 2021, Vol. 41
2021, Vol. 41文章信息
- 彭钟通, 郭明明, 张远东, 顾峰雪, 邵辉, 刘世荣
- PENG Zhongtong, GUO Mingming, ZHANG Yuandong, GU Fengxue, SHAO Hui, LIU Shirong
- 升温突变对川西道孚林线川西云杉和鳞皮冷杉生长的影响
- Effects of abrupt warming on Picea likiangensis var. balfouriana and Abies squamata growth at tree line in Dafu, Sichuan, China
- 生态学报. 2021, 41(20): 8202-8211
- Acta Ecologica Sinica. 2021, 41(20): 8202-8211
- http://dx.doi.org/10.5846/stxb202012113161
-
文章历史
- 收稿日期: 2020-12-11
- 修订日期: 2021-08-16





2. 河北农业大学, 保定 071000;
3. 中国农业科学院农业环境与可持续发展研究所, 农业部旱作节水农业重点实验室, 北京 100081
2. Hebei Agricultural University, Baoding 071000, China;
3. Key Laboratory of Dryland Agriculture, Ministry of Agriculture, Institute of Environment and Sustainable Development in Agriculture, Chinese Academy of Agricultural Sciences, Beijing 100081, China
高山林线是对气候变化非常敏感的生态过渡带, 也是理想的全球气候变化监测器[1]。高山林线作为陆地森林生态系统分布的最高界限, 受到人为干扰相对较少, 成为研究植被与气候变化关系的重要场所[2]。温度是高山林线分布的重要限制因子, 特别是低温对高山林线植物光合特性、养分特征等有着重要影响[3]。通常来说, 寒区树木生长与温度表现为正相关, 气候变暖对寒区树木生长是正效应[4], 但近些年发现温度升高可能会降低一些环北极地区树木径向生长的温度敏感性, 甚至会存在正效应转为负效应[5], 或是同一地点内的树木在数十年内从生长趋势相同到生长趋势分化[6], 这在树轮研究中被称为“响应分歧现象”[6]。如Lloyd和Fastie[7]研究发现, 在阿拉斯加地区, 除最为湿润的阿拉斯加山脉以外的其它区域的树木在20世纪后半期生长均普遍下降, 而阿拉斯加北方森林对于升温整体表现为负响应。而在北美洲西北部的白云杉生长调查研究中发现, 白云杉的生长格局在1950s后分化为两类, 一类对生长季温度升高呈现正效应, 另一类则表现为负效应, 但与生长季降水表现为一定的正相关[8]。高山林线也是低温限制的产物, 在欧洲阿尔卑斯高山区, Büntgen等[9]研究表明:20世纪以来挪威云杉(Picea abies)树轮温度敏感性下降, 而降水敏感性则增加[10]。而在近期国内川西贡嘎山区的研究发现, 处在高山林线上的冷杉树轮宽度与温度依然表现为一定的正相关[11];而在川西卧龙地区, 冷杉树轮-气候响应具有时间变异性, 树轮指数与温度序列在1956—1976年时段吻合程度较高, 但树轮指数在1977—2008年时段则明显偏低, 表现出“响应分歧现象”[12]。“响应分歧现象”挑战了以往对于气候变化情景下高山林区森林生长和生产力动态的预测。
另一方面, 树木生长与气候限制因子间的关系一般在树木年轮气候学的研究中一般被假定认为是长期稳定的[13]。树木生长对外界环境变化的反应是一个生理生态的过程, 会受到外界环境和自身遗传因素的影响[14]。而树木生长的弹性则是在生态系统中抵抗干扰并维持和恢复自身结构和功能的能力, 即包括抵抗力和恢复力[15]。研究表明, 树木生长在受到极端事件影响时, 只要不超过弹性阈值, 树木就能积极抵抗不利干扰的影响并恢复到干扰前的生长状况[16]。由于生长敏感性、气孔特征和生理特征的差异, 不同树种对极端气候的恢复能力存在差异[17], 而外界环境的变化(如干旱时间)对树木恢复力也有重要影响[18]。高海拔树木生长的环境相对较为恶劣, 但高海拔树木的生长尤其是林线上树木生长受环境变化的影响还尚未全面了解[19], 所以林线树木对环境的响应尤其是弹性变化的研究也显得十分重要。因此, “响应分歧现象”存在会对区域树轮气候重建的准确性具有一定的影响。但青藏高原东缘高山林线的响应分歧现象发生规律尚不明朗, 亟需更多的研究来探讨该区主要树种对近期升温的响应。
