 2021, Vol. 41
2021, Vol. 41文章信息
- 付梦娣, 贾强, 任月恒, 周汉昌, 李俊生, 张渊媛, 白加德, 陈克林
- FU Mengdi, JIA Qiang, REN Yueheng, ZHOU Hanchang, LI Junsheng, ZHANG Yuanyuan, BAI Jiade, CHEN Kelin
- 环渤海滨海湿地鸻鹬类水鸟多样性及其环境影响因子
- Diversity and environmental influencing factors of curlew waterbirds in coastal wetlands of the Bohai Rim
- 生态学报. 2021, 41(22): 8882-8891
- Acta Ecologica Sinica. 2021, 41(22): 8882-8891
- http://dx.doi.org/10.5846/stxb202107051785
-
文章历史
- 收稿日期: 2021-07-05
- 修订日期: 2021-10-21





2. 中咨集团生态技术研究所(北京)有限公司, 北京 100048;
3. 中国科学院生态环境研究中心, 北京 100085;
4. 北京生物多样性保护研究中心, 北京 100076;
5. 北京麋鹿生态实验中心, 北京 100076;
6. 北京源河国际湿地文化交流中心, 北京 100029
2. China International Engineering Consulting Corporation, Ecological Technical Research Institute, Beijing 100048, China;
3. Research Center for Eco-Environment Sciences, Chinese Academy of Sciences, Beijing 100085, China;
4. Beijing Biodiversity Conservation Research Center, Beijing 100076, China;
5. Beijing Milu Ecological Research Center, Beijing 100076, China;
6. International Wetlands & River, Beijing 100029, China
随着经济社会的不断发展, 人类活动对环境影响逐步增大, 鸟类的物候学、行为模式、种群动态和分布范围也随之发生剧烈改变。鸟类对环境因子的响应机制是生态学研究中最基础和根本的问题之一[1]。20世纪的研究显示, 长距离迁徙鸟类的数量显著下降[2], 引起许多学者的关注, 发现气候因子[3-5]、食物量[6]、捕食作用[7-8]、栖息地质量[9-11]等是影响迁徙鸟类数量及分布的重要因子。此外, 不同时空下环境因子之间的相互作用, 增加了水鸟和环境因子之间关系及作用机制研究的不确定性[12]。研究水鸟群落与环境因子之间的关系, 对修复鸟类栖息地具有重要意义。
环渤海湿地是水鸟南北迁徙的重要驿站, 是东亚-澳大利西亚水鸟迁徙路线的重要组成部分, 尤其对于该线路上的鸻鹬鸟类具有非常重要的意义[13]。环渤海地区作为我国经济发展的热点区域, 滨海城市化及临海工业发展迅猛, 黄河、海河、辽河汇入渤海, 陆源污染物排海量大。近年来, 大规模的滩涂围垦、填海造地和高密度的养殖, 以及近岸海域环境污染, 致使滨海湿地大面积减少、功能退化严重[14], 水鸟栖息地不断丧失。环渤海滨海湿地成为整条水鸟迁徙路线上最受胁的节点之一, 水鸟保护形势严峻, 特别是鸻鹬类水鸟, 近年来种群数量下降趋势明显[15-16]。因此, 开展鸻鹬类水鸟调查, 明确鸻鹬类水鸟群落组成及其时空变化, 研究水鸟多样性与环境因子的响应关系十分必要。本研究采用多元回归分析和结构方程模型(Structural Equation Modeling, SEM) 对2016—2020年春季鸻鹬类水鸟多样性与气候、食物、保护强度等因子之间的关系进行分析, 以期为物种保护策略的制定提供必要的依据。
1 调查区概况将环渤海的12个重要湿地作为本研究的调查区(表 1、图 1), 总面积4431 km2, 涉及辽宁省大连、盘锦、锦西, 河北省秦皇岛、唐山、沧州, 天津, 以及山东省滨州和东营。渤海主要入海河流有辽河、滦河、永定河、海河、子牙河、黄河等。调查区涉及辽河、子牙河、黄河3处河口湿地。除辽东半岛和山东半岛属基岩海岸外, 绝大部分为平原海岸, 平原海岸湿地多为潮间带滩涂湿地和养殖池塘。广阔的滨海湿地孕育了极其丰富的生物区系, 包括腔肠动物、软体动物、藻类等在内的物种数量达到542种, 是各种水鸟的栖息、取食场所。调查区涉及了各级各类型自然保护地, 包括自然保护区、海洋公园、海洋特别保护区、森林公园、地质公园等, 总面积3197 km2, 覆盖了调查区的72%。
| 序号 Code |
