 2021, Vol. 41
2021, Vol. 41文章信息
- 郭佳佳, 张雷廷, 车建芳, 焦海华, 茹文明, 白志辉
- GUO Jiajia, ZHANG Leiting, CHE Jianfang, JIAO Haihua, RU Wenming, BAI Zhihui
- 太行山南段连翘群落结构及其影响因子研究
- Analysis of Forsythia suspensa community structure and influencing factors in the southern section of Taihang Mountain
- 生态学报. 2021, 41(21): 8589-8601
- Acta Ecologica Sinica. 2021, 41(21): 8589-8601
- http://dx.doi.org/10.5846/stxb201911242543
-
文章历史
- 收稿日期: 2019-11-24
- 网络出版日期: 2021-07-06





2. 长治学院生物科学与技术系, 长治 046011;
3. 中国科学院生态环境研究中心, 北京 100085;
4. 湖南师范大学附属光琇医院, 长沙 410013;
5. 太行山生态与环境研究所, 长治 046011
2. Department of Biological Sciences and Technology, Changzhi University, Changzhi 046011, China;
3. Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China;
4. Department of Basic Medicine, School of Medicine, Hunan Normal University, Changsha 410013, China;
5. Taihang Mountain Institute of ecology and environment, Changzhi 046011, China
连翘(Forsythia suspensa)为木犀科(Oleaceae)、连翘属(Forsythia)多年生落叶灌木, 是我国一味重要的传统中药。在山西境内, 连翘广泛分布于太行山南段、太岳山和吕梁山[1]。有关连翘组份的药用价值、生理活性及其提取方法的研究较多, 如Zhang等[2]报道了连翘提取物的抗氧化和消炎特性。Yuan等[3]探讨了连翘叶的主要活性化合物的提取、纯化方法及其抗氧化活性。王建华等[4]探讨了光照强度对连翘叶片光合特性、叶绿素荧光参数的影响, 揭示了连翘耐阴性强且能够适应多种光环境的植物生长特性。郎莹等[5]通过受控温室内盆栽实验探讨了连翘光合作用对土壤水分的响应。胡贝娟等[6]对山西太岳山连翘群落优势种种间关联性进行了定量研究。张玲[7]应用数量生态学的方法, 从群系、种群生态学的角度, 探讨了分布在山西的吕梁山、太岳山和中条山的连翘种群特征。韩虹[8]对山西历山、蟒河自然保护区连翘群落的数量排序、物种多样性及群落优势种的种间关系进行了研究。李宝堂等[9]探讨了山西历山自然保护区连翘群落及其影响因素。但目前, 有关影响太行山南段连翘群落结构的环境因子的研究还未见报道。
地处太行山南段的陵川县是我国道地药材连翘的主产地之一。近年来, 被山西省列为连翘种植基地和种植示范园区。从产业发展的角度来看, 目前该区还缺少大力发展连翘产业的技术支撑, 如对环境条件(包括地形、地貌、温度变化、土壤肥力)以及人类活动等对连翘生长及其药用品质影响的了解还很少。规模化人工种植时山地高度、坡向的选择、施肥种类、时间与方式等方面还缺少科学化种植与管理技术的指导。本研究以该区广泛分布的连翘群落为研究对象, 通过野外样方调查方法, 探讨连翘群落分布格局, 揭示影响连翘分布的主要环境因子, 发现制约连翘生长的环境因素, 为该区基地化连翘种植、管理提供理论指导和技术支撑。
1 研究区概况研究区位于太行山南段的陵川县境内, 以石山、土石丘陵地形为主, 平均海拔1058 m。属典型大陆性气候, 年平均降水量在600—700 mm之间, 年平均气温7.9 ℃左右。该区域的群落类型主要包括森林群落、灌丛群落、灌草丛和草丛群落, 其中, 森林群落的多样性较高。区域内分布的树种, 乔木层优势树种主要是辽东栎(Quercus wutaishanica)、油松(Pinus tabuliformis)、山杨(Populus davidiana)等;灌木层主要是连翘(Forsythia suspensa)、三裂绣线菊(Spiraea trilobata)、黄刺玫(Rosa xanthina )和胡枝子(Lespedeza bicolor)等;草本层主要是披针叶苔草(Carex lanceolata)、蛇莓(Duchesnea indica)和小红菊(Dendranthema chanetii)等。该区连翘多成片分布形成典型的灌木丛群落[10-11]。
2 研究方法 2.1 样方设置与样品采集在研究区选择连翘成片分布的灌丛群落和森林群落为研究对象。于2017年7月, 根据海拔高度、坡形、坡向和连翘的分布状况, 设置60个5 m×5 m的大样方(图 1), 每个大样方内设置2个1 m×1 m的草本样方。利用生态学计数方法, 测量、记录样方内灌木和草本的丛数、盖度、高度;对胸径(DBH)大于4 cm的乔木记录种名、胸径、树高、冠幅(东西、南北)等生长指标。利用GPS定位样地的经、纬度、海拔高度、坡度、坡位和坡向。60个样方分布于东经113.24°—113.32°, 北纬35.40°—35.46°, 海拔(H)在1000—2000 m, 坡向指数(TRP)在0.1—0.9之间。以网格方式采集土壤样品, 在每个样方内采集25—30个样点土壤。土壤采集时先去除表层枯枝落叶层, 用不锈钢铲子取表层(0—25 cm)土壤, 然后混合记作一个样方的样品。将样品土壤去除碎石和植物残体, 运送回实验室分析[12]。

