 2021, Vol. 41
2021, Vol. 41文章信息
- 吴洪潘, 刘冬志, 马伟, 戚英杰, 邵长亮, 初红军
- WU Hongpan, LIU Dongzhi, MA Wei, QI Yingjie, SHAO Changliang, CHU Hongjun
- 新疆卡拉麦里山有蹄类野生动物自然保护区啮齿动物群落多样性及其与环境因子的关系
- Community diversities of rodents and its relationship with environmental factors in the Mt. Kalamaili Ungulate Nature Reserve, Xinjiang
- 生态学报. 2021, 41(23): 9511-9524
- Acta Ecologica Sinica. 2021, 41(23): 9511-9524
- http://dx.doi.org/10.5846/stxb202011082856
-
文章历史
- 收稿日期: 2020-01-10
- 网络出版日期: 2021-07-24





2. 新疆卡拉麦里山有蹄类野生动物自然保护区管理中心, 乌鲁木齐 830000;
3. 新疆环疆绿源环保科技有限公司, 乌鲁木齐 830019;
4. 新疆林业科学研究院森林生态研究所, 乌鲁木齐 830002
2. Management Center, Mt. Kalamaili Ungulate Nature Reserve, Urumqi 830000, China;
3. Huanjiang Lvyuan Environmental Protection Technology Co., Ltd. Urumqi 830019, China;
4. Institute of Forest Ecology, Xinjiang Academy of Forestry Sciences, Urumqi 830002, China
生物群落(Biotic community)是指栖居在一定空间、地域或生境中各生物种群通过相互作用而有机结合的复合体, 群落研究的两大基本内容包括群落组成和结构[1]。群落组成是决定群落性质的最重要因素, 通过研究群落的物种组成、多样性、丰富度、优势度及群落中各物种之间的相互作用性质和共存模式, 进一步认识群落[2]。群落结构包括群落的种类组成、种的优势度、群落多样性和优势种形态格局等[3], 其核心是群落的多样性, 它包括群落的物种组成和各物种的相对丰度。作为各种自然生态系统的一个功能单位和组织结构, 动物群落始终是动物生态学研究的核心之一。因具有扩散性强等特点, 动物群落成为动物生态学研究的重要内容, 除部分研究对象为昆虫外, 绝大部分的研究对象和材料是啮齿动物群落[4]。啮齿动物群落生态学的研究发展已成为当今生态学研究的热点领域, 并且在动物群落生态学研究中占有相当重要的位置[4-5]。
啮齿动物的群落多样性变化常被用来作为环境变化的重要测量指标, 是较好的生物多样性指示类群[6-8]。啮齿类动物(包括兔形目和啮齿目)具有种类数量多、分布广、取样相对简单、对环境变化敏感、移动性小等特点, 除了少数种类外, 一般体型均较小、繁殖快、适应力强, 多为穴居性, 能适应多种生境, 是群落学的理想研究对象[9-10]。群落的物种多样性与生态、生物地理和进化的诸因素相互作用有关。为了解这些因素如何影响物种的多样性, 则需研究啮齿动物群落与生境的关系[11]。关于啮齿动物群落多样性与不同环境因子的关系, 国内外已有大量研究[12-16]。研究啮齿动物群落多样性及其与环境因子关系不仅在生态学理论上具有重要意义, 而且可为鼠害防治拟订合理的防治方案、提高防治效果提供科学依据。
新疆卡拉麦里山有蹄类野生动物自然保护区(以下简称卡山保护区)位于我国新疆准噶尔盆地东部, 拥有荒漠、半荒漠的独特生态景观。一些学者对该地区的啮齿动物进行了调查, 如马勇等[17-19]、王思博等[20]和邵明勤等[21]。该区域还有针对大沙鼠(Rhombomys opimus)和其他鼠种的研究报道, 但多偏向于鼠疫[22-23]、鼠害[24-25]和道路生态[26]的调查。由于一些原因导致该保护区啮齿动物调查结果存在差异性。造成差异的原因:一、卡山保护区面积广阔, 横跨两个地州和六个县市[27], 未开展啮齿动物系统调查; 二、以前的调查研究都以防疫或植被[28-30]、大型有蹄类野生动物及其天敌为主[31-37], 专门对啮齿类动物群落的研究鲜见报道。为明晰卡山保护区啮齿动物的种类组成及群落结构的特点, 并探究啮齿动物群落多样性与不同环境因子之间的关系, 2019、2020年春季和夏季, 作者对卡山保护区九种生境中啮齿动物及其环境因子进行了系统调查研究, 以期为卡山保护区生态恢复、鼠害预防监测及综合治理提供基础资料和科学依据。
1 研究方法 1.1 研究地概况卡山保护区成立于1982年, 东经88°30′—90°00′, 北纬44°36′—46°00′, 东西宽117.5 km, 南北长147.5 km, 总面积14856.48 km2(图 1)。景观主要是戈壁、沙漠、裸岩山地和荒漠草原。该地区为典型的温带大陆性气候, 冬季漫长寒冷, 夏季炎热短暂, 年平均气温为2℃, 7月平均为20.5℃, 1月平均为-24.3℃, ≥10℃年积温为2617.1℃, 无霜期为117 d。受西风环流影响, 降水主要集中在冬季和春季, 年平均降水量低至159.1 mm, 年平均蒸发量为2090 mm[27]。由于地理位置、气候和土壤等生态因素的制约, 卡山保护区荒漠植被稀少, 结构单调, 植被覆盖稀疏。主要以旱生、超旱生和盐生的灌木、半灌木、小灌木和小半灌木为主, 无乔木。主要建群植物以菊科、藜科和柽柳科的种数较多, 其次为蒺藜科(Zygophyllaceae)、禾本科(Gramineae)。常见灌木有梭梭(Halxylon ammodendron)、木蓼(Atraphaxis frutescens)、沙拐枣(Calligonum mongolicum)、驼绒藜(Ceratoides latens)等。常见草本有假木贼(Anabasis salsa)、角果藜(Ceratocarpus arenarius)、针茅(Stipa capillata)、芨芨草(Achnatherum splendens)、木地肤(Kochia prostrata)、小甘菊(Cancrinia discoidea)、阿魏(Ferula sinkiangensis)等。卡山保护区气候干早, 植被稀疏, 野生动物以适应干旱和半干旱的种类占优势, 大中型有蹄类动物和小型啮齿动物是该荒漠和半荒漠景观的代表性物种[27, 38]。卡山保护区分布有大沙鼠、子午沙鼠(Meriones meridianus)、柽柳沙鼠(Meriones tamariscinus)、三趾跳鼠(Dipus sagitta)、五趾跳鼠(Allactaga sibirica)等啮齿动物。

|
| 图 1 2019年和2020年卡山保护区鼠铗样方设置 Fig. 1 Mousetraps samples in Mt. Kalamaili Ungulate Nature Reserve (MKUNR), 2019 and 2020 |
卡山保护区生境群落类型复杂多样, 根据资料查询和实地考察, 结合啮齿动物分布特点, 将调查区域划分为九种生境类型, 分别是:
(1) 戈壁生境(生境Ⅰ):
卡拉麦里山南和北坡前山带区域, 主要为砾石戈壁, 灌木有刺旋花(Convolvulus tragacanthoides)、蛇麻黄(Ephedra distachya)、梭梭、假木贼等, 主要的草本植物有角果藜、沙葱(Allium mongolicum)、沙蒿(Artemisia desertorum)、针茅、小甘菊、滨藜(Atriplex patens)、霸王(Zygophyllum fabago)等。该生境样地28块, 布铗共89 d, 调查面积17.5 km2。
(2) 恢复区生境(生境Ⅱ):
卡山保护区内原有的矿业、石料开采厂等已经实施了复垦、修复、再植等不同阶段生态恢复和修复的区域。主要的植物有雾滨藜(Bassia dasyphylla)、盐爪爪(Kalidium foliatum)、盐生草(Halogeton glomeratus)、沙蓬(Agriophyllum squarrosum)等, 部分区域复植梭梭。该生境样地29块, 布铗共83 d, 调查面积18.125 km2。
