 2021, Vol. 41
2021, Vol. 41文章信息
- 楼一恺, 范忆, 戴其林, 王铮屹, 库伟鹏, 赵明水, 余树全
- LOU Yikai, FAN Yi, DAI Qilin, WANG Zhengyi, KU Weipeng, ZHAO Mingshui, YU Shuquan
- 天目山常绿落叶阔叶林群落垂直结构与群落整体物种多样性的关系
- Relationship between vertical structure and overall species diversity in an evergreen deciduous broad-leaved forest community of Tianmu Mountain Natural Reserve
- 生态学报. 2021, 41(21): 8568-8577
- Acta Ecologica Sinica. 2021, 41(21): 8568-8577
- http://dx.doi.org/10.5846/stxb202007301989
-
文章历史
- 收稿日期: 2020-07-30
- 修订日期: 2021-03-30





2. 国家林业和草原局华东调查规划设计院, 杭州 310019;
3. 浙江天目山国家级自然保护区管理局, 临安 311311
2. East China Inventory and Planning Institute, National Forestry and Grassland Administration, Hangzhou 310019, China;
3. Management Bureau of Tianmu Mountain National Nature Reserve, Lin' an 311311, China
生物多样性作为人类生存与发展的基础, 正以前所未有的速度丧失[1]。森林是全球陆地上生物多样性最丰富的群落, 已有的许多研究表明, 森林生物多样性丰富程度与森林生态系统的生产力、稳定性等功能密切相关。Franklin等指出具有高树种多样性的天然森林生态系统的长期生产力比具有低树种多样性的森林的长期生产力更高[2]。Tilman等的研究也发现, 群落结构对群落物种多样性以及生态系统功能有重要影响[3]。了解森林群落的生物多样性构成机制, 对森林生物多样性的保护以及可持续森林管理具有重要意义。
森林群落在垂直结构上的成层现象是群落的基本特征之一, 也导致了光照等生境条件在群落内部分化, 为更多物种的生存提供了生态位。桂旭君等研究发现森林群落在垂直结构上的成层现象是林内环境因子高异质性的关键之一[4]。Huang等发现热带森林的物种多样性受到森林垂直结构的显著影响[5]。Ishii等通过对温带森林生态系统结构的分析认为林层结构的复杂性可以提高林分生产力和群落结构的物种多样性[6]。因此, 森林群落垂直结构对群落物种多样性影响的研究成为群落生态研究的核心问题受到广泛关注。但是, 如何有效揭示群落垂直结构对群落整体物种多样性影响作用, 如何定量直接表达各林层对群落整体物种多样性贡献大小, 并未有系统有效分析方法, 本研究旨在开展一些有益探索。
常绿阔叶林是我国亚热带地区最具代表性的森林植被类型, 是结构层次最复杂、生物多样性最丰富的地带性植被类型之一[7]。在亚热带海拔1000—2000 m的中山地带, 亚热带常绿阔叶林由于适应低温环境而出现不同程度的落叶成分, 从而形成亚热带常绿落叶阔叶混交林, 亚热带常绿落叶阔叶混交林具有落叶阔叶林向常绿阔叶林之间过渡的特征, 对森林植被的恢复和发展具有重要意义[8]。已有学者分别对亚热带常绿落叶阔叶林垂直结构[9-13]、物种多样性[14-15]等进行研究。但是迄今为止, 对亚热带常绿落叶阔叶混交林群落垂直结构与群落整体物种多样性相互关系的研究还未见报道。本文旨以处于长期保护状态下的浙江省天目山1 hm2常绿落叶阔叶混交林样地调查数据为基础, 运用K-means聚类方法将DBH≥1 cm的个体根据树高划分为不同林层, 研究比较了各个林层的物种多样性特点;探索利用通径分析方法和决策系数定量计算各个林层物种多样性对群落整体物种多样性的直接作用和间接作用, 揭示了天目山常绿落叶阔叶林垂直结构与群落物种多样性间相互关系;分析稀有种、偶见种在物种多样性构成中作用。希望通过本研究能进一步从更多的视角深入探究森林群落物种多样性维持机制, 为亚热带同类森林生物多样性保护及可持续森林管理提供借鉴。
1 研究区概况天目山国家级自然保护区位于浙江省杭州市临安区境内, 北纬30°18′30″—30°21′37″N, 东经119°24′11″—119°27′11″E, 主峰仙人顶海拔1506 m[16]。保护区内土壤600—800 m以下为红壤带, 600—800 m至1200 m为黄壤带, 1200—1500 m为黄棕壤, 土壤腐质层明显[17-18]。受地形和海拔的影响, 保护区山麓(禅源寺)至山顶(仙人顶)气候垂直变化明显, 依次形成相应的垂直带谱, 海拔230—850 m为常绿阔叶林, 850—1100 m为常绿落叶阔叶混交林, 1100—1350 m为落叶阔叶林, 1350—1500 m为落叶矮林, 其中地带性植被为中亚热带常绿阔叶林[19-20]。
2 研究方法 2.1 调查方法2018年1月在天目山自然保护区1060 m处设置一个1 hm2(100 m×100 m)的监测样地, 其植被为常绿落叶阔叶混交林(119°25′47.154″E, 30°20′22.200″N), 中心点海拔1066.16 m;利用南方测绘NTS-300R全站仪以西南角作为原点, 东西向为横轴(x), 南北向为纵轴(y)将样地划分为25个20 m×20 m的大样方, 然后将每个20 m×20 m的大样方再划分为16个5 m×5 m的小样方, 以胸径DBH≥1 cm为木本植物起测径阶, 对每木进行编号、挂牌, 并记录其种名、树高、胸径、冠幅、枝下高、生长状况及位置坐标等, 胸径测量处均用油漆标记, 样地内共有DBH≥1 cm的木本植物4406株, 分属39科76属119种。