|
| 图 1 样方分布及坡向 Fig. 1 Distribution and slope direction of sample plots |
土壤样品经自然风干研磨过0.5 μm筛, 用重铬酸钾氧化外加热法测定土壤有机质含量;凯氏定氮法测定土壤全氮、有效氮含量;钼锑抗比色法测定土壤全磷、有效磷含量;火焰光度法测定土壤全钾、速效钾含量;pH值采用1 ∶ 2.5土水比悬液酸度计测定;电导率(EC)采用1 ∶ 5土水比浸提液法[13-14]。
2.3 数据处理与分析 2.3.1 重要值计算物种数据采用能较全面反映物种在群落中的地位和作用的物种的重要值表示[15-16]。计算公式:

|
(1) |
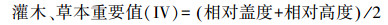
|
(2) |
坡向用坡向指数(TRP)表示。由GPS测量得到的0—360°的坡向度数值, 利用公式转换成0—1之间的指数值(TRP), 用于MRT、冗余分析(RDA)[17]。转换公式:

|
(3) |
式中, TRP为坡向指数, 变化范围在0—1之间, 数值越大, 表示生境越向阳、干热;aspect为坡向角度。所有样方的TRP值见图 1。
2.3.3 冗余分析通过(Redundancy analysis, RDA)分析模型对物种与环境因子的关系进行排序;用蒙特卡罗(Monte Carlo)检验, 度量各个环境因子与物种之间关系的显著性。RDA排序图中, 环境变量用带有箭头的实线表示, 箭头连线与排序轴的夹角表示该环境因子与排序轴相关性的大小, 箭头所示的方向为该因子的发展趋势。
数据分析在SPSS 17.0中进行, 利用MATLAB R 2016做图比较不同样方土壤性质的差异性, 利用软件R 3.3.3采用多元回归树(MRT) 对该区分布的连翘群落进行群落划分[18-19], 利用Canoco 4.5的排序结果完成冗余分析。
3 结果与分析 3.1 连翘群落类型划分依据公式(1)、(2)计算各群落物种的重要值。以地形因子:海拔高度(H)、坡向指数(TRP), 土壤因子:全氮(TN)、全磷(TP)、全钾(TK)、有效氮(AN)、有效磷(AP)和速效钾(AK)的含量设为自变量, 物种的重要值为因变量, 对60个样方建立多元回归树(MRT)。利用交叉验证对分类结果进行剪枝, 在保证误差和树的规模都尽可能小的条件下[20-21], 根据1-SE规则:保证通过交叉验证获得的预测误差在尽量小(最小的误差±一个相应标准误差)的范围内, 选取规模最小的树[22-23], 结合宋永昌[24]对群丛的命名原则, 对连翘群落进行命名(图 2)。