调查地点 Survey site |
面积/ km2 Area |
滨海湿 地类型 Coastal wetland type |
涉及自然保护地名称 Name of the natural protected areas |
自然保护地面积占比/% Proportion of natural protected areas |
| P1 | 辽宁蛇岛老铁山湿地 | 90 | 滩涂湿地 | 辽宁蛇岛老铁山国家级自然保护区 | 64 |
| P2 | 辽宁营口-大连沿海湿地 | 226 | 养殖池塘 | 大连斑海豹国家级自然保护区 | 64 |
| P3 | 辽宁辽河口湿地 | 800 | 河口湿地 | 辽宁辽河口国家级自然保护区、辽宁辽河口红海滩国家海洋公园、辽宁盘锦辽河口省级地质公园 | 77 |
| P4 | 辽宁葫芦岛-锦州沿海湿地 | 210 | 养殖池塘 | 葫芦岛六股河入海口滨海湿地市级自然保护区 | 64 |
| P5 | 河北北戴河湿地 | 300 | 滩涂湿地 | 海滨国家森林公园、北戴河国家级海洋公园 | 91 |
| P6 | 河北乐亭沿海湿地 | 82 | 滩涂湿地 | 河北乐亭菩提岛省级自然保护区 | 77 |
| P7 | 河北曹妃甸湿地 | 183 | 养殖池塘 | 曹妃甸湿地和鸟类省级自然保护区 | 91 |
| P8 | 天津汉沽沿海湿地 | 360 | 滩涂湿地 | 天津古海岸与湿地国家级自然保护区 | 68 |
| P9 | 天津北大港沿海湿地 | 440 | 河口湿地 | 天津北大港湿地省级自然保护区 | 77 |
| P10 | 河北沧州沿海湿地 | 160 | 养殖池塘 | 河北南大港湿地和鸟类省级自然保护区 | 82 |
| P11 | 山东滨州贝壳堤岛湿地 | 440 | 养殖池塘 | 滨州贝壳堤岛与湿地国家级自然保护区 | 100 |
| P12 | 山东黄河三角洲湿地 | 1140 | 河口湿地 | 山东黄河三角洲国家级自然保护区、东营黄河口生态国家级海洋特别保护区、山东东营黄河三角洲国家地质公园、黄河口国家森林公园 | 91 |
| P: 位点Position | |||||

|
| 图 1 环渤海滨海湿地水鸟调查点位分布图 Fig. 1 The spatial distribution of waterbirds survey points in coastal wetlands of Bohai Rim P: 调查点位Position |
(1) 水鸟调查
环渤海湿地鸻鹬类水鸟调查于2016—2020年每年4月开展。依据《生物多样性观测技术导则鸟类》(HJ 710.4—2014), 采用样线法和样点法相结合的方法, 通过双筒望远镜(Bushnell legend 10×42)、高倍单筒望远镜(Leica televid 32×77)对调查区域内的水鸟进行观测, 记录物种、数量和位置。对于个体数量较少的鸟种直接计数, 对个体数量较大或集群繁殖鸟种以每10只、20只或50只为一个单位计数[17]。鸟类分类依据《中国鸟类分类与分布名录》(第三版)。鸟类保护等级依据《国家重点保护野生动物名录》(2021年)、《世界自然保护联盟濒危物种红色名录》(2018年)。
(2) 环境数据
气温、降水数据来源于调查区及其周边气象台站数据, 采用ANUSPLIN[18]对气象数据进行空间插值, 空间分辨率1 km, 时间序列2016—2020年。保护地空间分布数据来源于国家林业和草原局。浮游动植物密度、大型底栖生物密度数据来源于国家海洋科学数据中心的黄渤海滨海带环境污染与生态状况综合调查结果, 调查方法依据《海洋监测规范》(GB 17378—2007), 其中浮游动物利用Ⅰ型网采集样品;分析方法依据《近岸海洋生态健康评价指南》(HY/T 087—2005)。
2.2 研究方法(1) 水鸟多样性测度
鸟类多样性能够表征鸟类群落的组成结构和栖息环境的良好程度[19], 对维持生态系统的稳定性具有重要作用。参考蒋志刚[20]、邵明勤[21]等人的研究, 选取Shannon-Wiener多样性指数指征调查区鸻鹬类水鸟多样性, 计算公式为:
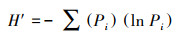
|
式中, H′为Shannon-Wiener多样性指数, Pi为i物种的个体在所有物种个体总数的比例。
(2) 结构方程模型建立
结构方程模型(Structural Equation Modeling, SEM)是基于变量的协方差矩阵分析变量之间关系的一种方法, 主要用于对复杂的多变量数据进行处理[22]。该模型由测量方程和结构方程两部分组成, 测量方程用来描述潜变量和显变量之间的关系, 结构方程是用来描述潜变量与潜变量之间的关系[23]。计算公式为:
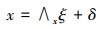
|
(1) |
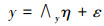
|
(2) |