2.2 垂直结构划分运用K-means聚类方法将样地内全部个体(DBH≥1 cm)的树高进行分类以获得垂直结构层次和各层树高范围[4]。首先由于分类需要根据经验或其他条件事先设定分类数, 所以要先确定树高的最佳分类数。用于确定最佳分类数的常用指标SI值(林冠分层指数法)[21]、CCH值(树冠光竞争法)[22], 其中CCH=aCL+HW, 其中a为截止系数(a通常在0.3—0.5之间), CL为树冠长度, HW为枝下高;还可以根据不同分类情况下的组内平方和变化情况来判断[4]。
本文综合CCH的分层效应和组内平方和变化情况这两个指标来确定最佳分层数。选取由这两个指标综合确定的最佳分类组数代入K-means聚类中, 输出得到每一层的树高范围, 根据树高大小将样地内各树(DBH≥1 cm)划分到相应的层次中。
2.3 重要值及物种多样性群落的物种多样性是群落内物种丰富度和物种多度格局的综合体现, 常用于度量群落物种多样性的大小[23-25]。本文在20 m×20 m混交林样地地面空间尺度上采用常用的Margalef丰富度指数、Shannon-Wiener指数、Pielou指数以及Simpson指数研究群落的α多样性。
树种重要值:
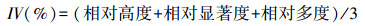
|
(1) |
Margalef丰富度指数:
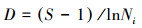
|
(2) |
Shannon-Weiner指数:
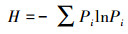
|
(3) |
Pielou指数:
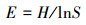
|
(4) |
Simpson指数:
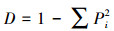
|
(5) |
式中, Ni为种i的个体数, N为群落中全部物种的个体数。S为物种数目, Pi为属于种i的个体在全部个体中的比例。
本文采用LSD法对各林层间α多样性指数大小进行多重比较, 判断差异是否显著。
2.4 林层间物种比较与种类划分利用Chazdon等提出的基于两个生境物种相对多度的多项式:CLAM(Classification Method)法分类模型, 结合混交林样地的实际情况将混交林样地各林层的物种组成进行两两比较[26]。两林层(林层A、林层B)的物种比较结果分为3个部分:两林层共有物种、林层A的特有种和林层B的特有种。
根据Hubbell和Foster对群落稀有种偶见种的划分, 本研究将样地内只有1株的物种定义为稀有种, 2—10株的为偶见种, 多于10株的为常见种[27]。
2.5 通径分析通径分析是通过自变量与因变量之间相关性分解的多元统计技术[28], 它可以将自变量对因变量的影响区分为直接重要性和间接重要性, 在此基础上计算决策系数, 能更好反映各个自变量对因变量的综合决定作用[29]。我们把群落整体物种多样性作为因变量, 各林层物种多样性作为自变量, 通过通径分析, 定量揭示不同林层物种多样性对群落整体物种多样性的直接重要性、间接重要性和综合决定作用。
本文采用R.4.0.3和Origin 8.0软件对数据进行数据处理, 分析作图。
3 结果分析 3.1 群落垂直层次结构 3.1.1 群落垂直层次结构划分根据CCH值和组内平方和将混交林样地的所有树种归入不同的高度层次。由图 1可知, 分类组数随截止系数a值的波动而变化, 大致可分为4—6层;同时, 在不同分类组数的组内平方和变化折线图中, 随分类组数的增大, 当组内平方和的大小变化趋缓时, 对应的分类组数可为最佳分类组数。根据不同分类组数时的组内平方和变化趋势图可知, 当分类组数为5时组内平方和大小变化开始趋于平缓, 5组之后基本不变。因此, 综合考虑这两个指数, 确定最优树高聚类组数为5类, 即将样地群落垂直结构划分为5层:即林层从下到上树高范围依次为[1.4, 4.2] m、[4.3, 7.6] m、[7.7, 13.4] m、[13.5, 21.5] m、[21.6, 36.5] m, 对应于传统垂直结构分类中的灌木层、亚冠层、林冠层, 进一步将林冠层划分为林冠下层、林冠中层和林冠上层(表 1)。

|
| 图 1 最佳分类组数判断指标:不同截止系数分层数和不同分类组数对应的组内平方和 Fig. 1 The judgment index of optimal classification group number: different cutoff coefficient number of layers and the sum of squares in different groups |
| 林层 Layer |
最低树高/m Minimum height |
最高树高/m Maximum height |
平均胸径/cm Average DBH |
多度(占比) Abundance/% |
物种丰富度(占比) Species richness/% |
特有种数(占比) Specialist% |
| 灌木层Shrub | 1.4 | 4.2 | 2.3±1.3 | 2847(64.62) | 93(78.15) | 22(56.41) |
| 亚冠层Subcanopy | 4.3 | 7.6 | 6.9±3.9 | 984(22.33) | 79(66.39) | 9(23.08) |