青藏高原东缘高山森林广布, 主要树种包括云杉、冷杉等, 是我国第二大林区和重要的生物碳库[20], 是我国树轮气候重建的重点区域, 同时也是全球25个生物多样性优先保护热点地区之一[21]。青藏高原作为全球气候变化最为敏感的地区之一, 全球变暖背景下, 青藏高原地区气温也普遍升高[22]。吴成启等[23]对青藏高原1961—2010年气候资料分析表明, 青藏高原年均气温以0.0228℃/a的速率递增, 1996年左右为青藏高原年均气温开始突变的年份。郭明明等[24]对川西马尔康主要针叶树种的研究表明, 1995年变暖突变后树木年轮指数呈下降趋势, 表现出“响应分歧现象”, 但该研究仅针对于中高海拔岷江冷杉(Abies faxoniana)和低海拔岷江柏(Cupressus chengiana), 缺乏对于高山林线的研究。
在青藏高原东缘, 随着降水量由东南向西北逐渐减少, 以喜阴湿的冷杉属植物为优势种的森林逐渐过渡为更耐干冷的云杉林。川西道孚至理塘一线, 处于该过渡带上, 云冷杉并存;在该线东南区域, 冷杉林占优势;该线以西北则主要是云杉林[25]。在这一过渡带上探究林线云冷杉生长对升温突变的响应及树种间差异, 对揭示响应分歧现象的发生规律具有重要意义。本研究选择位于青藏高原东缘半湿润区的川西道孚县为研究区, 该区高山林线同时有鳞皮冷杉(Abies squamata)和川西云杉(Picea likiangensis var. balfouriana)分布, 常在林线附近形成二者的混交林。升温突变后, 该区林线云杉和冷杉是否出现响应分歧现象?树种间有何差别?本文拟使用树轮生态学方法, 对这一问题进行深入分析和探讨。
1 材料与方法 1.1 研究区域概况道孚县位于四川省甘孜州东北部, 地理坐标东经100°32′—101°44′、北纬32°21′—30°32′, 地处青藏高原东缘鲜水河断裂带, 海拔2800—5300m, 属温带大陆性季风气候。根据四川道孚气象站(E101°7′, N30°59′, 海拔2957m)数据, 1958—2015年间, 年均气温8.0℃, 年降水量约604.2mm, 无霜期为113d左右, 昼夜温差较大, 冬季漫长且寒冷干燥。该区高山森林广布, 主要树种为川西云杉和鳞皮冷杉, 分布在海拔2800—4100m之间, 高山气候林线主要分布在4000—4100m, 林线之上为高山灌丛或高山草甸[26]。
1.2 树轮材料2016年夏季在道孚县木茹乡林线(4021—4042m)上选择川西云杉和鳞皮冷杉混交林分, 基于国际树轮数据库标准, 选择林线上生长良好的冷杉和云杉个体, 在树高1.3m处用生长锥获取样芯, 平行于坡向钻取到树木髓芯, 对林线上的每株树木都采集1个样芯, 须采集云杉和冷杉各30颗以上的树芯, 并将样品放入塑料管内同时进行编号。因采样点位于海拔4000m以上的林线, 几乎无人类活动影响, 故而所采树芯适合做树木年轮气候学相关分析[27]。将收集好的样芯带回实验室后, 应剔除断裂或心腐等不符合条件的部分样芯, 并按照Stokes和Smile[28]阐述的标准方法对样品进行预处理。将样品带回实验室进行晾干, 而后将树芯用白乳胶粘着、固定于木质凹槽内, 用砂纸打磨处理至出现清晰年轮[29]。通过骨架图法并使用双筒显微镜进行目测交叉定年。而后使用LINTAB 5树木年轮仪(精度为0.01mm)对树木年轮宽度进行测量, 交叉定年和年轮宽度测量的结果则用COFECHA程序[30]来检验, 同时手动消除交叉定年和年轮宽度测量过程中出现的误差。为保留更多的低频变化趋势, 降低因年龄生长速率不同而带来的影响, 使用ARSTAN程序采用负指数或线性函数对经过交叉定年的年轮序列进行去趋势, 而后用双权重平均法对数据进行标准化, 最终建立并获得标准、残差和自回归共三个年表[31]。对比获得的年表统计特征值, 根据平均敏感度指标, 本研究选用优于标准年表的差值年表进行年轮-气候的相关分析。