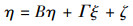
|
(3) |
式中, 方程(1)和方程(2)为测量方程, 方程(3)为结构方程。x是外生可测变量向量;x是外生潜变量向量;y是内生可测变量向量;η是内生潜变量向量;∧x为外生可测变量与外生潜变量之间的关系, 是外生可测变量在外生潜变量上的因子负荷矩阵;∧y为内生可测变量与内生潜变量之间的关系, 是内生可测变量在内生潜变量上的因子负荷矩阵;B是内生潜变量的关系阵;Γ则表示外生潜变量对于内生潜变量的影响;δ和ε为测量方程的误差项;ζ为结构方程的误差项。
水鸟对停歇点食物资源需求不同于繁殖地和越冬地, 食物资源的丰富度是影响水鸟对停歇地的选择的重要因素[24-25]。气候条件变化对湿地水鸟的影响包括气候变暖导致迁徙时间和迁徙距离发生变化, 以及改变停歇点湿地食物和栖息资源的提供等[26]。调查区已建立数量众多、类型丰富、功能多样的各级各类自然保护地, 在保护鸟类多样性、改善生态环境质量方面发挥了一定作用[27-28]。调查区为潮间带滩涂湿地、养殖池塘和河口湿地, 调查时间内水位、盐度、植被盖度基本稳定, 栖息地质量变化小。基于以上考虑和数据可获得性, 建立环渤海滨海湿地鸻鹬类水鸟多样性与环境因子响应关系的概念模型(图 2)。模型包括2个潜变量:气候和食物, 7个可测变量:年均温度、年均降雨、浮游植物密度、浮游动物密度、大型底栖生物密度、鸻鹬类水鸟多样性、保护强度, e1—e8是残差变量。其中保护强度指各调查区覆盖的自然保护地面积比例(表 1)。假设气候、食物、保护强度对鸻鹬类鸟类多样性有正向影响;气候、保护强度对食物有正向影响。运用Amos24.0进行结构方程模型的分析。采用绝对拟合指数、相对拟合指数和精简指数三类拟合指数, 包括x2/df、GFI、RMSEA、NFI、TLI、CFI、IFI、AIC和ECVI, 指征验证结果是否具有统计学意义。

|
| 图 2 环渤海滨海湿地鸻鹬类水鸟多样性与环境因子响应关系概念模型 Fig. 2 Conceptual model for the relationship between diversity and environmental factors of curlew waterbird in coastal wetlands of Bohai Rim |
(1) 群落组成
2016—2020年的水鸟调查共记录鸻鹬类7科51种(表 2), 其中旅鸟50种。全球极危物种1种, 濒危物种3种, 近危物种9种。国家一级保护鸟类2种, 国家二级保护鸟类8种。年最大种群个体数量较多的水鸟有黑腹滨鹬(125089只)、大滨鹬(112370只)、黑尾塍鹬(49306只)、灰鸻(47449只)、斑尾塍鹬(43267只)、白腰杓鹬(42487只)、环颈鸻(40172只)、红腹滨鹬(33167只)。经计算, 斑尾塍鹬(近危物种)、白腰杓鹬(近危物种)、大滨鹬(濒危物种)、环颈鸻的大部分迁徙种群在环渤海的滨海湿地中停歇[29]。
| 目/科/种 Order/Family/Species |
居留类型 Residence type |
年最大种群个体数量/只 Maximum number of individuals annually |
国家保护级别 National protection category |
IUCN红色名录等级 IUCN redlist |
| 鸻形目Charadriiformes | ||||
| (1)彩鹬科Rostratulidae | ||||
| 1.彩鹬Rostratula benghalensis | S/P | 1 | ||
| (2)蛎鹬科Haematopodidae | ||||
| 2.蛎鹬Haematopus ostralegus | P | 1281 | NT | |
| (3)鹮嘴鹬科Ibidorhynchidae | ||||
| 3.鹮嘴鹬Ibidorhyncha struthersii | R | 14590 | Ⅱ | |
| (4)反嘴鹬科Recurvirostridae | ||||
| 4.黑翅长脚鹬Himantopus himantopus | S/P | 16025 | ||
| 5.反嘴鹬Recurvirostra avosetta | S/P | 9116 | ||
| (5)燕鸻科Glareolidae | ||||
| 6.普通燕鸻Glareola maldivarum | S/P | 73 | ||
| (6)鸻科Charadriidae | ||||
| 7.凤头麦鸡Vanellus vanellus | P | 163 | NT | |
| 8.灰头麦鸡Vanellus cinereus | S/P | 368 | ||
| 9.金鸻Pluvialis fulva | P | 18406 | ||
| 10.灰鸻Pluvialis squatarola | P | 47449 | ||