| 林冠下层Lower canopy | 7.7 | 13.4 | 16.5±7.6 | 378(8.58) | 48(40.34) | 5(12.82) |
| 林冠中层Middle canopy | 13.5 | 21.5 | 32.0±14.9 | 131(2.97) | 25(21.01) | 1(2.56) |
| 林冠上层Upper canopy | 21.6 | 36.5 | 47.5±23.2 | 66(1.50) | 12(10.08) | 2(5.13) |
| 总体Total | - | - | 6.1±9.4 | 4406 | 119 | 39 |
| DBH: 胸径Diameter at breast height | ||||||
从表 1可以看出, 以灌木层为参照, 随着林层垂直高度的上升, 混交林各林层间树高高度间隔先小后大, 群落各林层的平均胸径明显增大, 其中灌木层平均胸径为(2.3±1.3) cm, 林冠上层为(47.5±23.2) cm。各林层多度、物种丰富度和特有种数量则明显减小, 灌木层的多度和物种丰富度分别占混交林群落总多度和总物种丰富度的64.62%和78.15%, 亚冠层为22.33%和66.39%, 而林冠上层则仅有1.5%和10.08%;只在一个林层中出现的物种为该林层在整个混交林群落中的特有种, 其中灌木层拥有22个特有种, 占比达到56.41%, 林冠中层最少, 仅有1种。
从表 2可以看出, 通过各林层的主要树种种类(Ⅳ>5)、多度和平均胸径可以发现, 混交林群落不同林层的种类差异较大, 前三层以及整个混交林群落重要值最大的树种均为交让木(Daphniphyllum macropodum)。天目山常绿落叶阔叶混交林样地各林层特征如下:灌木层中植物个体数量和丰富度均最大, 该层内主要树种为交让木、大果山胡椒(Lindera praecox)、中国绣球(Hydrangea angustipetala)以及细叶青冈(Cyclobalanopsis gracilis)。亚冠层层内物种多为小乔木或乔木幼树, 主要树种为交让木、微毛柃(Eurya hebeclados)、细叶青冈和短尾柯(Lithocarpus brevicaudatus), 同时, 亚冠层树种个体数量分布十分不均匀, 交让木在亚冠层优势明显。林冠下层中多为中等乔木, 主要树种为交让木、细叶青冈、短尾柯、青钱柳(Cyclocarya paliurus)和杉木(Cunninghamia lanceolata)。林冠中层和林冠上层多为高大乔木, 其中林冠中层主要树种为青钱柳、杉木、细叶青冈、缺萼枫香(Liquidambar acalycina)、天目木姜子(Litsea auriculata)、蓝果树(Nyssa sinensis)和柳杉(Cryptomeria fortunei), 林冠上层物种数和多度最少, 主要树种为缺萼枫香、杉木、青钱柳、柳杉以及天目木姜子。
| 林层 Layer |
主要树种 Principal species |
多度 Abundance |
平均胸径/cm Average DBH |
重要值 Ⅳ/% |
| 灌木层 | 交让木Daphniphyllum macropodum | 436 | 3.0±1.8 | 19.24 |
| Shrub | 大果山胡椒Lindera praecox | 606 | 1.8±0.5 | 17.87 |
| 中国绣球Hydrangea angustipetala | 519 | 1.7±0.5 | 13.83 | |
| 细叶青冈Cyclobalanopsis gracilis | 129 | 2.8±2.5 | 6.03 | |
| 亚冠层 | 交让木Daphniphyllum macropodum | 367 | 7.7±3.7 | 39.92 |
| Subcanopy | 微毛柃Eurya hebeclados | 85 | 8.3±3.1 | 9.34 |
| 细叶青冈Cyclobalanopsis gracilis | 60 | 9.3±5.8 | 7.93 | |
| 短尾柯Lithocarpus brevicaudatus | 53 | 8.7±4.5 | 6.38 | |
| 林冠下层 | 交让木Daphniphyllum macropodum | 116 | 14.1±5.8 | 27.20 |
| Lower canopy | 细叶青冈Cyclobalanopsis gracilis | 88 | 20.8±8.2 | 27.17 |
| 短尾柯Lithocarpus brevicaudatus | 32 | 16.6±5.3 | 8.15 | |
| 青钱柳Cyclocarya paliurus | 19 | 22.4±7.8 | 6.39 | |
| 杉木Cunninghamia lanceolata | 22 | 17.0±6.1 | 5.99 | |
| 林冠中层 | 青钱柳Cyclocarya paliurus | 25 | 34.1±9.5 | 19.47 |
| Middle canopy | 杉木Cunninghamia lanceolata | 25 | 30.5±9.2 | 17.66 |
| 细叶青冈Cyclobalanopsis gracilis | 14 | 28.7±10.1 | 9.40 | |
| 缺萼枫香Liquidambar acalycina | 10 | 35.8±13.9 | 8.22 | |
| 天目木姜子Litsea auriculata | 9 | 37.4±6..7 | 7.33 | |