(3) 碱滩生境(生境Ⅲ):
季节性流水及融雪汇聚到卡山自然低洼地域, 在水分蒸腾干枯后析出盐碱, 形成碱滩生境。碱滩地势平坦, 是卡山保护区内野生动物补充矿物质的场所。土壤主要为盐碱土、龟裂土, 普通植被难以生长, 只有少数耐盐碱的植被才能存活。主要植物有琵琶柴(Reaumuria songonica)、盐爪爪和假木贼等。该生境样地31块, 布铗共92 d, 调查面积19.375 km2。
(4) 居民点生境(生境Ⅳ):
卡山保护区内国家公益林管护站及管理站、野生动物救护饲草料库, 是人工生境, 灌木主要有榆叶梅(Amygdalus triloba)、胡杨(Populus euphratica)、榆(Ulmus pumila)、沙枣(Elaeagnus angustifolia)、驼绒藜等, 草本植物主要有顶羽菊(Acroptilon repens)、狗尾草(Setaria viridis)等。该生境样地30块, 布铗共113 d, 调查面积18.6875 km2。
(5) 平滩生境(生境Ⅴ):
卡拉麦里山以南准东国家级工业园区及火烧山油田附近区域, 海拔相对高度差约400—600 m左右。灌木主要有梭梭、白梭梭、木蓼等, 草本植物主要有蛇麻黄、针茅、盐生草、滨藜等。该生境样地33块, 布铗共105 d, 调查面积20.625 km2。
(6) 沙地生境(生境Ⅵ):
卡山保护区卡喀木斯特管护站以西, 乔木西拜管护站以北, 朱马汗路沿线的大片区域, 有植被覆盖, 尚未沙漠化区域。主要的灌木植物有梭梭、驼绒藜、木本猪毛菜、琵琶柴等, 主要的草本植物有针茅、独尾草(Eremurus chinensis)、大叶补血草(Limonium gmelinii)、骆驼蓬(Peganum harmala)、肉苁蓉(Cistanche deserticola)等。该生境样地28块, 布铗共79 d, 调查面积17.5 km2。
(7) 沙漠生境(生境Ⅶ):
卡山保护区沙十九管护站、沙北油田、彩八油井、沙漠料场、库尔图牧办以及由此向北至保护区边界等为沙漠化区域, 土壤以风沙土为主。该区域灌木主要有白梭梭、沙拐枣、蛇麻黄等, 主要的草本植物有沙蒿、羽毛三芒草(Aristida pennata)、准噶尔无叶豆(Eremosparton songoricum)等。该生境样地38块, 布铗共111 d, 调查面积23.5625 km2。
(8) 山区生境(生境Ⅷ):
卡拉麦里山是东西走向的低山脉, 山体以东西向条山和突起的山岭为主, 岩体以中生代形成的残蚀岩为主, 山沟南北向较多。灌木植物主要有梭梭、驼绒藜、琵琶柴、柽柳(Tamarix chinensis)等, 草本植物主要有针茅、假木贼、木地肤、糙隐子草(Cleistogenes squarrosa)、骆驼蓬等。该生境样地23块, 布铗共60 d, 调查面积14.375 km2。
(9) 湿地生境(生境Ⅸ):
卡山保护区属内陆干旱区, 蒸腾作用强烈, 无常年地表径流水, 水资源匮乏, 仅在保护区的中部、西部、北部的沟谷地有裂隙水溢出形成山泉, 当地牧业人工开挖的泉水井, 卡拉麦里山南部戈壁与沙漠交界有20世纪50年代打出的自流井等水源地, 形成湿地生境, 是保护区主要的隐域景观。灌木主要有柽柳、白刺、盐穗木等, 草本植物主要有芨芨草、芦苇(Phragmites communis)、乳苣(Mulgedium tataricum)、小獐茅(Aeluropuspungens littoralis)等。该生境样地44块, 布铗共123 d, 调查面积27.125 km2。
1.2.2 调查方法2019年和2020年的春季和夏季(4—7月), 采用铗捕法[39-40], 对卡山保护区9种不同生境啮齿动物进行调查, 有效铗日55269个, 调查总面积176.875 km2。选择标准铁质板铗, 每块样地布设100个, 以浸泡过清水的花生米为诱饵。布设鼠铗时选择有啮齿动物痕迹的线路。利用GPS进行定位, 记录样地信息。保证每块样地持续布铗时间3 d至5 d。隔日检查时更换诱饵, 并对捕获过啮齿类的铁铗消毒后重新布铗。检查时, 将捕获的啮齿动物放入塑封袋, 喷洒杀虫剂并密封; 编号并记录日期、时间、经纬度、海拔、种类、地点及环境因子等信息; 返回实验室将啮齿动物放入-20℃冰箱冷冻。鉴定参考《新疆脊椎动物简志》、《新疆啮齿动物志》、《新疆北部地区啮齿动物的分类和分布》[19-20, 41]资料或咨询专家。
1.2.3 环境因子以捕获啮齿类个体的点为中心, 设置5 m×5 m的样方, 测量并记录样方内环境因子, 主要包括:
海拔高度(Altitude, AL):通过GPS记录捕获地的海拔高度;
植被种类(Number of plant species, NPS):样方的植物种数;
灌木种数(Number of shrub species, NSS):样方的灌木种数;
灌木高度(Shrub height, SH):样方的平均灌木高度;
灌木盖度(Species density, SD):样方的平均灌木盖度;
草本种数(Number of herbs species, NHS):样方的草本种数;
草本高度(Herbal height, HH):样方的平均草本高度;
草本盖度(Herbal density, HD):样方的平均草本盖度;
坡度(Slope, SL):平坡(≤5°), 缓坡(6°—15°), 斜坡(16°—25°), 陡坡(26°—35°), 急坡(36°—45°), 险坡(≥46°);
距水源地距离(Distance to water hole, DWH):样方到最近水源地的垂直距离;
距道路距离(Distance to road, DR):样方到最近道路(国道、便道)的垂直距离;
距居民点距离(Distance to residential area, DRA):样方到居民点(管护站等)的垂直距离。
1.2.4 数据分析(1) 啮齿动物生态地理分布型
采用郑智民等[42]对啮齿动物的地域分布同生态适应相结合的分型方法, 将卡山保护区啮齿动物分布区划分为2个一级生态地理分布型, 4个二级生态地理分布型, 17个三级生态地理分布型。
(2) 捕获的啮齿动物物种组成
捕获量比例计算公式如下:

|
(1) |
式中, N为捕获总数, Ni为某一种啮齿类的捕获数量。
捕获率计算公式如下:
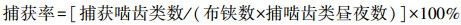
|
(2) |
(3) 啮齿动物群落多样性
群落多样性以群落多样性指数H、均匀性指数E、物种丰富度指数R和优势度指数D, 4个指标进行衡量。
群落多样性指数(Diversity index)以Shannon-Wiener公式计算, 计算公式:
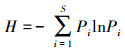
|
(3) |
式中, S为组成群落的啮齿类种数; Pi为第i种啮齿类个体在群落中所占的比例。
均匀性指数采用Pielou指数分析, 计算公式为:
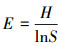
|
(4) |
式中, H为群落多样性指数; S为组成群落的啮齿类种数。
丰富度指数计算公式为:
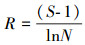
|
(5) |
式中, S为组成群落的啮齿类种数; N为所有啮齿类种个体总数。
优势度指数采用Simpson生态优势度指数分析。计算公式:

|
(6) |
式中, S为组成群落的啮齿类种数; N为所有啮齿类种个体总数; Ni为第i种啮齿类个体数。
(4) 啮齿动物群落相似性
用Jac-card群落相似性系数(q)分析啮齿动物群落的相似性。公式如下:

|
(7) |
式中, a和b分别为群落A和群落B包含的种类数量, c为群落A和群落B共有的种类数量, 当q∈(0, 0.25)时, 表示极不相似; 当q[0.25, 0.5)时, 表示中等不相似; 当q[0.5, 0.75)时, 表示中等相似; 当q[0.75, 1)时, 表示极相似。
采用Excel统计环境因子、啮齿动物物种组成、啮齿动物群落多样性指标、啮齿动物群落相似性系数等数据。使用SPSS19.0单因素方差分析(One-way ANOVA)对啮齿动物群落和环境因子在不同生境中的差异进行分析。采用SPSS19.0 Pearson相关性分析对环境因子与啮齿动物群落多样性指数进行相关性分析。对捕获量比例和不同生境类型啮齿动物群落相似性系数分别进行显著性检验, 对不同生境类型和样地内的啮齿动物环境因子进行多重比较(least significant difference, LSD)。