|
| 图 2 连翘群落划分多元回归树图 Fig. 2 Multivariate regression tree for Forsythia suspensa community classification CV Error: 交叉验证相对误差Cross-validation relative errors; Error: 相对误差Relative error; SE: 标准误Standard error; TK: 土壤全钾含量Total soil potassium; n: 各群丛包含的样方数; 17.4、0.14、1.23、0.333、0.283为各群丛物种重要值 |
由图 2可知, 海拔、土壤全钾、坡向形成了3个节点, 将连翘群落分型为5个群丛:
群丛Ⅰ: 连翘(Forsythia suspensa)+三裂绣线菊(Spiraea trilobata)-披针叶苔草(Carex lanceolata)群丛。主要分布在海拔1090—1330 m处。群落所有物种重要值的平均值为17.4, 连翘重要值为16.0, 三裂绣线菊重要值为6.97, 披针叶苔草重要值为6.30。
群丛Ⅱ: 连翘(Forsythia suspensa)+白刺花(Sophora davidii)-披针叶苔草(Carex lanceolata)+铁杆蒿(Tripolium vulgare)群丛。分布在海拔1450 m处。群丛物种重要值的平均值为0.140, 连翘重要值为0.660, 白刺花重要值为0.247, 披针叶苔草重要值为0.417, 铁杆蒿重要值为0.138。
群丛Ⅲ: 辽东栎(Quercus wutaishanica)-连翘(Forsythia suspensa)+三裂绣线菊(Spiraea trilobata)-披针叶苔草(Carex lanceolata)+白头翁(Pulsatilla chinensis)群丛。分布在海拔1340—1450 m处, 丛中物种重要值的平均值为0.283, 其中, 连翘重要值为0.779, 三裂绣线菊重要值为0.484, 披针叶苔草重要值为0.268, 白头翁重要值为0.101。
群丛Ⅳ: 油松(Pinus tabuliformis)-连翘(Forsythia suspensa)-披针叶苔草(Carex lanceolata)群丛。主要分布在海拔1460—1480 m的油松林边缘地区, 连翘植株高大。群丛物种重要值的平均值为0.333, 其中, 油松为2.05, 连翘为1.36, 披针叶苔草为0.839。
群丛Ⅴ: 连翘(Forsythia suspensa)+黄刺玫(Rosa xanthina)-披针叶苔草(Carex lanceolata)群丛。海拔1400—1470 m之间。群丛物种重要值的平均值为1.23, 其中, 连翘重要值为2.39, 黄刺玫为1.19, 披针叶苔草为1.49。
同时可知, 海拔(H)、土壤全钾(TK)、坡向(TRP)3个因子相互作用, 在四个层级构成了该区连翘群落分型主导因子。H高度1330 m是群落分型的一级主导因子;当H高度大于1330 m其群落结构的分型主要取决于土壤TK的含量, 构成了二级分型主导因子, TK含量大于19.07 mg/kg的群落为典型的Ⅱ群丛, 其优势物种主要有连翘、黄刺玫、披针叶苔草、铁杆蒿;在海拔大于1330 m, 土壤TK含量低于19.07 mg/kg和坡向(TRP)构成了三级分型主导因子, TRP小于0.2775的样方, 群落结构为群丛Ⅲ, 其优势物种主要包括辽东、栎、连翘、三裂绣线菊、披针叶苔草、白头翁;在H大于1330 m, TRP大于0.2775的样方, TK含量小于18.47 mg/kg, 是群落分型的四级主导因子, 群落为群丛Ⅴ, 连翘、黄刺玫、披针叶苔草为优势种。总体来看H(1330 m), TK(19.07、18.47 mg/kg)和TRP(0.2775)是群落结构分型的节点, 表明海拔、坡向和土壤钾含量对连翘的分布有较大的影响。
3.2 土壤性质根据2.2中的方法, 分析土壤的pH、有机质(TOC)、全氮(TN)、全磷(TP)、全钾(TK)、有效氮(AN)、有效磷(AP)和速效钾(AK)的含量。以纬度(N)为X轴, 以经度(E)为Y轴, 以海拔高度(H)为Z轴, 以色彩表征不同的化学特征值, 样方土壤中各组分的含量情况如图 3—10所示。