| 11.长嘴剑鸻Charadrius placidus | S/P | 121 | ||
| 12.金眶鸻Charadrius dubius | S/P | 1739 | ||
| 13.环颈鸻Charadrius alexandrinus | S/P | 40172 | ||
| 14.蒙古沙鸻Charadrius mongolus | P | 1699 | ||
| 15.铁嘴沙鸻Charadrius leschenaultii | P | 506 | ||
| (7)鹬科Scolopacidae | ||||
| 16.黑尾塍鹬Limosa limosa | P | 49306 | NT | |
| 17.斑尾塍鹬Limosa lapponica | P | 43267 | NT | |
| 18.小杓鹬Numenius minutus | P | 1383 | Ⅱ | |
| 19.中杓鹬Numenius phaeopus | P | 13975 | ||
| 20.白腰杓鹬Numenius arquata | P | 42487 | Ⅱ | NT |
| 21.大杓鹬Numenius madagascariensis | P | 25973 | Ⅱ | EN |
| 22.小青脚鹬Tringa guttifer | P | 182 | Ⅰ | EN |
| 23.青脚鹬Tringa nebularia | P | 6779 | ||
| 24.红脚鹬Tringa totanus | P | 516 | ||
| 25.鹤鹬Tringa erythropus | P | 972 | ||
| 26.白腰草鹬Tringa ochropus | P/W | 2620 | ||
| 27.林鹬Tringa glareola | P | 933 | ||
| 28.翘嘴鹬Xenus cinereus | P | 1044 | ||
| 29.勺嘴鹬Calidris pygmeus | P | 1 | Ⅰ | CR |
| 30.泽鹬Tringa stagnatilis | P | 11398 | ||
| 31.矶鹬Actitis hypoleucos | P | 1123 | ||
| 32.翻石鹬Arenaria interpres | P | 212 | Ⅱ | |
| 33.丘鹬Scolopax rusticola | P | 5 | ||
| 34.扇尾沙锥Gallinago gallinago | P | 67 | ||
| 35.针尾沙锥Gallinago stenura | P | 8 | ||
| 36.孤沙锥Gallinago solitaria | P | 3 | ||
| 37.大沙锥Gallinago megala | P | 8 | ||
| 38.三趾滨鹬Calidris alba | P | 916 | ||
| 39.红腹滨鹬Calidris canutus | P | 33167 | NT | |
| 40.红颈滨鹬Calidris ruficollis | P | 14457 | NT | |
| 41.大滨鹬Calidris tenuirostris | P | 112370 | Ⅱ | EN |
| 42.小滨鹬Calidris minuta | P | 13 | ||
| 43.青脚滨鹬Calidris temminckii | P | 5215 | ||
| 44.长趾滨鹬Calidris subminuta | P | 20 | ||
| 45.尖尾滨鹬Calidris acuminata | P | 2443 | ||
| 46.黑腹滨鹬Calidris alpina | P | 125089 | ||
| 47.弯嘴滨鹬Calidris ferruginea | P | 6691 | NT | |
| 48.阔嘴鹬Calidris falcinellus | P | 620 | Ⅱ | |
| 49.流苏鹬Calidris pugnax | P | 43 | ||
| 50.半蹼鹬Limnodromus semipalmatus | P | 264 | Ⅱ | NT |
| 51.红颈瓣蹼鹬Phalaropus lobatus | P | 1856 | ||
| R: 留鸟,S: 夏候鸟,W: 冬候鸟,P: 旅鸟;Ⅰ: 国家Ⅰ级保护动物,Ⅱ: 国家Ⅱ级保护动物;CR:极危Critically Endangered,EN:濒危Endangered,NT:近危Near Threatened | ||||
(2) 群落时空变化
2016—2020年, 调查区鸻鹬类水鸟种类较多的湿地依次为山东黄河三角洲湿地、河北沧州沿海湿地(P10)、辽宁辽河口湿地(P3)、天津北大港沿海湿地(P9)、山东滨州贝壳堤岛湿地(P11)(图 3)。其中, 山东黄河三角洲湿地(P12)、辽宁辽河口湿地(P3)、天津北大港沿海湿地(P9)均为河口湿地, 占调查区总面积的53.71%, 单位面积鸻鹬类水鸟数量较少(图 4)。河北沧州沿海、山东滨州贝壳堤岛及其周边区域湿地集中连片, 为环渤海地区湿地集中区, 鸻鹬类水鸟种类较多。