| 蓝果树Nyssa sinensis | 8 | 34.0±17.9 | 6.57 | |
| 柳杉Cryptomeria fortunei | 2 | 81.5±66.6 | 5.49 | |
| 林冠上层 | 缺萼枫香Liquidambar acalycina | 21 | 49.5±19.9 | 32.08 |
| Upper canopy | 杉木Cunninghamia lanceolata | 16 | 40.5±7.7 | 20.82 |
| 青钱柳Cyclocarya paliurus | 11 | 41.5±6.9 | 14.19 | |
| 柳杉Cryptomeria fortunei | 4 | 107.2±46.9 | 14.19 | |
| 天目木姜子Litsea auriculata | 5 | 41.0±2.7 | 6.54 | |
| 总体 | 交让木Daphniphyllum macropodum | 922 | 6.4±5.3 | 17.95 |
| Total | 细叶青冈Cyclobalanopsis gracilis | 291 | 10.8±10.4 | 8.91 |
| 大果山胡椒Lindera praecox | 634 | 1.9±0.5 | 7.79 | |
| 杉木Cunninghamia lanceolata | 149 | 14.2±14.5 | 6.92 | |
| 缺萼枫香Liquidambar acalycina | 45 | 32.9±24.5 | 6.13 | |
| 中国绣球Hydrangea angustipetala | 524 | 1.7±0.5 | 6.04 | |
| 青钱柳Cyclocarya paliurus | 63 | 27.8±14.3 | 5.67 | |
| Ⅳ: 重要值Important value | ||||
从表 3可以看出, 天目山常绿落叶阔叶混交林各林层物种多样性指数关系如下:沿林层向上, Shannon-Wiener指数呈逐渐降低趋势, 灌木层最高, 林冠上层最低, 且各林层之间差异显著(P < 0.05)。Pielou均匀度指数方面, 林冠中层的均匀度指数最高, 林冠上层次之, 灌木层最小, 其中灌木层和亚冠层、林冠下层、林冠中层和林冠上层之间差异显著。Simpson指数的变化趋势与Shannon-Wiener指数一致, 随林层的上升而降低, 灌木层最大, 林冠上层最小。
| 林层 Layer |
Shannon-Weiner指数 Shannon-Wiener index |
Pielou指数 Peliou evenness index |
Simpson指数 Simpson index |
| 灌木层Shrub | 2.187±0.295a | 0.748±0.090c | 0.808±0.078a |
| 亚冠层Subcanopy | 1.928±0.384b | 0.790±0.123c | 0.769±0.123a |
| 林冠下层lower canopy | 1.642±0.293c | 0.874±0.085b | 0.752±0.089ab |
| 林冠中层Middle canopy | 1.290±0.354d | 0.958±0.042a | 0.692±0.109b |
| 林冠上层upper canopy | 0.941±0.326e | 0.948±0.087a | 0.574±0.129c |
| 所有层All layers | 2.485±0.253 | 0.757±0.063 | 0.852±0.048 |
| 同一列中不同字母表示差异显著 | |||
从表 4可以看出, 通过天目山常绿落叶阔叶混交林样地各林层群落物种的比较可知, 沿林层向上, 各层之间的共有种数量(占比)逐渐减少, 特有种数量(占比)则逐渐增加, 其中灌木层和亚冠层共有种数量最多, 两个林层间共有种数量占比达到60%以上, 其余林层之间共有种数量占比均在40%以下, 林层之间物种差异明显;灌木层和林冠上层之间的特有种数量最多, 占比达到90%以上, 亚冠层和林冠上层的差异最大, 两者特有种占比达到92.94%。相邻两林层间物种共有种数量随林层增加呈递减趋势, 而特有种数量的占比大致呈递增趋势, 其中, 林冠下层和林冠中层的共有种数量在占比方面高于亚冠层和林冠下层, 特有种占比则低于亚冠层和林冠上层。
| 共有种数量(占比) Generalist |
特有种数量(占比) Specialist |
A特有种数量(占比) A specialist |
B特有种数量(占比) B specialist |
总计 Total |
|
| 灌木层和亚冠层 Shrub vs subcanopy |
65(60.75%) | 42(39.25%) | 28(26.17%) | 14(13.08%) | 107 |
| 灌木层和林冠下层 Shrub vs lower canopy |
36(34.29%) | 69(65.71%) | 57(54.28%) | 12(11.43%) | 105 |
| 灌木层和林冠中层 Shrub vs middle canopy |
17(16.83%) | 84(83.17%) | 76(75.25%) | 8(7.92%) | 101 |
| 灌木层和林冠上层 Shrub vs upper canopy |
8(8.25%) | 89(91.75%) | 85(87.63%) | 4(4.12%) | 97 |
| 亚冠层和林冠下层 Subcanopy vs lower canopy |
35(38.04%) | 57(61.96%) | 44(47.83%) | 13(14.13%) | 92 |