|
| 图 3 不同样方的土壤酸碱度 Fig. 3 Soil pH of the sample plots |

|
| 图 4 不同样方的土壤有机质 Fig. 4 Soil organic matter (TOC) of the sample plots |

|
| 图 5 不同样方的土壤全磷 Fig. 5 Soil total phosphorus (TP) of the sample plots |

|
| 图 6 不同样方的土壤有效磷 Fig. 6 Soil available phosphorus (AP) of the sample plots |

|
| 图 7 不同样方的土壤全钾 Fig. 7 Soil total potassium (TK) of the sample plots |

|
| 图 8 不同样方的土壤速效钾 Fig. 8 Soil available potassium (AK) of the sample plots |

|
| 图 9 不同样方的土壤总氮 Fig. 9 Soil total nitrogen (TN) of the sample plots |

|
| 图 10 不同样方的土壤有效氮 Fig. 10 Soil available nitrogen (AN) of the sample plots |
由图 3可知, 所有样方的土壤pH范围在7.0—9.0之间, 大部分样方大于7.5呈现弱碱性或碱性, 且海拔高度对其有一定的影响, 海拔越高酸度越大(图 3)。样方土壤有机质含量在2.0%—15.0%之间, 呈现不均匀分布状态, 大多数样方的含量在2.0%—5.0%之间, 随经纬度的变化不明显, 但海拔高度越高, 有机质含量越少(图 4);土壤总磷的含量相对比较均匀, 在0.03%—0.86%之间, 且与经、纬度、海拔高度没有明显的差异(图 5), 不同样方间有效磷的含量差异较大, 大多数样方在10—80 mg/kg之间(图 6);总的钾含量在区域内分布相对比较一致, 整体来看含量在1.0%—2.0%之间(图 7), 速效钾的含量在100—300 mg/kg之间, 不均匀性分布(图 8);土壤总氮含量在0.1%—1.0%之间, 有随海拔高度增高而增大的趋势(图 9), 而有效氮在区域内的分布不均匀, 碎片化分布特点较为明显, 区域的含量在50—1000 mg/kg之间(图 10)。
3.3 地形因子与群落数量特征值的梯度分析利用Cannoco软件, 先对物种数据进行降趋势对应分析(Detrended correspondence analysis, DCA), 依据第一排序轴的梯度值(Lengths of gradient)选择分析模型。该梯度检测值小于3, 因此, 该分析选择冗余分析模型(Redundancy analysis, RDA), 并进行了Monte Carlo置换检验。以样方的物种数量和连翘物种的重要值与环境地形因子构成数据矩阵, 应用冗余分析(RDA)方法, 输出RDA二维排序图。2个排序轴分别解释了变量的95.5%和4.5%, 解释了总变量的100%(表 1)。分别以物种—地形因子、样方—地形因子做图(图 11—12)。图中实线箭头表示环境因子, 虚线表示物种因子, 箭头连线的长短表示群落的分布与该环境因子相关性的大小, 箭头连线与排序轴夹角的大小表示环境因子与排序轴相关性的大小, 夹角小说明相关性大, 箭头所在的象限表示方位环境因子与排序轴的正负相关性。环境变量对群落结构影响的显著性检验结果(表 1)。Pr表示显著性大小, 当Pr为0.001, 代表极显著, 说明排序结果可以接受环境因子对连翘物种分布的解释量。由表 1可知, 纬度(N)、坡向指数(TRP)、海拔高度(H)对连翘物种分布的影响极显著。
| 排序轴Axis | 轴1 Axis1 | 轴2 Axis2 | r2 | Pr |
| 北纬Northern latitude(N) | 0.995 | 0.102 | 0.343 | 0.001 *** |
| 东经East longitude(E) | 0.016 | -0.999 | 0.057 | 0.186 |
| 坡向系数Coefficient of slope(TRP) | 0.9031 | -0.429 | 0.230 | 0.001 *** |
| 海拔高度Altitude(H) | 0.998 | -0.068 | 0.480 | 0.001 *** |
| 特征值Eigenvalue | 20.8 | 19.8 | ||
| 解释量Interpretation/% | 95.5 | 4.50 |

|
| 图 11 连翘群落物种特征与样方地形因子的冗余分析 Fig. 11 Redundancy analysis of community characteristic in relation to major topographic factors QIV-F: 连翘的重要值, Important value of Forsythia suspensa; NB: 群落的物种数量, Number of species in a community; TRP: 坡向指数Transformation of aspect; E: 东经East longitude; N: 北纬Northern latitude; H: 海拔高度Height above sea level |