1.3 生长弹性的计算一般来说, 树木生长弹性展现了树木对干扰的抵抗能力和恢复到原来状况的能力, 因而可以用抵抗力稳定性和恢复力稳定性来表示树木生长弹性[32]。抵抗力(Resistance, Rt)和恢复力(Recovery, Rc)的计算公式如下[33]:
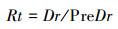
|
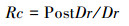
|
其中Dr表示当年的年轮指数, PreDr和PostDr分别表示前后4年的平均年轮指数。
1.4 气象资料本研究气象数据使用距离采样点最近(约35km距离)的道孚气象站(国家标准气象台站, 30°59′N, 101°07′E, 海拔2957.2m;国家气象科学数据中心提供, https://data.cma.cn/), 包括1958—2015年期间的年均温、年降水量数据和各月平均气温、月均最高温、月均最低温和月降水量数据。树木生长不仅受当季气候影响, 还受上一季气候影响, 因此, 本研究采用的前一年9月至当年9月作为月气象指标的分析区间。使用IBM SPSS Statistics 22数据分析软件对年表与气候要素之间进行相关分析计算。
研究发现, 青藏高原气温在20世纪80年代中期已经发生了变暖, 同时发现90年代中期青藏高原气温开始升温突变[34-37], 吴成启等[23]发现1996年左右为青藏高原年均气温开始突变的年份, 而丁一汇等[38]则发现青藏高原地表气温增温突变发生在1995年左右。本研究所在的道孚地区90年代中期以后年均温升温也开始加快(图 1), 1995—2015年的均温极显著高于1958—1994年的均温(P < 0.001, 图 2)。因此, 本研究选定1995年为升温分界点, 分为1958—1994和1995—2015年两个时间段对树轮指数与气候要素进行相关分析。

|
| 图 1 道孚地区升温突变前后年均温与降水量的变化 Fig. 1 The trends of annual mean temperature and annual precipitation before and after warming abrupt change in Daofu |

|
| 图 2 道孚地区升温突变前后年均温与降水量的变化对比 Fig. 2 Comparison of the trends of annual mean temperature and annual precipitation before and after warming abrupt change in Daofu |
从表 1可知:林线川西云杉的最大树龄长于鳞皮冷杉, 平均敏感度高于鳞皮冷杉, 说明川西云杉各轮宽间的年际变化更明显。但川西云杉信噪比、样本代表性和第一主成分方差解释量均低于鳞皮冷杉, 说明鳞皮冷杉年表含有较多的气候信息量, 年表质量相对较好。其中, 川西云杉和鳞皮冷杉样本代表性均大于0.85, 表明树芯的代表性强, 符合理论年表, 可以进行年轮气候分析[24, 39]。
| 特征指标 Statistic characteristics |
川西云杉 P. likiangensis var. balfouriana |
鳞皮冷杉 A. squamata |
| 序列长度Time span | 1525—2015 | 1693—2015 |
| 平均敏感度Mean sensitivity | 0.13 | 0.12 |
| 标准差Standard deviation | 0.12 | 0.11 |
| 平均相关系数Mean correlation coefficient | 0.28 | 0.31 |
| 信噪比Signal-to-noise ratio | 6.68 | 10.41 |
| 样本代表性Expressed population signal | 0.87 | 0.91 |
| 第一主成分方差解释量Variance in first eigenvector /% | 32.86 | 35.35 |
年尺度上, 升温突变前, 川西云杉和鳞皮冷杉年轮指数表现出缓慢上升趋势;升温突变后, 川西云杉年轮指数趋势为负, 鳞皮冷杉的趋势为正, 但均不显著(图 3), 没有发生明显的响应分歧现象。