| 亚冠层和林冠中层 Subcanopy vs middle canopy |
17(19.54%) | 70(80.46%) | 62(71.26%) | 8(9.20%) | 87 |
| 亚冠层和林冠上层 Subcanopy vs upper canopy |
6(7.06%) | 79(92.94%) | 73(85.88%) | 2(7.06%) | 85 |
| 林冠下层和林冠中层 Lower canopy vs middle canopy |
19(35.19%) | 35(64.81%) | 29(53.70%) | 6(11.11%) | 54 |
| 林冠下层和林冠上层 Lower canopy vs upper canopy |
9(17.65%) | 42(82.35%) | 39(76.47%) | 3(5.88%) | 51 |
| 林冠中层和林冠上层 Middle canopy vs upper canopy |
9(32.14%) | 19(67.86%) | 16(57.14%) | 3(10.72%) | 28 |
通过表 5可以发现:灌木层x1对群落Shannon-Wiener指数的直接作用最大, 亚冠层x2次之, 林冠上层x5最小;间接作用中, 林冠上层x5最大, 总影响方面表现为x1>x5>x2>x3>x4。Pielou指数的影响方面, 灌木层x1的直接影响最大, 林冠中层最小, 总体影响方面表现为x1>x4>x5>x2>x3。Simpson指数方面, 灌木层x1的直接影响远高于其他四个林层, 总体影响方面表现为x1>x5>x2>x4>x3。
| 因变量(y) Dependent variable(y) |
自变量(x) InDependent variable(x) |
通径系数(直接) Path coefficient (Direct) |
通径系数(间接) Path coefficient (Indirect) |
总影响 Total effect |
决策系数 Decision coefficient |
| Shannon-Weiner指数 | x1 | 0.935 | -0.013 | 0.922 | 0.850 |
| Shannon-Wiener index | x2 | 0.260 | -0.047 | 0.213 | 0.043 |
| x3 | 0.078 | -0.085 | -0.007 | -0.007 | |
| x4 | 0.091 | -0.218 | -0.127 | -0.031 | |
| x5 | -0.058 | 0.317 | 0.259 | -0.033 | |
| Pielou指数 | x1 | 1.105 | -0.258 | 0.847 | 0.651 |
| Peliou evenness index | x2 | 0.514 | -0.555 | -0.041 | -0.304 |
| x3 | 0.056 | -0.478 | -0.422 | -0.050 | |
| x4 | -0.075 | 0.304 | 0.229 | -0.040 | |
| x5 | -0.046 | 0.210 | 0.164 | -0.017 | |
| Simpson指数 | x1 | 1.047 | -0.163 | 0.884 | 0.755 |
| Simpson index | x2 | 0.381 | -0.449 | -0.068 | -0.197 |
| x3 | 0.233 | -0.362 | -0.129 | -0.114 | |
| x4 | 0.077 | -0.191 | -0.114 | -0.023 | |
| x5 | 0.095 | 0.307 | 0.402 | 0.067 | |
| 表中x1为灌木层、x2为亚冠层、x3为林冠下层、x4为林冠中层、x5为林冠上层 | |||||
利用决策系数可以把各林层物种多样性对群落整体物种多样性的综合作用进行大小排序, 排序最大且为正数的变量为主要决策变量, 排序最小且为负数的变量为主要限制变量。从表 5中可以看出, 灌木层x1在Shannon-Wiener指数、Pielou指数以及Simpson指数上的决策系数均为最高, 故灌木层x1为群落物种多样性的主要决策变量, 林冠上层x5是Shannon-Wiener指数主要限制变量, 而亚冠层x2则是Pielou指数和Simpson指数的主要限制变量。
3.3.2 稀有种和偶见种对群落物种多样性的影响通过表 6各可以发现, 稀有种和偶见种在各林层的分布具体表现为:灌木层拥有数量最多的稀有种, 林冠中层不存在稀有种, 其在林层中的占比方面表现为林冠上层>灌木层>林冠下层>亚冠层>林冠中层。偶见种同样在灌木层中分布最多, 林冠上层最少, 偶见种在林层中的占比方面具体表现为亚冠层>林冠中层>灌木层>林冠下层>林冠上层;稀有种和偶见种在混交林和各林层种均占有大量的物种丰富度, 其在整个混交林群落中占有60%以上的丰富度, 在灌木层和亚冠层中达到50%, 在林冠下层、林冠中层以及林冠上层也达到40%以上。稀有种和偶见种的存在为群落整体和各林层的丰富度起到重要作用。
| 林层 Layer |
稀有种(占比) Rare species |
偶见种(占比) Occasional species |
常见种(占比) Common species |
总计 Total |
| 灌木层Shrub | 12(12.90%) | 37(39.79%) | 44(47.31%) | 93 |
| 亚冠层Subcanopy | 5(6.33%) | 36(45.57%) | 38(48.10%) | 79 |