|
| 图 12 样方与地形因子的冗余分析 Fig. 12 Redundancy analysis of sample plots in relation to major topographic factors |
从图 11可知, 样方的坡向(TRP)、经度(E)、纬度(N)、海拔高度(H)与连翘的重要值(QIV-F)有正相关关系, 其中, 纬度与QIV-F的密切度最高(夹角最小), 但地形因子与群落的物种数量(NB)呈负相关关系, 其中, TRP的影响最大。坡向指数越大, 越向阳、环境越干燥, 物种越少;由图 12可知, TRP、N、H与前2个排序轴均有正相关关系, 其中TRP因子与第一个排序轴是强正相关, 与第二排序轴正相关关系较弱(夹角较大), N因子与2个排序轴有较弱的正相关关系(夹角较大), 而H因子与第二排序轴有较强的正相关关系, 与第一排序轴的关系较弱(夹角较大);经度(E)因子与第二排序轴有强的正相关关系而与第一轴是强负相关。总体上, 样方与不同地形因子的密切度呈TRP > N > H > E的趋势。说明TRP和N地形因子是制约连翘群落分布的关键性位置因子, 其次是海拔高度, 影响力最小的是经度。
3.4 土壤因子与群落数量特征值的梯度分析以样方的物种数量和连翘物种的重要值与土壤因子构成数据矩阵, 做约束性排序分析(RDA), 并进行Monte Carlo置换检验[25]。各土壤因子变量与物种排序轴的相关系数、特征值、累积贡献率及检验结果见(表 2)。第一排序轴解释了变量的88.6%, 与TK、TP、AP和EC相关性较大, 基本上代表了影响连翘生长的主要环境因子;第二排序轴, 解释了总变量的11.4%与AP、AK、TN、TOC、TP、EC的相关性较高。两个排序轴的累积贡献率达到100%, 很好地解释了土壤因子对群落物种分布的影响。分别以物种-土壤因子做图、样方-土壤因子做图(图 13—14)。从Pr值看, TN、TK对连翘物种分布有高度的相关性, Pr值为0.010。其次是土壤总盐分(EC, Pr=0.060)、总磷(TP, Pr=0.259)和总有机质(TOC, Pr=0.338)。
| 排序轴Axis | 轴1 Axis1 | 轴2 Axis2 | r2 | Pr |
| 电导率Electrical conductivity(EC) | -0.894 | -0.447 | 0.084 | 0.060. |
| 总有机质Total organic matter(TOC) | -0.997 | -0.072 | 0.038 | 0.338 |
| 总氮Total potassium(TN) | 0.783 | -0.622 | 0.145 | 0.010 ** |
| 总磷Total phosphorus(TP) | 0.282 | 0.959 | 0.046 | 0.259 |
| 总钾Total potassium(TK) | 0.146 | 0.989 | 0.413 | 0.001 *** |
| 速效氮Available potassium(AN) | -0.620 | -0.785 | 0.003 | 0.891 |
| 速效磷Available phosphorus(AP) | 0.509 | 0.861 | 0.005 | 0.830 |
| 速效钾Available potassium(AK) | -0.975 | -0.221 | 0.013 | 0.720 |
| 特征值Eigenvalue | 91.8 | 19.6 | ||
| 解释量Interpretation/% | 88.6 | 11.4 |

|
| 图 13 连翘群落结构与土壤环境因子的冗余分析 Fig. 13 Redundancy analysis of community characteristic in relation to soil environmental variables EC: 电导率Electrical conductivity; TOC: 总有机质Total organic matter; TN: 总氮Total potassium; TP: 总磷Total phosphorus; TK: 总钾Total potassium; AN: 速效氮Available potassium; AP: 速效磷Available phosphorus; AK: 速效钾Available potassium |