|
| 图 3 环渤海滨海湿地鸻鹬类水鸟种类统计图 Fig. 3 Statistical map of the species of curlew waterbirds in coastal wetlands of Bohai Rim |

|
| 图 4 环渤海滨海湿地单位面积鸻鹬类水鸟数量统计图 Fig. 4 Statistical map of quantity per unit area of curlew waterbirds in coastal wetlands of Bohai Rim |
辽宁蛇岛老铁山湿地(P1)为基岩海岸, 海拔最高(87 m), 森林覆盖度35%, 鸻鹬类水鸟种类最少, 但单位面积鸻鹬类水鸟数量较多。河北曹妃甸湿地(P7)紧邻河北省唐山市曹妃甸区, 人类活动影响最大, 鸻鹬类水鸟种类、数量年际变化均最大。由于逐年的填海造地, 天津汉沽沿海湿地(P8)鸻鹬类水鸟种类年际变化大, 河北乐亭沿海湿地(P6)鸻鹬类水鸟数量年际变化大[30]。辽宁营口-大连沿海湿地(P2)、辽宁葫芦岛-锦州沿海湿地(P4)、河北北戴河湿地(P5)面积较小, 多为养殖池塘, 鸻鹬类水鸟种类、数量均不高。
各调查区的水鸟多样性差异明显。2016—2020年, 鸻鹬类水鸟多样性最高的区域为环渤海西南部、西部地区, 位列前四名的湿地依次为山东黄河三角洲湿地(P12)、河北沧州沿海湿地(P10)、山东滨州贝壳堤岛湿地(P11)、天津北大港沿海湿地(P9)(图 5)。辽宁蛇岛老铁山湿地(P1)、河北北戴河湿地(P5)、辽宁葫芦岛-锦州沿海湿地(P4)的鸻鹬类水鸟多样性最低。河北北戴河湿地(P5)、天津汉沽沿海湿地(P8)鸻鹬类水鸟种类年际变化大, 导致多样性年际变化最大。辽宁营口-大连沿海湿地(P2)、辽宁辽河口湿地(P3)、河北乐亭沿海湿地(P6)、河北曹妃甸湿地(P7)鸻鹬类水鸟多样性年均稳定在2左右。

|
| 图 5 环渤海滨海湿地鸻鹬类水鸟多样性统计图 Fig. 5 Statistical map of the diversity of curlew waterbirds in coastal wetlands of Bohai Rim |
经过反复拟合、评价、修正、再评价后得到了标准化系数修正模型(图 6), 拟合指数分析结果如表 3所示, 拟合指数总体表现较好, 均达到拟合指数要求, 认为从统计学角度经过修正后得到的环渤海滨海湿地鸻鹬类水鸟多样性与环境因子响应关系模型比较合理。潜变量及可测变量之间总得影响效果见图 6。

|
| 图 6 鸻鹬类水鸟多样性与环境因子响应关系修正模型 Fig. 6 Correction model for the relationship between diversity and environmental factors of curlew waterbirds |
| 指数名称 Indices name |
参考标准 Evaluation criterion |
评价结果 Results |
|
| 绝对拟合指数Absolute fitting index | x2/df | < 3 | 1.106 |
| GFI | > 0.9,越近1越好 | 0.993 | |
| RMSEA | < 0.05 | 0.042 | |
| 相对拟合指数Relative fit index | NFI | >0.9,越近1越好 | 0.982 |
| TLI | 0.979 | ||
| CFI | 0.998 | ||
| 精简指数Compact index | IFI | 0.998 | |
| AIC | 越小越好 | 29.106 | |
| ECVI | 越小越好 | 0.493 | |
| x2/df:卡方自由度比, GFI:拟合优度指数Goodness of Fit Index, RMSEA:近似误差均方根Root Mean Square Error of Approximation, NFI:赋范拟合指数Normed Fit Index, TLI:塔克-刘易斯指数Tucker-Lewis Index, CFI:比较拟合指数Comparative Fit Index, IFI:增量拟合指数Incremental Fit Index, AIC:讯息效标系数Akaike Information Criterion, ECVI:预期交叉验证指数Expected Cross-Validation Index | |||
2016—2020年辽宁辽河口湿地的多年平均气温最低, 为10.59℃;辽宁蛇岛老铁山湿地的多年平均降水量最低, 为451.2 mm, 其次为天津北大港沿海湿地、河北沧州沿海湿地、山东滨州贝壳堤岛湿地;其他湿地的年均温度及降雨量, 随所处纬度的增高而增高。温度、降雨两个可测变量对气候潜变量的影响路径系数分别为1.18和-0.34。气候对鸻鹬类水鸟多样性的直接影响为0.33, 间接影响为-0.26。辽宁蛇岛老铁山湿地的温度、降雨量均较低, 水鸟多样性最低。