(5) 冗余分析
啮齿生物群落结构与环境因子的关系采用冗余分析(redundancy analysis, RDA)。将啮齿动物的捕获率进行hellinger转换成响应变量, 用九种生境中12个环境因子的标准化均值做解释变量。采用vegan软件(R语言)进行冗余分析(RDA)。开展响应变量去趋势对应分析(DCA), 若第一排序轴的长度小于3.0, 数据就适合进行RDA分析。利用RDA可明晰卡山保护区的啮齿动物群落与环境因子之间的关系, 分析影响啮齿动物群落结构变化的关键因子。
2 结果 2.1 卡山保护区啮齿动物物种组成与生态地理分布型在卡山保护区捕获啮齿动物419只, 分属1目4科10属12种, 包括赤颊黄鼠(Spermophilus erythrogenys)、褐家鼠(Rattus norvegicus)、大沙鼠、子午沙鼠、柽柳沙鼠、灰仓鼠(Cricetulus migratorius)、黄兔尾鼠(Eolagurus luteus)、五趾跳鼠、小五趾跳鼠(Allactaga elater)、羽尾跳鼠(Stylodipus telum)、三趾跳鼠、肥尾心颅跳鼠(Salpingotus crassicauda)(表 1)。12种啮齿动物包含2个一级地理分布型即耐旱型和人类伴生型。人类伴生型只有褐家鼠一种, 占捕获物种的1.91%。其余11种均为耐旱型, 占捕获物种的98.09%。说明耐旱型是卡山保护区的主要分布型, 17个三级地理分布型中占有4个(表 1)。
| 序号 No. |
科 Family |
属 Genus |
种类 Species |
生境Ⅰ Habitat Ⅰ |
生境Ⅱ Habitat Ⅱ |
生境Ⅲ Habitat Ⅲ |
生境Ⅳ Habitat Ⅳ |
生境Ⅴ Habitat Ⅴ |
生境Ⅵ Habitat Ⅵ |
生境Ⅶ Habitat Ⅶ |
生境Ⅷ Habitat Ⅷ |
生境Ⅸ Habitat Ⅸ |
地理分布型 Distribution pattrn |
总数 Total |
百分比/% Percentage |
丰富度 Abundance |
| 1 | 松鼠科 Sciuridae |
黄鼠属 Spermophilus |
赤颊黄鼠 Sp. erythrogenys |
9(14.29)* | 0 | 0 | 1(8.33) | 0 | 34(54.84) | 0 | 0 | 7(6.14) | (1)哈萨克-天山温旱型 | 51 | 12.17 | +++ |
| 2 | 鼠科 Muridae |
家鼠属 Rattus |
褐家鼠 Ra. norvegicus |
0 | 0 | 0 | 8(66.67) | 0 | 0 | 0 | 0 | 0 | (2)人类伴生型 | 8 | 1.91 | ++ |
| 3 | 仓鼠科 Cricetidae |
大沙鼠属 Rhombomys |
大沙鼠 Rh. opimus |
8(12.7) | 3(9.38) | 0 | 2(16.67) | 3(9.09) | 12(19.35) | 23(28.40) | 2(25.00) | 12(10.53) | (3)亚州西部温旱型 | 65 | 15.51 | +++ |
| 4 | 沙鼠属 Meriones |
子午沙鼠 M. meridianus |
3(4.76) | 0 | 2(14.29) | 0 | 2(6.06) | 0 | 0 | 1(12.5) | 3(2.63) | (3)亚州西部温旱型 | 11 | 2.63 | ++ | |
| 5 | 柽柳沙鼠 M. tamariscinus |
0 | 0 | 0 | 0 | 0 | 0 | 0 | 2(25.00) | 2(1.75) | (3)亚州西部温旱型 | 4 | 0.95 | + | ||
| 6 | 仓鼠属 Cricetulus |
灰仓鼠 C. migratorius |
1(1.59) | 4(12.5) | 0 | 1(8.33) | 0 | 1(1.61) | 0 | 0 | 0 | (3)亚州西部温旱型 | 7 | 1.67 | ++ | |
| 7 | 兔尾鼠属 Eolagurus |
黄兔尾鼠 E. luteus |
0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1(0.88) | (4)蒙新温旱型 | 1 | 0.24 | + | |
| 8 | 跳鼠科 Dipadidae |
五趾跳鼠属 Allactaga |
五趾跳鼠 A. sibirica |
33(52.38) | 19(59.38) | 3(21.43) | 0 | 9(27.27) | 10(16.13) | 1(1.23) | 0 | 41(35.96) | (1)哈萨克-天山温旱型 | 116 | 27.68 | ++++ |
| 9 | 小五趾跳鼠 A. elater |
0 | 1(3.13) | 0 | 0 | 0 | 0 | 0 | 0 | 9(7.89) | (3)亚州西部温旱型 | 10 | 2.39 | ++ | ||
| 10 | 羽尾跳鼠属 Stylodipus |
羽尾跳鼠 St. telum |
1(1.59) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1(0.88) | (1)哈萨克-天山温旱型 | 2 | 0.48 | + | |
| 11 | 三趾跳鼠属 Dipus |
三趾跳鼠 D. sagitta |
7(11.11) | 5(15.63) | 9(64.29) | 0 | 19(57.58) | 5(8.06) | 57(70.37) | 3(37.50) | 38(33.33) | (3)亚州西部温旱型 | 143 | 34.13 | ++++ | |
| 12 | 三趾心颅跳鼠属 Salpingotus |
肥尾心颅跳鼠 Sa. crassicauda |
1(1.59) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | (4)蒙新温旱型 | 1 | 0.24 | + | |
| 个体数(N)Individuals number | 63 | 32 | 14 | 12 | 33 | 62 | 81 | 8 | 114 | 419 | 100 | |||||
| 种数(S)Number of species | 8 | 5 | 3 | 4 | 4 | 5 | 3 | 4 | 9 | 12 | ||||||
| 耐旱型Drought tolerance | 8 | 5 | 3 | 3 | 4 | 5 | 3 | 4 | 9 | 11 | ||||||
| 伴生型Associated species | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | ||||||
| *括号数字表示物种占该生境的比例(%); ++++:优势种(P≥20%); +++:常见种(5%≤P<20%); ++:偶见种(1%≤P<5%); +:稀有种(P<1%).(1)哈萨克-天山温旱型; (2)人类伴生型; (3)亚州西部温旱型; (4)蒙新温旱型 | ||||||||||||||||
参考石锐等[8]的研究, 此次调查, 卡山保护区啮齿动物优势种是三趾跳鼠(34.13%)和五趾跳鼠(27.68%); 赤颊黄鼠(12.17%)和大沙鼠(15.51%)为常见种; 褐家鼠(1.91%)、子午沙鼠(2.63%)、灰仓鼠(1.67%)和小五趾跳鼠(2.39%)为偶见种; 柽柳沙鼠(0.95%)、黄兔尾鼠(0.24%)、羽尾跳鼠(0.48%)和肥尾心颅跳鼠(0.24%)为稀有种。其中, 肥尾心颅跳鼠是特别稀有的种类。在九种生境类型中优势种有一定的差异, 戈壁和恢复区生境中五趾跳鼠是优势种, 碱滩、平滩和湿地生境中三趾跳鼠和五趾跳鼠是优势种, 居民点生境中褐家鼠是优势种, 沙地生境中赤颊黄鼠是优势种, 沙漠生境中大沙鼠和三趾跳鼠是优势种, 山区生境中柽柳沙鼠、大沙鼠和三趾跳鼠是优势种(表 1)。
2.2 卡山保护区啮齿动物群落多样性在选取的生境类型中, 捕获的啮齿动物种类及个体数最多的是湿地生境(9种), 其次为戈壁生境(8种), 再则是恢复区生境(5种)、沙地生境(5种), 较少是居民点生境(4种)、平滩生境(4种)、山区生境(4种), 最少是碱滩生境(3种)和沙漠生境(3种)。丰富度指数(R)的变化规律为湿地生境>戈壁生境>恢复区生境=沙地生境>居民点生境=平滩生境=山区生境>碱滩生境=沙漠生境(表 2)。