|
| 图 14 样方与土壤环境因子的冗余分析 Fig. 14 Redundancy analysis of sample plots in relation to soil environmental variables |
由图 13可知, TN、AN、AP、AK、TP、AP与连翘的重要值(QIV-F)正相关, 尤其氮的影响更加显著, 而TK、TOC和EC与连翘的多样性呈负相关关系, 尤其是, TK的影响最大。同时可知, TK、AN、AP、AK、TP、AP与样方物种的数量(NB)有正相关关系, 而TN、TOC、EC与其呈现负相关关系。表明研究区域土壤钾是连翘群落物种多样性及其生长的促进因子, 而TOC和EC是连翘群落的2个典型的制约性因子。
在样方与土壤因子的RDA排序图中样方用三角形表示, 实线箭头表示土壤环境因子, 箭头连线的长短表示连翘群落的分布与该环境因子相关性的大小, 箭头连线与排序轴夹角的大小表示该环境因子与排序轴相关性的大小, 夹角小说明关系密切, 箭头所处的象限表示环境因子与排序轴的正、负相关性。每个样方的点投影到某一环境因子的箭头, 投影点的相对位置代表拟合的多度值(即能够被排序模型所解释的部分)。从图 14可知, 土壤TN因子与两个排序轴均是强正相关;TK因子与2个排序轴均是强负相关关系;TP、AN、AP、AK与第一轴是弱负相关(与第一轴的夹角很大), 与第二轴是强正相关;总有机质含量(TOC)和土壤盐离子含量(EC)与第一轴负相关, 与第二轴正相关。从样方分布看, 可分为三组, 第一组与TN、TOC、EC环境因子密切相关, 说明这些土壤因子对样方有明显的影响;第二组与所有土壤因子均有较密切的关系;第三组则与TP、TK、AP、AK、AN等土壤因子的关系较为密切。表明土壤因子对不同样方均有影响, 但影响力度有一定的差异, 认为该结果与土壤因子的碎片化分布有关。
4 讨论 4.1 群落分型的四级主导因子根据连翘群落结构特征及环境因子相关性分析, 我们认为影响该区连翘群落结构的环境因子可分为四个等级。第一级环境因子是海拔高度(H), 引起群落分型的H在1330 m。在低于1330 m的区域, H是连翘群落结构的主导因子, 而在高海拔区域(H≥1330 m), 土壤总钾量(TK)是群落结构分型的第二个界限(TK≥19.07 mg/kg), TK构成了连翘群落结构的二级主导因子;在高海拔(H≥1330 m)和TK < 19.07 mg/kg的环境条件下, 坡向系数(TRP≥0.2275)构成了连翘群落分型的第三个界限, 坡向是连翘群落结构的三级主导因子;在高海拔H≥1330 m、TRP≥0.2275区域, 连翘群落再次以土壤钾含量(TK≥18.47)为主导因子出现了分型, 构成了第四级主导因子。再次以钾分型的原因, 作者认为, 因为土壤中的钾通常有全钾、速效钾和缓释钾三种状态, 而且钾是植物生长的重要肥力之一。速效钾是能够被植物直接吸收的类型, 但也容易发生淋溶与洗脱。在山地环境容易随降雨等自然现象发生沿坡向流失或积累, 导致碎片化随机分布。
总体来看, 我们认为影响该区连翘群落分型的因子有三个, 分为四级主导群落分型。海拔高度是主导该区连翘群落分型的一级环境因子。1330 m的海拔高度是样区连翘群落结构分型的界线。在海拔低于1330 m的样方, 群落类型主要为Ⅰ型群丛(连翘+三裂绣线菊-披针叶苔草群丛), 是该区最典型的群落组成结构(78.3%的样方属于此类型)。在高海拔区域, 影响连翘群落分型的因子是土壤钾含量和坡向。原因分析认为, 海拔和坡向一方面会影响微环境的温度、湿度, 另一方面也会影响土壤母质的生长发育过程, 直接影响着土壤中植物生长所需的营养物质的种类与含量。Kessell等[26]研究认为海拔和地形(地表朝向)、土壤基质、坡度等因素会影响空气和地面温度, 以及群落范围内的湿度、营养物质和其他物质(如污染物)的数量等。潮湿的气团在丘陵和山区流动, 较高的降水量通常落在较高的海拔, 形成地形效应, 进而对植被和群落内能量流动产生影响[27]。
主导群落物种组成的因子主要是坡向、海拔、土壤总氮、有机质和土壤盐分。我们认为在样方分布的丘陵、山地等陆生环境中, 坡向越向阳、海拔越高, 环境中的水分、温度和土壤肥力等条件越差, 对植物的选择性越高。坡向导致的土壤含水量差异性分布是影响植物生长和分布的主要因子, 植物群落组成也因坡向不同而显著变化[28-29]。研究[30-31]认为随着海拔梯度的变化, 环境中的水热因子等相应发生变化, 群落的物种组成会逐步发生改变, 且有利于植物不同特征性状的建立, 从而导致群落中的物种种类组成表现出一定的差异。张玲等[7]对吕梁山、太岳山、中条山连翘种群数量特征的研究表明, 海拔900—1300 m可形成连翘自然群落, 900 m以下或1300 m以上易形成混生群落, 主要与其它乔木、灌木、草本植物混生。
4.2 连翘群落生境的碎片化样方分布区地处太行山南端, 属于丘陵山地地形, 其海拔高度、坡向复杂, 受冲刷、遮阴等自然条件、植被因子和人类活动的影响, 土壤因子的分布呈斑块化, 不同样方之间差异较大。总磷量从0.03%到0.86%, 有效磷的含量在10—80 mg/kg之间, 包括了低磷、中磷和高磷的土壤类型;总钾量在1.0%—2.0%之间, 速效钾的含量在100—300 mg/kg之间;总氮量在0.1%—1.0%之间, 有效氮含量在50—1000 mg/kg之间;有机质含量在2.0%—15.0%之间, 土壤pH在7.0—9.0之间。总体来看, 影响植物生长的三大元素(氮、磷、钾)的分布呈碎片化状态, 不同样方之间差异较大。碎片化的环境因子, 必然会导致植物群落结构的变化。