浮游植物对环境变化反应敏感, 其动态可以良好地指示气候变化, 受温度的影响为0.31, 对鸻鹬类水鸟多样性的间接影响为0.34。浮游动物具有连接初级生产者和高级营养级的作用, 对鸻鹬类水鸟多样性的间接影响为0.26。大型底栖生物对鸻鹬类水鸟多样性的间接影响为0.38。食物对鸻鹬类水鸟多样性的直接影响为0.44。河北北戴河湿地及周边海水富营养化严重, 浮游植物密度(8714.67万个/m3)过高, 浮游动物密度(528个/m3)、生物量过高, 底栖动物密度(91.47个/m3)、生物量过低, 水鸟多样性低。
保护强度与食物的相关系数为0.68, 对鸻鹬类水鸟多样性的间接影响为0.3、直接影响为0.47。调查区覆盖18处自然保护地, 其中自然保护区11处、森林公园2处、地质公园2处、海洋公园2处、海洋特别保护区1处。国家级、省级、市级自然保护地分别12处、5处、1处。各级各类自然保护地的建设管理能有效保护鸟类多样性。
4 讨论鸟类多样性是潜变量与潜变量、潜变量与可测变量、可测变量与可测变量相互作用的结果, 结构方程模型的建立可以定量描述这一复杂关系。本研究根据环渤海滨海湿地鸻鹬类水鸟群落组成时空变化特征, 探索性地将结构方程模型引入了水鸟多样性与环境因子响应关系研究, 定量分析环境因子的影响力度, 运算、分析结果表明:气候、食物、保护强度对鸻鹬类水鸟多样性具有直接影响作用, 其中食物的直接影响力最大;保护强度不仅对鸻鹬类水鸟多样性存在直接影响, 此外通过食物对其产生间接影响, 合计影响力最大。今后需要强化自然保护地建设管理在维护提升鸟类物种多样性过程中的作用。建议:①调整河北南大港湿地和鸟类省级自然保护区保护等级, 提升至国家级;②将滨州贝壳堤岛与湿地国家级自然保护区周边集中连片湿地纳入保护区, 扩大保护区范围;③对山东黄河三角洲、辽宁辽河口覆盖的各级各类自然保护地进行优化整合, 确保水鸟多样性集中分布区及其所承载的景观、地质地貌得到有效保护。
此外, SEM模型拟合结果是由现状可测变量所支撑的, 是反映研究时段内的各变量响应关系, 对于其未来的发展趋势尚难以定论。对于不同时期、不同尺度和不同条件下的水鸟多样性与环境因子响应关系则需要补充完善相关变量来拟合, 从而使结论更具科学性和准确性。
5 结论环渤海地区12个典型滨海湿地, 共记录鸻鹬类水鸟7科51种, 其中旅鸟50种。全球极危物种1种, 濒危物种3种, 近危物种9种。国家一级保护鸟类2种, 国家二级保护鸟类8种。黑腹滨鹬、大滨鹬、黑尾塍鹬、灰鸻、斑尾塍鹬、白腰杓鹬、环颈鸻、红腹滨鹬个体数量最多。斑尾塍鹬(近危物种)、白腰杓鹬(近危物种)、大滨鹬(濒危物种)、环颈鸻的大部分迁徙种群在环渤海的滨海湿地中停歇。山东黄河三角洲、辽宁辽河口、天津北大港等河口湿地, 水鸟种类多, 单位面积水鸟数量较少。河北沧州沿海、山东滨州贝壳堤岛及其周边区域为环渤海地区湿地集中区, 水鸟种类较多。环境因子的影响强度由大到小依次为保护强度、食物、气候。
| [1] |
Fang J Y, Wang Z H, Zhao S Q, Li Y K, Tang Z Y, Yu D, Ni L Y, Liu H Z, Xie P, Da L J, Li Z Q, Zheng C Y. Biodiversity changes in the lakes of the Central Yangtze. Frontiers in Ecology and the Environment, 2006, 4(7): 369-377. DOI:10.1890/1540-9295(2006)004[0369:BCITLO]2.0.CO;2 |
| [2] |
Baillie S R, Peach W J. Population limitation in Palaearctic-African migrant passerines. Ibis, 1992, 134(S1): 120-132. |
| [3] |
Sillett T S, Holmes R T, Sherry T W. Impacts of a global climate cycle on population dynamics of a migratory songbird. Science, 2000, 288(5473): 2040-2042. DOI:10.1126/science.288.5473.2040 |
| [4] |
Studds C E, Deluca W V, Baker M E, King R S, Marra P P. Land cover and rainfall interact to shape waterbird community composition. PLoS One, 2012, 7(4): e35969. DOI:10.1371/journal.pone.0035969 |