| 生境 Habitat |
捕获数量/只 Catches |
捕获率 Capture-rate |
物种数 Species number |
丰富度指数R Richness index R |
多样性指数H Diversity index H |
均匀性指数E Evenness index E |
优势度指数D Dominance index D |
| 生境Ⅰ Habitat Ⅰ | 63 | 0.71% | 8 | 1.1593 | 1.4654 | 0.5897 | 0.9926 |
| 生境Ⅱ Habitat Ⅱ | 32 | 0.59% | 5 | 0.6625 | 1.1899 | 0.4789 | 0.9977 |
| 生境Ⅲ Habitat Ⅲ | 14 | 0.44% | 3 | 0.3312 | 0.8921 | 0.3590 | 0.9995 |
| 生境Ⅳ Habitat Ⅳ | 12 | 0.23% | 4 | 0.4969 | 0.9830 | 0.3956 | 0.9996 |
| 生境Ⅴ Habitat Ⅴ | 33 | 1.38% | 4 | 0.4969 | 1.0600 | 0.4266 | 0.9974 |
| 生境Ⅵ Habitat Ⅵ | 62 | 1.44% | 5 | 0.6625 | 1.2110 | 0.4873 | 0.9919 |
| 生境Ⅶ Habitat Ⅶ | 81 | 2.18% | 3 | 0.3312 | 0.6589 | 0.2652 | 0.9785 |
| 生境Ⅷ Habitat Ⅷ | 8 | 0.40% | 4 | 0.4969 | 1.3209 | 0.5316 | 0.9999 |
| 生境Ⅸ Habitat Ⅸ | 114 | 0.57% | 9 | 1.3250 | 1.5929 | 0.6410 | 0.9806 |
湿地生境和戈壁生境的多样性指数(H)较高, 且差异较小, 分别为1.5929和1.4654, 其次是山区生境1.3209, 沙地生境1.2110, 恢复区生境1.1899, 平滩生境1.0600, 居民点生境0.9830, 碱滩生境0.8921, 最小是沙漠生境0.6589。多样性指数(H)的变化规律是湿地生境>戈壁生境>山区生境>沙地生境>恢复区生境>平滩生境>居民点生境>碱滩生境>沙漠生境(表 2)。
均匀性指数(E):湿地生境(0.6410)>戈壁生境(0.5897)>山区生境(0.5316)>沙地生境(0.4873)>恢复区生境(0.4789)>平滩生境(0.4266)>居民点生境(0.3956)>碱滩生境(0.3590)>沙漠生境(0.2652)(表 2)。
优势度指数(D):山区生境(0.9999)>居民点生境(0.9996)>碱滩生境(0.9995)>恢复区生境(0.9977)>平滩生境(0.9974)>戈壁生境(0.9926)>沙地生境(0.9919)>湿地生境(0.9806)>沙漠生境(0.9785)(表 2)。
研究区域不同生境之间的群落相似系数结果显示, 平滩与碱滩生境、沙漠与平滩生境之间啮齿动物群落间的Jac-card相似系数最高(0.75), 达到极相似水平q∈[0.75, 1), 11个生境系数达到中等相似水平q∈[0.5, 0.75), 18个生境系数达到中等不相似水平q∈[0.25, 0.5), 5个生境系数达到极不相似水平q∈(0, 0.25)(表 3)。
| 生境类型 Habitat types |
生境Ⅰ Habitat Ⅰ |
生境Ⅱ Habitat Ⅱ |
生境Ⅲ Habitat Ⅲ |
生境Ⅳ Habitat Ⅳ |
生境Ⅴ Habitat Ⅴ |
生境Ⅵ Habitat Ⅵ |
生境Ⅶ Habitat Ⅶ |
生境Ⅷ Habitat Ⅷ |
生境Ⅸ Habitat Ⅸ |
| 生境Ⅰ Habitat Ⅰ | - | ||||||||
| 生境Ⅱ Habitat Ⅱ | 0.444** | - | |||||||
| 生境Ⅲ Habitat Ⅲ | 0.375 | 0.333 | - | ||||||
| 生境Ⅳ Habitat Ⅳ | 0.333 | 0.286 | 0.000 | - | |||||
| 生境Ⅴ Habitat Ⅴ | 0.500 | 0.500 | 0.750** | 0.143 | - | ||||
| 生境Ⅵ Habitat Ⅵ | 0.625 | 0.667 | 0.333 | 0.500 | 0.500 | - | |||
| 生境Ⅶ Habitat Ⅶ | 0.375 | 0.600 | 0.500** | 0.167 | 0.750** | 0.600 | - | ||
| 生境Ⅷ Habitat Ⅷ | 0.333 | 0.286 | 0.400* | 0.143 | 0.600* | 0.286 | 0.400** | - | |
| 生境Ⅸ Habitat Ⅸ | 0.545** | 0.400** | 0.333** | 0.182 | 0.444** | 0.400 | 0.333* | 0.444 | - |
| 差异不显著(P≥0.05), *差异显著(0.01<P<0.05), **差异极显著(P≤0.01) | |||||||||
恢复区、戈壁和湿地生境海拔高度高, 湿地生境的植被种数、灌木种数、灌木盖度、距居民点距离的环境因子和其他生境差异明显, 居民点生境灌木高度高, 沙地生境草本种数多, 湿地和山区生境草本高度高, 湿地和沙地生境草本盖度大, 山区生境坡度大, 沙漠生境距水源距离远, 平滩生境距道路距离远。单因素方差分析显示, 12个环境因子在九种生境类型中均呈极显著差异(P<0.01), 说明卡山保护区啮齿动物分布的生境异质性高(表 4)。
| 生境类型 Habitat types |
海拔高度 AL/m |
植被种数 NPS/n |
灌木种数 NSS/n |
灌木高度 SH/m |
灌木盖度 SD/% |
草本种数 NHS/n |
草本高度 HH/m |
草本盖度 HD/% |
坡度 SL/(°) |
距水源距离 DWH/km |
距道路距离 DR/km |
距居民点距离 DRA/km |
| 生境Ⅰ Habitat Ⅰ | 991.39±32.99a | 5±0.23a | 2.18±0.18a | 0.90±0.12a | 15.61±1.68a | 2.85±0.17a | 0.31±0.64a | 18.24±2.28a | 8.36±2.62a | 8.06±1.41a | 0.30±0.09a | 11.92±2.12a |
| 生境Ⅱ Habitat Ⅱ | 923.10±42.51a | 4.43±0.40a | 2.00±0.25a | 1.28±0.40a | 9.86±2.24a | 2.48±0.25a | 0.23±0.03a | 12.00±3.04a | 13.62±4.65a | 11.57±2.28a | 0.67±0.18a | 16.97±3.26a |
| 生境Ⅲ Habitat Ⅲ | 467.91±2.48b | 3.91±0.34a | 2.36±0.24a | 1.56±0.23a | 15.64±2.04a | 1.55±0.16b | 0.17±0.04a | 12.09±2.35a | 0.00a | 4.72±0.53a | 0.39±0.13a | 5.60±0.67a |
| 生境Ⅳ Habitat Ⅳ | 640.17±61.55b | 4.67±0.26a | 2.00±0.20a | 3.90±0.80b | 15.56±3.82a | 2.61±0.20a | 0.63±0.10a | 15.44±2.54a | 6.22±3.49a | 1.95±0.45b | 0.19±0.06a | 1.22±0.93a |
| 生境Ⅴ Habitat Ⅴ | 583.14±83.74b | 3.57±0.53a | 2.00±0.31a | 1.43±0.19a | 15.43±4.53a | 1.71±0.29a | 0.36±0.23a | 14.57±7.05a | 0.14±0.14a | 9.29±3.56a | 0.87±0.70a | 4.29±1.17a |
