spoil at Singrauli, India. Biology and Fertility of Soils, 1999, 29(4): 371-378. DOI:10.1007/s003740050567 |
| [33] |
Boeckx P, Van Cleemput O. Methane oxidation in a neutral landfill cover soil: influence of moisture content, temperature, and nitrogen-turnover. Journal of Environmental Quality, 1996, 25(1): 178-183. |
| [34] |
Smith K A, Dobbie K E, Ball B C, Bakken L R, Sitaula B K, Hansen S, Brumme R, Borken W, Christensen S, Priemé A, Fowler D, Macdonald J A, Skiba U, Klemedtsson L, Kasimir-Klemedtsson A, Degórska A, Orlanski P. Oxidation of atmospheric methane in Northern European soils, comparison with other ecosystems, and uncertainties in the global terrestrial sink. Global Change Biology, 2000, 6(7): 791-803. DOI:10.1046/j.1365-2486.2000.00356.x |
| [35] |
Reith F, Drake H L, Küsel K. Anaerobic activities of bacteria and fungi in moderately acidic conifer and deciduous leaf litter. FEMS Microbiology Ecology, 2002, 41(1): 27-35. DOI:10.1111/j.1574-6941.2002.tb00963.x |
| [36] |
Peichl M, Arain M A, Ullah S, Moore T R. Carbon dioxide, methane, and nitrous oxide exchanges in an age-sequence of temperate pine forests. Global Change Biology, 2010, 16(8): 2198-2212. |
| [37] |
Wu J P, Liu Z F, Chen D M, Huang G M, Zhou L X, Fu S L. Understory plants can make substantial contributions to soil respiration: Evidence from two subtropical plantations. Soil Biology and Biochemistry, 2011, 43(11): 2355-2357. DOI:10.1016/j.soilbio.2011.07.011 |
| [38] |
Camprodon J, Brotons L. Effects of undergrowth clearing on the bird communities of the Northwestern Mediterranean Coppice Holm oak forests. Forest Ecology and Management, 2006, 221(1/3): 72-82. |
| [39] |
Peng Y Y, Thomas S C, Tian D L. Forest management and soil respiration: Implications for carbon sequestration. Environmental Reviews, 2008, 16(1): 93-111. |
| [40] |
Powers R F, Busse M D, Mcfarlane K J, Zhang J W, Young D H. Long-term effects of silviculture on soil carbon storage: does vegetation control make a difference?. Forestry, 2013, 86(1): 47-58. DOI:10.1093/forestry/cps067 |
| [41] |
Zhang J J, Li Y F, Chang S X, Qin H, Fu S L, Jiang P K. Understory management and fertilization affected soil greenhouse gas emissions and labile organic carbon pools in a Chinese chestnut plantation. Forest Ecology and Management, 2015, 337: 126-134. DOI:10.1016/j.foreco.2014.11.004 |
| [42] |
Carmichael M J, Bernhardt E S, Bräuer S L, Smith W K. The role of vegetation in methane flux to the atmosphere: should vegetation be included as a distinct category in the global methane budget?. Biogeochemistry, 2014, 119(1/3): 1-24. |
| [43] |
菊花, 申国珍, 马明哲, 葛结林, 徐文婷, 赵常明, 张秋良. 北亚热带地带性森林土壤温室气体通量对土地利用方式改变和降水减少的响应. 植物生态学报, 2016, 40(10): 1049-1063. DOI:10.17521/cjpe.2016.0069 |
| [44] |
Liu L, Estiarte M, Peñuelas J. Soil moisture as the key factor of atmospheric CH4 uptake in forest soils under environmental change. Geoderma, 2019, 355: 113920. DOI:10.1016/j.geoderma.2019.113920 |
| [45] |
张强, 沈燕, 韩天宇, 勾蒙蒙. 湖南莽山4种林型甲烷通量及其影响因子. 中南林业科技大学学报, 2017, 37(9): 104-111. |
| [46] |
Wu B, Mu C C. Effects on greenhouse gas (CH4, CO2, N2O) emissions of conversion from over-mature forest to secondary forest and korean pine plantation in northeast china. Forests, 2019, 10(9): 788-805. DOI:10.3390/f10090788 |
| [47] |
胡海清, 罗碧珍, 魏书精, 魏书威, 文正敏, 孙龙, 罗斯生, 王立明, 马洪斌. 大兴安岭5种典型林型森林生物碳储量. 生态学报, 2015, 35(17): 5745-5760. |