| 林冠下层Lower canopy | 4(8.33%) | 17(35.42%) | 27(56.25%) | 48 |
| 林冠中层middle canopy | 0(0.00%) | 11(44.00%) | 14(56.00%) | 25 |
| 林冠上层upper canopy | 2(16.67%) | 3(25.00%) | 7(58.33%) | 12 |
| 总体Total | 23(19.33%) | 52(43.70%) | 44(36.97%) | 119 |
根据混交林样地内所有个体的树高进行聚类, 将天目山常绿落叶阔叶林样地群落的垂直结构划分为5个林层, 分别命名为灌木层、亚冠层、林冠下层、林冠中层、林冠上层。这与祝燕等将古田山常绿阔叶林划分为灌木层、亚乔木层和乔木层的结果相似[30]。天目山混交林群落各林层层次丰富, 是由于该群落自1956年以来一直处于保护状态下, 发育时间长, 形成了复杂垂直空间结构。在一些研究中, 研究者也常根据经验以5 m或10 m为间隔划分垂直各林层[31]。K-means聚类算法是目前应用最广泛的划分算法之一, 在众多学科领域被广泛研究和应用。本研究中利用CCH的分层效应和组内平方和变化情况对混交林群落进行K-means聚类的划分, 各林层间树高高度间隔先小后大, 其中灌木层层高2.8 m, 而林冠上层层高14.9 m, 相比利用5 m或10 m树高间隔划分森林垂直结构具有更高的合理性。
4.2 多层结构在维持构建森林物种多样性中具有关键作用根据CLAM法分类模型对混交林群落各林层树种进行分类, 沿林层向上, 各层之间共有种数量逐渐减少, 特有种数量逐渐增加, 这是由于灌木层和亚冠层中树种的局限性所导致的。相邻两林层间物种共有种和特有种在数量上也大致呈递减和递增趋势, 其中灌木层和亚冠层共有种占比超过60%, 这是因为亚冠层的树种多为小乔木, 与灌木层相似性较大;而亚冠层、林冠下层、林冠中层、林冠上层之间的特有种占比在55%以上, 这是由于群落垂直方向上的成层现象导致光照等生境条件在群落内部分化, 提高了环境因子的异质性, 环境的高异质性为混交林中更多物种的生存提供了可能。同时林层间物种组成的差异性提高了群落的物种多样性, 灌木层与亚冠层中的乔木树幼树(细叶青冈、杉木和青钱柳)为林冠下层、林冠中层和林冠上层源源不断的提供优势种物种, 为保持整个群落的多样性起到重要作用。这与Ishii等研究温带森林群落林层结构时发现了复杂的冠层结构促进了群落的物种多样性的结果相一致[6]。
4.3 灌木层在维持构建森林物种多样性中具有决定作用在天目山混交林样地的5个林层中, 灌木层的Shannon-Weiner指数以及Simpson优势度指数远高于其他林层, 通过不同林层物种多样性对群落整体物种多样性的通径分析可以发现, 灌木层物种多样性对混交林群落整体物种多样性的贡献最大, 灌木层对群落整体Shannon-Wiener指数、Pielou指数以及Simpson指数上的决策系数分别达到0.845、0.651、0.755, 远高于其它林层, 是维持整个群落物种多样性的主要决策变量。灌木层的发育程度会对混交林群落物种组成的复杂程度、均匀程度以及群落的多样性均产生决定性的影响。反过来灌木层的物种数目又受到林冠下层、林冠中层和林冠上层植株数量的影响, 从25个400m2样方研究发现(图 2), 林冠层密度越大, 灌木层的物种数目越少, 复杂程度越低。相关研究表明由于林冠层植株密度的不同导致光照、水分等差异, 影响了灌木层的发育[32-33]。因此, 我们认为在森林保护和管理中, 适当的控制林冠层的植株密度是有利于灌木层的发育, 对森林群落整体物种多样性的构建与维持能起到促进作用。

|
| 图 2 天目山混交林灌木层物种数目与林冠层植株密度的关系 Fig. 2 The relationship between number of species of shrub layer and canopy layer density in mixed forest of Tianmu Mountain |
稀有种和偶见种是群落中出现频率很低的物种, 过去在森林群落物种多样性研究中常常被忽视或没有引起足够重视。通过对天目山混交林群落中稀有种和偶见种共计75种267个个体分析发现, 这些稀有种和偶见种占了群落总物种数的63.03%, 但只占群落总个体数的6.06%, 从表 7可以看出, 稀有种和偶见种的缺失对丰富度指数的影响最大, 会导致各林层丰富度指数下降达40%以上;对Shannon-Wiener指数的影响次之, 但也可让林冠层Shannon-Wiener指数下降17%—28%不等;对Simpson指数的影响相对较小。可见天目山混交林群落中稀有种和偶见种对群落整体物种多样性的维持构建具有特殊作用。同时由于稀有种和偶见种的存在也可为群落应对环境变化提供更多的适应机会, 有助于群落应对外界的干扰。这与周文嵩对热度常绿季雨矮林的研究也发现稀有种和偶见种的数量变化与群落的Shannon-Wiener指数以及Simpson指数呈正相关相类似[34]。
| 林层 Layer |
丰富度指数下降百分比/% Richness index decline percentage |
Shannon-Weiner指数下降百分比/% Shannon-Wiener index decline percentage |
Simpson指数下降百分比/% Simpson index decline percentage |
| 灌木层Shrub | 52.99 | 6.69±0.62 | 2.12±0.24 |
| 亚冠层Subcanopy | 51.93 | 12.99±1.83 | 6.72±1.47 |