| 生境Ⅵ Habitat Ⅵ | 853.50±61.22a | 4.75±0.31a | 1.69±0.18a | 1.14±0.21a | 17.50±1.81a | 3.13±0.29a | 0.40±0.08a | 31.63±4.42a | 10.50±3.97a | 10.16±2.15a | 0.45±0.12a | 12.91±3.69a |
| 生境Ⅶ Habitat Ⅶ | 622.36±65.05b | 3.55±0.39b | 1.36±0.15a | 1.30±0.34a | 8.50±1.97a | 2.18±0.33a | 0.21±0.04a | 25.65±6.31a | 10.27±2.63a | 18.86±3.81c | 0.19±0.08a | 10.06±2.62a |
| 生境Ⅷ Habitat Ⅷ | 850.00±24.08a | 4.86±0.40a | 2.14±0.40a | 1.59±0.31a | 14.14±4.83a | 2.71±0.42a | 0.75±0.17a | 15.29±7.20a | 38.29±13.46b | 3.01±1.63a | 0.04±0.02a | 9.50±4.76a |
| 生境Ⅸ Habitat Ⅸ | 911.65±21.74a | 5.29±0.17a | 2.47±0.11a | 1.80±0.20a | 22.20±1.65a | 2.80±0.15a | 0.85±0.09b | 30.00±2.27b | 16.43±2.40a | 1.01±0.36b | 0.58±0.17a | 19.27±1.77a |
| F | 2363.86 | 2120.25 | 1007.51 | 166.14 | 359.69 | 1027.07 | 151.29 | 310.68 | 80.98 | 92.82 | 38.52 | 185.55 |
| df | 290 | 290 | 290 | 290 | 290 | 290 | 290 | 290 | 290 | 290 | 290 | 290 |
| P | 0.000** | 0.000** | 0.000** | 0.000** | 0.000** | 0.000** | 0.000** | 0.000** | 0.000** | 0.000** | 0.000** | 0.000** |
| 差异不显著(P≥0.05), *差异显著(0.01<P<0.05), **差异极显著(P≤0.01); AL, 海拔高度Altitude; NPS, 植被种类Number of plant species; NSS, 灌木种数Number of shrub species; SH, 灌木高度Shrub height; SD, 灌木盖度Species density; NHS, 草本种数Number of herbs species; HH, 草本高度Herbal height; HD, 草本盖度Herbal density; SL, 坡度Slope; DWH, 距水源地距离Distance to water hole; DR, 距道路距离Distance to road; DRA, 距居民点距离Distance to residential area | ||||||||||||
Pearson相关性分析结果显示, 捕获啮齿动物的个体数量(N)与草本盖度、距居民点距离存在强正相关关系; 啮齿动物物种数量(S)与海拔高度、植被种数、灌木盖度、距居民点距离存在强正相关关系; 群落多样性指数(H)与海拔高度、植被种数、灌木种数、灌木盖度、草本种数存在强正相关关系; 均匀性指数(E)与海拔高度、植被种数、灌木种数、灌木盖度、草本种数存在强正相关关系; 丰富度指数(R)与海拔高度、植被种数、灌木盖度、距居民点距离存在强正相关关系; 优势度指数(D)与草本盖度存在强负相关关系(表 5)。
| 环境因子 Environmental factors |
个体数(N) Individuals number |
种类个数(S) Number of species |
多样性指数(H) Diversity index |
均匀性指数(E) Evenness index |
丰富度指数(R) Richness index |
优势度指数(D) Dominance index |
| AL | 0.396 | 0.759* | 0.813** | 0.813** | 0.759* | -0.179 |
| NPS | 0.251 | 0.760* | 0.858** | 0.858** | 0.760* | -0.026 |
| NSS | -0.094 | 0.497 | 0.627 | 0.627 | 0.497 | 0.314 |
| SH | -0.339 | -0.182 | -0.189 | -0.189 | -0.182 | 0.263 |
| SD | 0.322 | 0.620 | 0.653 | 0.653 | 0.620 | -0.070 |
| NHS | 0.358 | 0.583 | 0.626 | 0.626 | 0.583 | -0.214 |
| HH | 0.159 | 0.429 | 0.585 | 0.585 | 0.430 | -0.086 |
| HD | 0.821** | 0.416 | 0.225 | 0.225 | 0.416 | -0.783* |
| SL | -0.039 | 0.132 | 0.424 | 0.424 | 0.132 | -0.008 |
| DWH | 0.230 | -0.345 | -0.550 | -0.550 | -0.345 | -0.393 |
| DR | 0.207 | 0.218 | 0.161 | 0.161 | 0.218 | 0.004 |
| DRA | 0.677* | 0.668* | 0.599 | 0.599 | 0.668 | -0.525 |
| 差异不显著(P≥0.05), *差异显著(0.01<P<0.05), **差异极显著(P≤0.01) | ||||||
采用DCA分析, 第一排序轴的长度小于3.0, 确定数据适合做线性模型的RDA分析。RDA对12个环境因子变量进行筛选, 得到与啮齿动物群落结构呈显著关系的4个变量:海拔高度、灌木盖度、灌木高度、植被种数。横坐标表示RDA1排序轴, 解释41.95%的信息, 与海拔高度相关性最强, 灌木盖度、灌木高度次之, 均呈负相关, RDA1从左到右表示随着海拔高度、灌木盖度、灌木高度逐渐增强, 三趾跳鼠与这三个成分的相关性强, 呈正相关, 说明其偏好视野开阔、灌木低矮稀疏、海拔低的生境。与之相反是褐家鼠, 偏好选择灌木密集高大, 利于隐蔽, 容易获取食物的居民点生境。纵坐标表示RDA2排序轴, 解释28.35%的信息, 与海拔高度相关性最强, 灌木盖度次之, 都呈正相关, RDA2从上到下表示随着海拔高度降低, 灌木盖度较小, 柽柳沙鼠与这两个成分的相关性最强, 呈正相关, 说明其偏好海拔低、灌木稀疏的生境。与之相反是五趾跳鼠, 偏好选择海拔高, 灌木盖度较大的生境(图 2)。

|
| 图 2 新疆卡山保护区生境与啮齿动物环境因子RDA排序图 Fig. 2 RDA ranking of rodents and habitats in MKUNR, Xinjiang A, 戈壁Gobi Beach; B, 恢复区Recovery Area; C, 碱滩Alkaline Beach; D, 居民点Residential Area; E, 平滩Flat Beach; F, 沙地Sandy Land; G, 沙漠Desert; H, 山区Ravine; I, 湿地Wetland; SP, 赤颊黄鼠Spermophilus erythrogenys; RA, 褐家鼠Rattus norvegicus; RH, 大沙鼠Rhombomys opimus; MEM, 子午沙鼠Meriones meridianus; MET, 柽柳沙鼠Meriones tamariscinus; CR, 灰仓鼠Cricetulus migratorius; EO, 黄兔尾鼠Eolagurus luteus; AS, 五趾跳鼠Allactaga sibirica; AE, 小五趾跳鼠Allactaga elater; ST, 羽尾跳鼠Stylodipus telum; DS, 三趾跳鼠Dipus. sagitta; SA, 肥尾心颅跳鼠Salpingotus crassicauda |