冗余分析可知, 地形因子对连翘重要值有正向影响, 而与物种负相关;从群落结构与样方的对应性分析可知:地形因子, 尤其是坡向、海拔高度是影响群落分布的典型因子。从土壤因子与连翘重要值、群落物种的对应性分析可知:土壤中的钾、有机质和盐总量是连翘重要值的制约性因子, 而土壤氮、磷是连翘重要值的促进因子, 尤其是土壤钾、磷和植物可吸收性有效磷、有效氮、速效钾等因子与群落物种的多样性正相关, 且土壤总氮、磷、钾、速效钾是影响群落结构的主要土壤因子。冗余性分析结果与土壤因子的分析结果一致, 该区连翘群落生境的碎片化, 导致群落结构的差异较大。该结果与相关研究具有一致性, 如研究[32-33]认为在群落尺度上, 微生境和土壤养分是植被格局的主导因子, 在地貌与植被格局的关系中隐含着复杂的生态系统斑块, 特别是在山地环境中, 局部生境在温度、湿度, 尤其是在土壤的发育过程及其化学组份等因子之间存在较大的差异。由于海拔和地形(地表朝向)对太阳能和水体的影响, 从而导致地形和海拔高度等因子直接影响到群落的物种组成、结构和功能[34-35]。
5 结论本文以太行山南段陵川境内的连翘群落为研究对象, 得出如下结论:
(1) 太行山南段陵川境内的连翘群落有5种类型:Ⅰ型: 连翘+三裂绣线菊+披针叶苔草群丛;Ⅱ型: 连翘+白刺花-披针叶苔草+铁杆蒿群丛;Ⅲ型: 辽东栎-连翘+三裂绣线菊-披针叶苔草+白头翁群丛;Ⅳ型: 油松-连翘-披针叶苔草群丛;Ⅴ型: 连翘+黄刺玫-披针叶苔草群丛。
(2) 海拔高度、土壤钾、坡向是影响该区连翘分布的3个主导性环境因子。3个因子相互作用从四个层级引起连翘群落的分型。其中, 海拔是影响连翘群落分布格局的第一级环境因素, 以海拔1330 m为分界线。在海拔大于1330 m的区域, 土壤全钾、坡向对群落格局有不同的主导性作用。
(3) 土壤全钾、全磷、全氮、速效钾、有效氮、有效磷, 有机质等与植被生长关系密切的土壤因子, 在该区的分布碎片化, 导致了不同样方群落结构的差异。
建议在此山地环境进行连翘基地化种植过程中, 应选择在海拔1330 m以下、坡向系数小于0.2775的区域, 并适当补充土壤养分(氮、磷、钾)、调节土壤pH值(中性/弱碱性)和适当改造坡向, 调节生境的干热度。
| [1] |
山西森林编委会. 山西森林. 北京: 中国林业出版社, 1984: 27-27.
|
| [2] |
Zhang S, Shao S Y, Song X Y, Xia C Y, Yang Y A, Zhang P C, Chen N H. Protective effects of Forsythia suspense extract with antioxidant and anti-inflammatory properties in a model of rotenone induced neurotoxicity. NeuroToxicology, 2016, 52: 72-83. DOI:10.1016/j.neuro.2015.09.009 |
| [3] |
Yuan J F, Liu X Q, Yang G X, Cui X Q. Forsythia suspense leaves, a plant: extraction, purification and antioxidant activity of main active compounds. European Food Research and Technology, 2014, 238(4): 527-533. DOI:10.1007/s00217-014-2179-y |
| [4] |
王建华, 任士福, 史宝胜, 刘炳响, 周玉丽. 遮荫对连翘光合特性和叶绿素荧光参数的影响. 生态学报, 2011, 31(7): 1811-1817. |
| [5] |
郎莹, 汪明. 春、夏季土壤水分对连翘光合作用的影响. 生态学报, 2015, 35(9): 3043-3051. |
| [6] |
胡贝娟, 张钦弟, 张玲, 毕润成. 山西太岳山连翘群落优势种种间关系. 生态学杂志, 2013, 32(4): 845-851. |
| [7] |
张玲. 连翘的数量生态学研究[D]. 临汾: 山西师范大学, 2012.
|
| [8] |
韩虹. 连翘群落数量生态研究[D]. 太原: 山西大学, 2008.
|
| [9] |
李宝堂. 山西历山自然保护区连翘群落及物种多样性. 东北林业大学学报, 2011, 39(8): 36-39. DOI:10.3969/j.issn.1000-5382.2011.08.012 |
| [10] |
茹文明, 张桂萍, 张金屯. 太行山南段野生植物资源研究. 山西大学学报: 自然科学版, 2007, 30(3): 411-414. DOI:10.3969/j.issn.0253-2395.2007.03.026 |
| [11] |
茹文明, 张桂萍, 张金屯, 张峰. 太行山南段森林群落物种多样性研究. 西北植物学报, 2006, 26(5): 1036-1042. DOI:10.3321/j.issn:1000-4025.2006.05.026 |
| [12] |
焦海华, 张淑珍, 景旭东, 张通, 白志辉. 油田区多环芳烃污染盐碱土壤活性微生物群落结构解析. 生态学报, 2016, 36(21): 6994-7005. |
| [13] |
鲍士旦. 土壤农化分析 ((第三版)). 北京: 中国农业出版社, 2012.
|
| [14] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000.
|
| [15] |
Yamakura T, Kanzaki M, Itoh A, Ohkubo T, Ogino K, Chai E O K, Lee H S, Ashton P S. Topography of a large-scale research plot established within a tropical rain forest at Lambir, Sarawak. Tropics, 1995, 5(1): 41-56. |