| 林冠下层Lower canopy | 43.93 | 17.20±3.37 | 10.11±2.79 |
| 林冠中层middle canopy | 44.14 | 21.68±3.10 | 11.84±1.95 |
| 林冠上层upper canopy | 44.18 | 28.10±5.84 | 19.04±4.87 |
| 总体Total | 63.29 | 7.92±0.73 | 2.10±0.22 |
综上所述, 天目山常绿落叶阔叶混交林样地DBH≥1 cm的木本植物在垂直层次上可划分为灌木层、亚冠层、林冠下层、林冠中层和林冠上层5个层次, 群落垂直层次结构与物种多样性丰富, 随林层向上各林层平均胸径明显增加, 物种丰富度、多度、特有种数量、Shannon-Weiner指数和Simpson指数不断减小。多层结构在维持构建森林物种多样性中具有关键作用, 灌木层在维持构建森林物种多样性中具有决定作用, 稀有种和偶见种在维持构建森林物种多样性中具有特殊作用。
| [1] |
马克平. 未来十年的生物多样性保护目标. 生物多样性, 2011, 19(1): 1-2. |
| [2] |
Franklin J F, Perry D A, Schowaltr M E, Harmon M E, Mckee A, Spies T A. Importance of ecological diversity in maintaining long-term site productivity//Perry D A, Meurisse R, Thomas B, Miller R, Boyle J, Means J, Perry CR, Powers RF, eds. Maintaining the Long-Term Productivity of Pacific Northwest Forest Ecosystems. Portland: Timber Press, 1989: 82-97.
|
| [3] |
Tilman D, Knops J, Wedin D, Reich P, Ritchie M, Siemann E. The influence of functional diversity and composition on ecosystem processes. Science, 1997, 277(5330): 1300-1302. DOI:10.1126/science.277.5330.1300 |
| [4] |
桂旭君, 练琚愉, 张入匀, 李艳朋, 沈浩, 倪云龙, 叶万辉. 鼎湖山南亚热带常绿阔叶林群落垂直结构及其物种多样性特征. 生物多样性, 2019, 27(6): 619-629. |
| [5] |
Huang W D, Pohjonen V, Johansson S, Nashanda M, Katigula M I L, Luukkanen O. Species diversity, forest structure and species composition in Tanzanian tropical forests. Forest Ecology and Management, 2003, 173(1/3): 11-24. |
| [6] |
Ishii H T, Tanabe S, Hiura T. Exploring the relationships among canopy structure, stand productivity, and biodiversity of temperate forest ecosystems. Forest Science, 2004, 50(3): 342-355. |
| [7] |
丁圣彦, 宋永昌. 常绿阔叶林植被动态研究进展. 生态学报, 2004, 24(8): 1765-1775. |
| [8] |
唐青青, 黄永涛, 丁易, 臧润国. 亚热带常绿落叶阔叶混交林植物功能性状的种间和种内变异. 生物多样性, 2016, 24(3): 262-270. |
| [9] |
丁晖, 方炎明, 杨青, 陈晓, 袁发银, 徐辉, 何立恒, 严靖, 陈婷婷, 余朝健, 徐海根. 武夷山中亚热带常绿阔叶林样地的群落特征. 生物多样性, 2015, 23(4): 479-492. |
| [10] |
游诗雪, 张超, 库伟鹏, 朱弘, 赵明水, 余树全. 1996-2012天目山常绿落叶阔叶混交林乔木层群落动态. 林业科学, 2016, 52(10): 1-9. DOI:10.11707/j.1001-7488.20161001 |
| [11] |
姚良锦, 姚兰, 易咏梅, 艾训儒, 冯广, 刘峻城, 林勇, 黄伟, 丁易, 臧润国. 湖北七姊妹山亚热带常绿落叶阔叶混交林的物种组成和群落结构. 生物多样性, 2017, 25(3): 275-284. |
| [12] |
秦运芝, 张佳鑫, 刘检明, 刘梦婷, 万丹, 吴浩, 周阳, 孟红杰, 肖之强, 黄汉东, 徐耀粘, 卢志军, 乔秀娟, 江明喜. 湖南八大公山25ha常绿落叶阔叶混交林动态监测样地群落组成与空间结构. 生物多样性, 2018, 26(9): 1016-1022. |
| [13] |
Bai C, You S X, Ku W P, Dai Q L, Wang Z Y, Zhao M S, Yu S Q. Life form dynamics of the tree layer in evergreen and deciduous broad-leaved mixed forest during 1996-2017 in Tianmu Mountains, eastern China. Silva Fennica, 2020, 54(2): 10167. |