| [5] |
Zhang Y M, Tan W Z, Zeng Q, Tian H T, Jia Y F, Lei G C, Wen L. Lake productivity and waterbird functional diversity across geographic and environmental gradients in temperate China. Ecology and Evolution, 2020, 10(20): 11237-11250. DOI:10.1002/ece3.6763 |
| [6] |
Rodenhouse N L, Holmes R T. Results of experimental and natural food reductions for breeding black-throated blue warblers. Ecology, 1992, 73(1): 357-372. DOI:10.2307/1938747 |
| [7] |
Donovan T M, Jones P W, Annand E M, Thompson Ⅲ F R. Variation in local-scale edge effects: mechanisms and landscape context. Ecology, 1997, 78(7): 2064-2075. DOI:10.1890/0012-9658(1997)078[2064:VILSEE]2.0.CO;2 |
| [8] |
Francesiaz C, Yohannes E, Besnard A, Sadoul N, Blanchon T, Béchet A. Foraging niche shift maintains breeding parameters of a colonial waterbird during range expansion. Ecology and Evolution, 2020, 10(4): 1988-1997. DOI:10.1002/ece3.6030 |
| [9] |
Stutchbury B J. Competition for winter territories in a neotropical migrant: the role of age, sex and color. The Auk, 1994, 111(1): 63-69. DOI:10.2307/4088505 |
| [10] |
Guan L, Lei J L, Zuo A J, Zhang H, Lei G C, Wen L. Optimizing the timing of water level recession for conservation of wintering geese in Dongting Lake, China. Ecological Engineering, 2016, 88: 90-98. DOI:10.1016/j.ecoleng.2015.12.009 |
| [11] |
Almeida B A, Sebastián-González E, dos Anjos L, Green A J. Comparing the diversity and composition of waterbird functional traits between natural, restored, and artificial wetlands. Freshwater Biology, 2020, 65(12): 2196-2210. DOI:10.1111/fwb.13618 |
| [12] |
Kleijn D, Cherkaoui I, Goedhart P W, van der Hout J, Lammertsma D. Waterbirds increase more rapidly in ramsar-designated wetlands than in unprotected wetlands. Journal of Applied Ecology, 2014, 51(2): 289-298. DOI:10.1111/1365-2664.12193 |
| [13] |
陈克林, 杨秀芝, 吕咏. 鸻鹬类鸟东亚-澳大利西亚迁飞路线上的重要驿站: 黄渤海湿地. 湿地科学, 2015, 13(1): 1-6. |
| [14] |
肖洋, 张路, 张丽云, 肖燚, 郑华, 欧阳志云. 渤海沿岸湿地生物多样性变化特征. 生态学报, 2018, 38(3): 909-916. |
| [15] |
陈莹, 谭坤, 马志军. 黄渤海湿地面积变化及其对鸻鹬类的影响//第十二届全国鸟类学术研讨会暨第十届海峡两岸鸟类学术研讨会论文摘要集. 杭州: 浙江省科学技术协会, 2013: 84-84.