啮齿动物是生态系统营养网中的关键生物, 其丰度分布影响着捕食者的种群动态平衡[43-44], 其数目动态变化可以在更高水平的食物链中持续性显著影响其他动物的数目[43, 45]。数量可观的啮齿动物, 可为犬科、猫科、鼬科、猛禽、蛇类等天敌动物提供重要的食物来源, 对荒漠生态系统的平衡和稳定具有重要意义。
新疆地区已发现的啮齿动物有75种[46], 卡山保护区有记录的啮齿动物23种(内部资料), 本次卡山保护区调查发现的啮齿动物(12种)仅占新疆地区总种数的16%, 占卡山保护区记录种数的52.17%。此次调查物种数减少, 可能是因为诱饵单一, 样地选择的代表性需要进一步改进。目前, 准噶尔盆地啮齿类调查较为详细的是张渝疆等[22]对该区域鼠疫动物的调查, 发现1目3科13种啮齿动物, 与此次1目4科10属12种啮齿动物的调查结果基本相近。此次调查首次在卡山保护区捕捉到褐家鼠, 总捕获率为1.26%, 增加了卡山保护区啮齿动物种类。褐家鼠随人类活动侵入到卡山保护区, 且仅在居民点生境发现, 说明褐家鼠与人类伴生, 尚未侵入卡山保护区内其他生境。以往调查到的伴人型小家鼠(Mus musculus)在此次调查中未见捕获[22]。推测小家鼠、褐家鼠生境重叠竞争激烈[47-48], 或可能调查强度不够, 密度极低。
3.2 卡山保护区啮齿动物生态地理分布型郑智明等[42]认为物种分布区的地理位置、范围大小是动物在起源、演化、人为干扰、生态条件等多种因素影响下长期适应的结果。按其分类标准, 卡山保护区的啮齿动物的三级地理分布型有4种, 二级和一级地理分布型各是2种, 其中一级地理分布型耐旱型是本研究区域的主要分布型, 占捕获鼠种的98.09%。除褐家鼠外, 其余均为耐旱型(表 1)。而耐旱型占绝对优势, 与卡山保护区的地理生态环境有关。欧亚大陆中心地带的准噶尔盆地的啮齿动物地理群具有西部哈萨克荒漠和东部蒙古荒漠的过渡性质, 被认为是东部与西部起源的荒漠啮齿动物交汇区[10, 19, 20]。该地区干旱少雨, 但水源地、湿地等隐域生境植被茂密, 相对优越的自然环境使其成为荒漠野生动物栖息地的隐域景观。水源地、湿地等隐域生境, 为啮齿动物提供了食物来源和隐蔽条件。多种因素综合作用, 使得耐旱型啮齿动物在卡山保护区占据绝对的优势。
3.3 卡山保护区啮齿动物群落多样性卡山保护区啮齿动物分布的九种生境中的优势种有一定的差异, 不同鼠种对栖息地适应能力存在差异, 导致不同生境、同一生境的不同环境因子啮齿动物群落会产生差异。2020年春夏季卡山保护区遭遇多年罕见的干旱, 2019年春夏季的常见种赤颊黄鼠在2020年没有捕获。
多样性指数能反映群落内物种数以及不同物种数量的结构水平, 一般而言, 丰富度指数高的生境, 其物种多样性也较高[8]。湿地生境(1.3250)的丰富度指数高, 多样性指数也较高(1.5929)。湿地生境周边植被状况良好、邻近水源、离道路距离远、人为影响少、并处于多种生境交汇的区域, 符合“边缘效应”[1], 因此该区域捕获的物种数和数量都是最高的。但有例外, 居民点、平滩、山区生境的丰富度指数均是0.497, 多样性指数则为0.983、1.060、1.321。Hafner[49]认为啮齿类群落的多样性与其群落的复杂程度有一定关系。居民点属于人工生境, 人为干预影响的影响较大。除灌木种数、灌木高度和距道路距离基本接近外, 湿地生境的群落空间异质性远远高于植被结构单一的平滩生境, 这在捕获数量、物种数、丰富度指数、多样性指数、均匀性指数方面差异明显。山区捕获的啮齿动物数量相对湿地生境要少, 除优势度指数略比湿地生境高外, 其他群落多样性特征值均不如湿地生境(表 2、4)。居民点、平滩、山区生境捕获啮齿动物的种类和数量较少, 这可能是造成丰富度指数和多样性指数差异的重要原因。
姚圣忠等[50]认为, 植被单一、物种结构简单、气候多变的生态系统, 啮齿动物种群容易骤增, 从而形成灾害。卡山保护区西部的沙漠生境(81只)符合以上两个条件, 啮齿动物捕获量在九种生境中仅次于湿地生境(114只), 捕获的啮齿动物有57只大沙鼠、23只三趾跳鼠和1只五趾跳鼠。准噶尔盆地大沙鼠鼠疫自然疫源地是我国近期认定的新类型鼠疫自然疫源地[22], 若遇持续的极端气候, 如低温、干旱、植被大面积枯死, 极可能形成严重的鼠害, 发生大面积鼠害的同时散播鼠疫。
3.4 卡山保护区恢复区生境典型观念认为生态恢复就是物种回归, 指在迁地保护的基础上, 通过人工繁殖把植物引入到其原分布的自然或半自然的生境中, 并建立具有足够的遗传资源来适应进化改变、可自然维持和更新的新种群[51-52]。生态恢复是一个漫长的过程, 是帮助生态系统实现恢复区的演替, 要考虑到能够满足物种及其功能恢复的需要, 而不单单只是种类恢复。在生态系统中, 啮齿动物在地上和地下活动的同时带动了物质的循环、能量流动和信息传递过程中起着重要的地位和作用[4, 49, 53]。作为荒漠生态系统的初级消费者, 啮齿动物为次级消费者提供了重要的食物来源。通过对恢复区啮齿动物群落及其环境因子的研究, 可以了解卡山保护区生态恢复的效果, 为其他荒漠生境恢复提供借鉴。
2017年初开始, 卡山保护区开展了工矿企业整改工作, 并在矿区原址实施了生态恢复和修复, 包括矿区生态恢复和修复工程、湿地修复治理工程等多项工作。以卡山保护区戈壁生境等原始生境作为参考, 逐步使恢复区生境的各项指标达到或接近戈壁生境的程度。此次调查, 戈壁生境和恢复区生境啮齿类调查的有效铗日接近, 戈壁生境(63只)捕获啮齿动物数量比恢复区生境(32只)高近1倍, 除恢复区生境优势度指数略高于戈壁生境外, 其他指标戈壁生境均高于恢复区生境(表 2)。
在群落相似性系数方面, 恢复区与戈壁生境的相似性系数为0.444, 达到中等不相似水平; 比较两者间的环境因子, 灌木盖度和草本盖度, 戈壁比恢复区生境高; 灌木高度和距水源距离, 恢复区比戈壁生境高; 其他环境因子较接近。恢复区与沙地生境的相似性系数为0.667, 达到中等相似水平; 比较两者间的环境因子, 灌木盖度和草本盖度, 沙地比恢复区生境高; 恢复区生境的坡度比沙地高, 其他环境因子较接近。工矿企业将原戈壁生境的地表植被和土壤结构破坏, 人工恢复后的生境植被结构较单一, 以短命植物和草本为主, 灌木植物恢复相对较慢, 土壤结构类似于沙地生境, 所以两者的相似性系数较高, 达到中等相似水平(表 3)。
虽然已有植被再植, 啮齿动物定居, 生境逐渐恢复, 但需要注意的是目前矿区恢复区生态修复的工作远没有结束, 卡山保护区内尚有部分恢复区未进行植被再植修复, 地表还是裸岩和沙土。在今后的工作中, 还需进行长期细致的研究并配合持续性监测, 预防外来物种入侵和发生病原病害。
3.5 卡山保护区啮齿动物群落与环境因子关系啮齿动物群落与其生存环境变量之间的关系一直是生态学家、动物学家关注的问题。研究认为影响啮齿动物群落与环境因子之间关系的因素很多。王利清等[54]认为随着人类干扰活动的加剧, 小型啮齿动物的生物多样性有增高的趋势, 不同干扰方式对荒漠啮齿动物群落生物量会产生明显不同的作用。武晓东等[55]认为荒漠啮齿动物群落变量与植物群落变量中草本的关系最为密切, 可能与啮齿动物主要以草本植物为食物有关[56], 体现了食物资源利用共存原则。石锐等[8]研究认为植被盖度是决定内蒙古贺兰山啮齿动物群落结构最关键的环境因子。在干旱地区, 暖季水分对野生动物分布的影响巨大[57], 但大多数荒漠啮齿动物以植物为食并能从中获得水分, 重吸收水分能力强[58], 所以水源对荒漠啮齿动物群落的影响不似大型荒漠野生动物那么明显。
海拔高度、灌木盖度、灌木高度、植被种类4个环境因子是决定啮齿动物群落结构最主要的环境因子, 其中植被种数与群落多样性呈正相关, 随着植被种数数值的增加, 除优势度指数外, 其他多样性指数随之增加。RDA排序图反映了随着环境因子的变化, 影响啮齿动物群落结构变化的关键因子与生境选择的关系。5种啮齿动物都分布在植被种数高的湿地生境, 趋向性明显。因此, 卡山保护区荒漠啮齿动物群落多样性与环境因子密切相关, 植被种数是决定卡山保护区啮齿动物群落结构的关键环境因子。
致谢: 新疆环疆绿源环保科技有限公司和卡山自然保护区管理中心对研究提供帮助,新疆疾病预防与控制中心蒋卫研究员帮助鉴定鼠种, 宁夏大学周靖航博士、北京林业大学唐丽萍博士、新疆大学布威海丽且姆博士、张钧泳博士在软件计算和绘图方面提供帮助, 特此致谢。| [1] |
孙儒泳. 动物生态学原理 (第三版). 北京: 北京师范大学出版社, 2001.