| [14] |
王月霞, 金毅, 吴初平, 翁东明, 叶立新, 陈德良, 余建平, 刘金亮, 仲磊, 于明坚. 浙江省主要亚热带森林群落类型物种和谱系水平的α和β多样性比较. 生物多样性, 2016, 24(8): 863-874. |
| [15] |
胡芳, 曾馥平, 杜虎, 彭晚霞, 张芳, 谭卫宁, 宋同清. 桂西北喀斯特常绿落叶阔叶混交林物种多样性分布格局的尺度效应. 生态学报, 2018, 38(17): 6074-6083. |
| [16] |
商侃侃, 陈波, 达良俊. 浙江天目山孑遗落叶阔叶林群落分异格局及其地形解释. 生态环境学报, 2013, 22(2): 213-221. DOI:10.3969/j.issn.1674-5906.2013.02.007 |
| [17] |
周秀佳, 马炜梁, 刘永强. 西天目山森林植被类型及其分布规律. 生态学杂志, 1987, 6(3): 17-20. |
| [18] |
刘茂春. 西天目山森林植被的研究——南坡的森林植被分类. 浙江林学院学报, 1991, 8(1): 13-24. |
| [19] |
蔡壬侯. 浙江省森林植被的水平分布与垂直分布. 杭州大学学报, 1988, 15(3): 344-350. DOI:10.3321/j.issn:1008-9497.1988.03.015 |
| [20] |
汪岱华, 王幼芳, 左勤, 李敏, 吴文英, 黄建花, 赵明水. 浙江西天目山主要森林类型的苔藓多样性比较. 植物生态学报, 2012, 36(6): 550-559. |
| [21] |
Baker P J, Wilson J S. A quantitative technique for the identification of canopy stratification in tropical and temperate forests. Forest Ecology and Management, 2000, 127(1/3): 77-86. |
| [22] |
郑景明. 森林群落垂直层次划分方法. 林业科技通讯, 2006, 6(4): 4-6. |
| [23] |
MacArthur R. Fluctuations of animal populations and a measure of community stability. Ecology, 1955, 36(3): 533-536. DOI:10.2307/1929601 |
| [24] |
马克平, 黄建辉, 于顺利, 陈灵芝. 北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数. 生态学报, 1995, 15(3): 268-277. |
| [25] |
陈廷贵, 张金屯. 十五个物种多样性指数的比较研究. 河南科学, 1999, 17(S1): 55-57, 71-71. |
| [26] |
Chazdon R L, Chao A, Colwell R K, Lin S, Norden N, Letcher S G, Clark D B, Finegan B, Arroyo J P. A novel statistical method for classifying habitat generalists and specialists. Ecology, 2011, 92(6): 1332-1343. DOI:10.1890/10-1345.1 |
| [27] |
Hubbell S P, Foster R B. Commonness and rarity in a neotropical forest: implications for tropical tree conservation//Soulé M E, ed. Conservation Biology: the Science of Scarcity and Diversity. Sunderland: Sinauer Associates, 1986: 205-231.
|
| [28] |
宋小园, 朱仲元, 刘艳伟, 赵宏瑾. 通径分析在SPSS逐步线性回归中的实现. 干旱区研究, 2016, 33(1): 108-113. |
| [29] |
袁志发, 周静芋, 郭满才, 雷雪芹, 解小莉. 决策系数——通径分析中的决策指标. 西北农林科技大学学报: 自然科学版, 2001, 29(5): 131-133. |
| [30] |
祝燕, 赵谷风, 张俪文, 沈国春, 米湘成, 任海保, 于明坚, 陈建华, 陈声文, 方腾, 马克平. 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构. 植物生态学报, 2008, 32(2): 262-273. |
| [31] |
陈亚锋, 余树全, 严晓素, 伊力塔, 包春泉. 浙江桐庐3种森林类型群落结构. 浙江农林大学学报, 2011, 28(3): 408-415. |
| [32] |
陈高, 代力民, 周莉. 受干扰长白山阔叶红松林林分组成及冠层结构特征. 生态学杂志, 2004, 23(5): 116-120. |
| [33] |
田平, 韩海荣, 康峰峰, 程小琴, 朱江, 周文嵩. 密度调整对太岳山华北落叶松人工林冠层结构及林下植被的影响. 北京林业大学学报, 2016, 38(8): 45-53. |
| [34] |
周文嵩. 海南铜鼓岭热带常绿季雨矮林偶见种分布格局及其对生物多样性计算的影响研究[D]. 海口: 海南大学, 2014.
|