|
| [16] |
赵宁, 夏少霞, 于秀波, 段后浪, 李瑾璞, 陈亚恒. 基于MaxEnt模型的渤海湾沿岸鸻鹬类栖息地适宜性评价. 生态学杂志, 2020, 39(1): 194-205. |
| [17] |
陈克林, 吕咏, 王琳, 陈颀, 吕宪国, 姜明. 中国环绕黄海和渤海的湿地春季水鸟多样性及其分布. 湿地科学, 2019, 17(2): 137-145. |
| [18] |
Hijmans R J, Cameron S E, Parra J L, Jones P G, Jarvis A. Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 2005, 25(15): 1965-1978. DOI:10.1002/joc.1276 |
| [19] |
冯威, 赵成章, 岳冉, 张翔, 金建鑫. 张掖国家湿地公园冬春季鸟类群落多样性和相似性分析. 生态学杂志, 2017, 36(8): 2224-2231. |
| [20] |
蒋志刚, 纪力强. 鸟兽物种多样性测度的G-F指数方法. 生物多样性, 1999, 7(3): 220-225. DOI:10.3321/j.issn:1005-0094.1999.03.010 |
| [21] |
邵明勤, 曾宾宾, 徐贤柱, 尚小龙, 戴年华. 鄱阳湖流域非繁殖期鸟类多样性. 生态学报, 2013, 33(1): 140-149. |
| [22] |
李慧, 汪景宽, 裴久渤, 李双异. 基于结构方程模型的东北地区主要旱田土壤有机碳平衡关系研究. 生态学报, 2015, 35(2): 517-525. |
| [23] |
成思敏, 王继军, 郭满才, 李茂森, 乔梅, 赵晓翠. 基于结构方程模型的陕北退耕区农业产业-资源系统耦合机制分析——以吴起县为例. 自然资源学报, 2018, 33(7): 1165-1178. |
| [24] |
Tidwell P R, Webb E B, Vrtiska M P, Bishop A A. Diets and food selection of female mallards and blue-winged teal during spring migration. Journal of Fish and Wildlife Management, 2013, 4(1): 63-74. DOI:10.3996/072012-JFWM-062 |
| [25] |
张强, 马克明, 李金亚, 张育新. 不同尺度下停歇点湿地对迁徙水鸟的影响研究综述. 生态学报, 2017, 37(8): 2520-2529. |
| [26] |
Traill L W, Bradshaw C J A, Brook B W. Satellite telemetry and seasonal movements of Magpie Geese (Anseranas semipalmata) in tropical northern Australia. Emu-Austral Ornithology, 2010, 110(2): 160-164. DOI:10.1071/MU09098 |
| [27] |
陈龙, 张美玲, 李凤丽, 于文颖. 2017年春季辽河口湿地鸟类群落结构和多样性. 湿地科学, 2019, 17(2): 146-151. |
| [28] |
柴子文, 雷维蟠, 莫训强, 阙品甲, 尚成海, 阳积文, 张正旺. 天津市北大港湿地自然保护区的鸟类多样性. 湿地科学, 2020, 18(6): 667-678. |
| [29] |
张孚允, 杨若莉. 中国鸟类迁徙研究. 北京: 中国林业出版社, 1997: 30-33.
|
| [30] |
颜凤, 李宁, 杨文, 乔亚军, 安树青. 围填海对湿地水鸟种群、行为和栖息地的影响. 生态学杂志, 2017, 36(7): 2045-2051. |