|
| [2] |
姚圣忠. 张家口林地啮齿动物群落多样性及为害研究[D]. 北京: 北京林业大学, 2005.
|
| [3] |
Tilman D. A consumer-resource approach to community structure. American Zoologist, 1986, 26(1): 5-22. DOI:10.1093/icb/26.1.5 |
| [4] |
张晓东. 不同干扰下荒漠啮齿动物群落与植物群落、土壤相关性分析[D]. 呼和浩特: 内蒙古农业大学, 2010.
|
| [5] |
张渝疆, 孙素荣, 张大铭. 啮齿动物群落生态学研究现状. 新疆大学学报: 自然科学版, 2003, 20(4): 426-431. DOI:10.3969/j.issn.1000-2839.2003.04.021 |
| [6] |
Utrera A, Duno G, Ellis B A, Salas R A, de Manzione N, Fulhorst C F, Tesh R B, Mills J N. Small mammals in agricultural areas of the western Llanos of Venezuela: community structure, habitat associations, and relative densities. Journal of Mammalogy, 2000, 81(2): 536-548. DOI:10.1644/1545-1542(2000)081<0536:SMIAAO>2.0.CO;2 |
| [7] |
杨扬, 朱星, 赵豫桥. 北京小龙门地区啮齿类群落结构及变化调查. 安徽农业科学, 2008, 36(4): 1597-1598. DOI:10.3969/j.issn.0517-6611.2008.04.136 |
| [8] |
石锐, 李宗智, 高惠, 庞博, 燕永彬, 滕丽微, 刘振生. 内蒙古贺兰山啮齿动物群落多样性及其与环境因子关系. 兽类学报, 2019, 39(6): 651-661. |
| [9] |
Brown J H. Geographical ecology of desert rodents//Cody M L, Diamond J M, eds. Ecology and Evolution of Communities. Cambridge: Harvard University Press, 1975: 315-341.
|
| [10] |
胡德夫, 盛和林. 准噶尔盆地沙质荒漠啮齿动物群落在短命植物存在期的空间-食物资源利用. 兽类学报, 1999, 19(1): 25-36. DOI:10.3969/j.issn.1000-1050.1999.01.004 |
| [11] |
刘迺发, 范华伟, 敬凯, 宁瑞栋. 甘肃安西荒漠鼠类群落多样性研究. 兽类学报, 1990, 10(3): 215-220. |
| [12] |
Peay K G, Dickie I A, Wardle D A, Bellingham P J, Fukami T. Rat invasion of islands alters fungal community structure, but not wood decomposition rates. Oikos, 2013, 122(2): 258-264. DOI:10.1111/j.1600-0706.2012.20813.x |
| [13] |
Lohr C, van Dongen R, Huntley B, Gibson L, Morris K. Remotely monitoring change in vegetation cover on the Montebello Islands, western Australia, in response to introduced rodent eradication. PLoS One, 2014, 9(12): e114095. DOI:10.1371/journal.pone.0114095 |
| [14] |
乌云嘎, 查木哈, 张晓东, 袁帅, 付和平, 武晓东, 甘红军. 荒漠区啮齿动物群落与植物因子的冗余分析. 草业科学, 2014, 31(12): 2323-2332. DOI:10.11829/j.issn.1001-0629.2013-0699 |
| [15] |
蔡赫, 卞少伟. 天津古海岸与湿地保护区啮齿动物群落结构及其与环境因子关系. 兽类学报, 2015, 35(3): 288-296. |
| [16] |
叶丽娜, 袁帅, 付和平, 武晓东, 满都呼, 杨素文, 王剑然. 不同放牧方式对大针茅草原啮齿动物群落多样性的影响. 内蒙古大学学报: 自然科学版, 2017, 48(3): 312-319. |
| [17] |
马勇. 新疆北部地区啮齿动物地理分布的研究. 动物学报, 1981, 27(2): 180-188. |
| [18] |
马勇, 王逢桂, 金善科, 李思华, 林永烈, 叶宗耀. 新疆北部地区啮齿动物(GLIRES)的分类研究. 兽类学报, 1981, 1(2): 177-188. |
| [19] |
马勇, 王逢桂, 金善科, 李思华. 新疆北部地区啮齿动物的分类和分布. 北京: 科学出版社, 1987.
|
| [20] |
王思博, 杨赣源. 新疆啮齿动物志. 乌鲁木齐: 新疆人民出版社, 1983.
|
| [21] |
邵明勤, 阿布力米提·阿布都卡迪尔, 王德忠. 准噶尔荒漠鼠类群落结构及其栖息环境. 干旱区研究, 2001, 18(2): 65-68. |
| [22] |
张渝疆, 戴翔, 阿不力米提, 蒋卫, 阿不力克木, 王信惠, 布仁明德, 热娜, 李冰, 雷刚, 孟卫卫, 木合塔尔, 张晓兵, 王启国, 雒涛, 郭荣, 王忠, 唐建国, 刘成全, 阿扎提, 叶瑞玉, 于心, 曹汉礼. 准噶尔盆地鼠疫调查. 中华流行病学杂志, 2008, 29(2): 136-144. DOI:10.3321/j.issn:0254-6450.2008.02.008 |
| [23] |
叶超, 古丽阿依·包开西, 柳园园, 阿布力克木·阿布都热西提, 雒涛, 王启果, 孙开丽, 谢林芯, 张渝疆. 准噶尔盆地鼠疫自然疫源地东部区域大沙鼠体蚤群落结构分析. 疾病预防控制通报, 2017, 32(3): 1-4. |
| [24] |
杨永刚, 陈梦, 刘忠军, 廖力夫, 张娟. 三项调查指标与大沙鼠对林木危害率的相关性分析. 中国森林病虫, 2014, 33(1): 11-13. DOI:10.3969/j.issn.1671-0886.2014.01.004 |
| [25] |
盛兆湖, 陈梦, 刘忠军, 杨永刚, 王登元. 新疆昌吉州荒漠林区鼠类群落结构与梭梭被害关系的研究. 中国森林病虫, 2014, 33(2): 4-7. DOI:10.3969/j.issn.1671-0886.2014.02.002 |
| [26] |
Ji S N, Jiang Z G, Li L L, Li C W, Zhang Y J, Ren S E, Ping X G, Cui S P, Chu H J. Impact of different road types on small mammals in Mt. Kalamaili Nature Reserve. Transportation Research Part D: Transport and Environment, 2017, 50: 223-233. DOI:10.1016/j.trd.2016.11.006 |
| [27] |
初红军. 卡拉麦里山有蹄类自然保护区蒙古野驴(Equus hemionus)和鹅喉羚(Gazella subgutturosa)食性、种群和栖息地研究[D]. 北京: 中国科学院研究生院, 2008.