| [16] |
张荣, 刘彤. 古尔班通古特沙漠南部植物多样性及群落分类. 生态学报, 2012, 32(19): 6056-6066. |
| [17] |
陈云, 王海亮, 韩军旺, 韦博良, 贾宏汝, 叶永忠, 袁志良. 小秦岭森林群落数量分类、排序及多样性垂直格局. 生态学报, 2014, 34(8): 2068-2075. |
| [18] |
孙儒泳, 李博, 诸葛阳, 尚玉昌. 普通生态学. 北京: 高等教育出版社, 1993.
|
| [19] |
郝占庆, 于德永, 杨晓明, 丁之慧. 长白山北坡植物群落α多样性及其随海拔梯度的变化. 应用生态学报, 2002, 13(7): 785-789. DOI:10.3321/j.issn:1001-9332.2002.07.005 |
| [20] |
Zhang J T, Xu B, Li M. Vegetation patterns and species diversity along elevational and disturbance gradients in the Baihua Mountain reserve, Beijing, China. Mountain Research and Development, 2013, 33(2): 170-178. DOI:10.1659/MRD-JOURNAL-D-11-00042.1 |
| [21] |
Valencia R, Foster R B, Villa G, Condit R, Svenning J C, Hernández C, Romoleroux K, Losos E, Magård E, Balslev H. Tree species distributions and local habitat variation in the Amazon: large forest plot in eastern Ecuador. Journal of Ecology, 2004, 92(2): 214-229. DOI:10.1111/j.0022-0477.2004.00876.x |
| [22] |
赖江山, 米湘成, 任海保, 马克平. 基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例. 植物生态学报, 2010, 34(7): 761-769. |
| [23] |
张文静, 张钦弟, 王晶, 冯飞, 毕润成. 多元回归树与双向指示种分析在群落分类中的应用比较. 植物生态学报, 2015, 39(6): 586-592. |
| [24] |
宋永昌. 植被生态学. 上海: 华东师范大学出版社, 2001.
|
| [25] |
包小婷, 丁陆彬, 姚帅臣, 王景升, 石培礼, 王彤, 李超, 刘文婧. 拉萨河流域植物群落的数量分类与排序. 生态学报, 2019, 39(3): 779-786. |
| [26] |
Kessell S R. Gradient Modeling. New York: Springer, 1979.
|
| [27] |
文志, 赵赫, 刘磊, 王丽娟, 欧阳志云, 郑华, 李彦旻. 基于土地利用变化的热带植物群落功能性状与土壤质量的关系. 生态学报, 2019, 39(1): 371-380. |
| [28] |
李新娥. 亚高寒草甸阳坡-阴坡梯度上植物功能性状及群落构建机制研究[D]. 兰州: 兰州大学, 2011.
|
| [29] |
刘丽艳, 张峰, 秦浩. 山西太行山森林群落物种多样性研究. 山西大学学报: 自然科学版, 2019, 42(3): 673-681. |
| [30] |
Wehenkel C, Corral-Rivas J J, Gadow K V. Quantifying differences between ecosystems with particular reference to selection forests in Durango/Mexico. Forest Ecology and Management, 2014, 316: 117-124. DOI:10.1016/j.foreco.2013.05.056 |
| [31] |
李国旗, 邵文山, 赵盼盼, 靳长青, 陈彦云. 荒漠草原区4种植物群落土壤种子库特征及其土壤理化性质. 生态学报, 2019, 39(17): 6282-6292. |
| [32] |
马旭东, 张苏峻, 苏志尧, 区余瑞, 刘刚. 车八岭山地常绿阔叶林群落结构特征与微地形条件的关系. 生态学报, 2010, 30(19): 5151-5160. |
| [33] |
Swanson F J, Kratz T K, Caine N, Woodmansee R G. Landform effects on ecosystem patterns and processes: geomorphic features of the earth's surface regulate the distribution of organisms and processes. Bioscience, 1988, 38(2): 92-98. DOI:10.2307/1310614 |
| [34] |
田中平, 庄丽, 李建贵. 伊犁河谷北坡垂直分布格局及其与环境的关系——一种特殊的双峰分布格局. 生态学报, 2012, 32(4): 1151-1162. |
| [35] |
牛钰杰, 周建伟, 杨思维, 王贵珍, 刘丽, 杜国祯, 花立民. 基于地形因素的高寒草甸土壤温湿度和物种多样性与初级生产力关系研究. 生态学报, 2007, 37(24): 8314-8325. |