|
| [28] |
刘玉燕, 刘浩峰, 刘敏. 新疆卡拉麦里山有蹄类自然保护区生物多样性保护研究. 干旱环境监测, 2005, 19(3): 131-135. DOI:10.3969/j.issn.1007-1504.2005.03.002 |
| [29] |
唐海萍, 颜莉娟, 张新时. 新疆准噶尔盆地生物多样性保育与建立国家荒漠公园的构想. 生物多样性, 2008, 16(6): 618-626. DOI:10.3321/j.issn:1005-0094.2008.06.014 |
| [30] |
徐文轩, 杨维康, 张弛, 汪沐阳. 准噶尔盆地东部卡拉麦里山有蹄类自然保护区主要植物群落及其特征. 植物生态学报, 2016, 40(5): 502-507. |
| [31] |
葛炎, 刘楚光, 初红军, 陶永善. 新疆卡拉麦里山自然保护区蒙古野驴的资源现状. 干旱区研究, 2003, 20(1): 32-34, 66-66. |
| [32] |
吴洪潘, 初红军, 王渊, 马建伟, 葛炎, 布兰. 卡拉麦里山有蹄类自然保护区水源地蒙古野驴的活动节律: 基于红外相机监测数据. 生物多样性, 2014, 22(6): 752-757. |
| [33] |
董潭成, 初红军, 陈勇, 吴洪潘, 贺雷, 葛炎. 新疆卡拉麦里山有蹄类自然保护区鹅喉羚遗传多样性及系统发育地位. 兽类学报, 2016, 36(1): 77-86. |
| [34] |
吴兵, 初雯雯, 吴洪潘, 任松柏, 贺雷, 葛炎, 布兰, 初红军. 卡拉麦里山有蹄类自然保护区水源地野放普氏野马的活动节律: 基于红外相机监测数据. 动物学杂志, 2017, 52(4): 545-554. |
| [35] |
冯锦, 初雯雯, 端肖楠, 胡德夫, 吴洪潘, 葛炎, 初红军. 新疆卡拉麦里山有蹄类自然保护区蒙古野驴mtDNA D-loop区的遗传多样性及系统发育研究. 野生动物学报, 2018, 39(4): 737-744. DOI:10.3969/j.issn.1000-0127.2018.04.002 |
| [36] |
王渊, 初红军, 韩丽丽, 葛炎, 陶永善, 布兰. 基于红外相机陷阱技术的卡拉麦里山有蹄类自然保护区狼(Canis lupus)的活动节律. 干旱区研究, 2014, 31(4): 771-778. |
| [37] |
端肖楠, 初雯雯, 王渊, 杜聪聪, 何雷, 初红军. 新疆卡拉麦里山有蹄类自然保护区冬季狼的家域. 兽类学报, 2016, 36(4): 452-458. |
| [38] |
Xia C J, Cao J, Zhang H F, Gao X Y, Yang W K, Blank D. Reintroduction of Przewalski's horse (Equus ferus przewalskii) in Xinjiang, China: the status and experience. Biological Conservation, 2014, 177: 142-147. DOI:10.1016/j.biocon.2014.06.021 |
| [39] |
王思博, 孙玉珍, 赵新春. 准噶尔荒漠沙鼠Rhombomys opimus Licht. 种群密度调查方法. 地方病通报, 1998, 13(3): 81-83. |
| [40] |
李伟元. 夜夹法调查鼠情应注意的技术问题. 青海农技推广, 2007(2): 24-24. DOI:10.3969/j.issn.1008-7117.2007.02.042 |
| [41] |
袁国映. 新疆脊椎动物简志. 乌鲁木齐: 新疆人民出版社, 1991.
|
| [42] |
郑智民, 姜志宽, 陈安国. 啮齿动物学 (第二版). 上海: 上海交通大学出版社, 2012.
|
| [43] |
Mortelliti A, Boitani L. Interaction of food resources and landscape structure in determining the probability of patch use by carnivores in fragmented landscapes. Landscape Ecology, 2008, 23(3): 285-298. DOI:10.1007/s10980-007-9182-7 |
| [44] |
Ruiz-Capillas C, Mata C, Malo J E. Road verges are refuges for small mammal populations in extensively managed Mediterranean landscapes. Biological Conservation, 2013, 158: 223-229. DOI:10.1016/j.biocon.2012.09.025 |
| [45] |
Hanski I, Henttonen H, Korpimäki E, Oksanen L, Turchin P. Small-rodent dynamics and predation. Ecology, 2001, 82(6): 1505-1520. DOI:10.1890/0012-9658(2001)082[1505:SRDAP]2.0.CO;2 |
| [46] |
廖力夫. 新疆野生啮齿动物资源及其开发利用前景. 实验动物科学与管理, 2003, 20(z1): 18-20. DOI:10.3969/j.issn.1006-6179.2003.z1.006 |
| [47] |
于心, 张金桐, 叶瑞玉, 包兰宝, 吴克勤. 新疆铁路沿线和列车上鼠类的种属组成及其变迁——褐家鼠(Rattus norvegicus)在新疆的发现. 动物学研究, 1980, 1(1): 135-138. |
| [48] |
郭天宇. 家鼠控制中的若干问题. 中华卫生杀虫药械, 2002, 8(2): 6-8. |
| [49] |
Hafner M S. Density and diversity in Mojave desert rodent and shrub communities. Journal of Animal Ecology, 1977, 46(3): 925-938. DOI:10.2307/3650 |
| [50] |
姚圣忠, 胡德夫, 周娜, 陶炜煜, 宋淑霞, 刘恋. 我国森林啮齿动物的发生及防控措施研究现状. 中国森林病虫, 2005, 24(5): 22-26. DOI:10.3969/j.issn.1671-0886.2005.05.009 |
| [51] |
Maunder M. Plant reintroduction: an overview. Biodiversity & Conservation, 1992, 1(1): 51-61. |
| [52] |
Ren H, Zhang Q M, Lu H F, Liu H X, Guo Q F, Wang J, Jian S G, Bao H O. Wild plant species with extremely small populations require conservation and reintroduction in China. AMBIO, 2012, 41(8): 913-917. DOI:10.1007/s13280-012-0284-3 |
| [53] |
张美文, 王勇, 李波, 陈安国. "社会-经济-自然复合生态系统"中的鼠害治理. 兽类学报, 2003, 23(3): 250-258. DOI:10.3969/j.issn.1000-1050.2003.03.013 |
| [54] |
王利清, 武晓东, 付和平, 杨泽龙, 庄光辉, 甘红军. 不同干扰方式对荒漠植物与啮齿动物群落的作用. 内蒙古农业大学学报, 2006, 27(3): 46-52. |
| [55] |
武晓东, 阿娟, 付和平, 金珩. 人为不同干扰条件下荒漠啮齿动物群落格局的动态特征——动物与植物群落的典型相关分析. 生态学报, 2008, 28(12): 5999-6017. DOI:10.3321/j.issn:1000-0933.2008.12.028 |
| [56] |
Lima S L, Dill L M. Behavioral decisions made under the risk of predation: a review and prospectus. Canadian Journal of Zoology, 1990, 68(4): 619-640. DOI:10.1139/z90-092 |
| [57] |
周立志, 马勇. 中国西部干旱地区啮齿动物多样性分布格局. 生物多样性, 2002, 10(1): 44-48. DOI:10.3321/j.issn:1005-0094.2002.01.007 |
| [58] |
刘凌云, 郑光美. 普通动物学 (第四版). 北京: 高等教育出版社, 2009.
|