|
| 图 3 1995年前后温度及年轮指数变化趋势 Fig. 3 The change trend of temperature and tree ring index before and after 1995 川西云杉年轮指数变化趋势:y=0.0002x+0.5266 P>0.05 (1958—1994); y=-0.0002x+1.4043 P>0.05 (1995—2015); 鳞皮冷杉年轮指数变化趋势:y=0.0002x+0.5266 P>0.05(1958—1994);y=0.0003x+0.4024 P>0.05(1995—2015) |
相比于1995年升温突变以前, 升温突变后的川西云杉和鳞皮冷杉的抵抗力和恢复力随时间变化的下降速度加快, 说明升温降低了林线树木弹性, 长期不利于树木生长。其中升温突变以后云杉的抵抗力下降速率快于冷杉, 而冷杉的恢复力下降速率则快于云杉。但云杉和冷杉的生长弹性没有响应分歧现象(图 4)。

|
| 图 4 1995年前后抵抗力与恢复力变化趋势 Fig. 4 The change trend of tree resistance and recovery before and after 1995 川西云杉抵抗力变化趋势: y=-0.0004x+1.8918 P>0.05 (1962—1994), y=-0.0029x+6.819 P>0.05 (1995—2015); 鳞皮冷杉抵抗力变化趋势: y=0.0006x-0.2114 P>0.05 (1962—1994), y=-0.0019x+4.8506 P>0.05 (1995—2015);川西云杉恢复力变化趋势: y=-0.0006x+2.1888 P>0.05 (1958—1994), y=-0.0056x+12.1681 P>0.05 (1995—2011);鳞皮冷杉恢复力变化趋势: y=-0.0008x+2.5771 P>0.05 (1958—1994), y=-0.0075x+15.951 P>0.05 (1995—2011) |
月尺度上, 升温突变前, 川西云杉年轮指数与前一年11月的月均最高温、5—7月月均温、6月月均最高温、7月月均最低温呈显著正相关(表 2), 说明夏季温度是影响云杉生长的主要因素。升温突变后, 川西云杉年轮指数与2月降水量呈显著正相关, 与前一年11月均温显著负相关, 与7月月均最低温呈显著正相关(表 2), 7月温度仍是决定云杉生长的重要因素。
升温突变前, 鳞皮冷杉年轮指数与前一年11月份均温、11月份均最高温呈极显著正相关, 与前一年12月份均温呈显著正相关(表 3), 说明冬季温度是影响冷杉生长的主要因素。升温突变后, 年轮指数只与5月份均温、5月份均最低温呈显著负相关, 同时与5月降水的正相关系数达到最大值(表 3), 表明其在升温突变后受到春季干旱胁迫的影响。
| 月份 Month |
1958—1994 | 1995—2015 | |||||||
| 月均最低温 Tmin |
月均最高温 Tmax |
月均温 Tmean |
月降水量 P |
月均最低温 Tmin |
月均最高温 Tmax |
月均温 Tmean |
月降水量 P |
||
| -9 | 0.007 | -0.201 | -0.130 | 0.261 | -0.094 | -0.031 | -0.082 | 0.042 | |
| -10 | -0.138 | -0.283 | -0.255 | -0.148 | -0.183 | -0.260 | -0.279 | -0.132 | |
| -11 | 0.079 | 0.379* | 0.299 | -0.147 | -0.323 | -0.395 | -0.540* | 0.117 | |
| -12 | 0.149 | 0.243 | 0.289 | -0.126 | 0.142 | -0.072 | 0.067 | -0.047 | |
| 1 | 0.186 | 0.194 | 0.271 | -0.253 | 0.349 | 0.160 | 0.308 | -0.009 | |
| 2 | -0.093 | 0.053 | 0.015 | -0.014 | 0.387 | 0.112 | 0.192 | 0.454* | |
| 3 | -0.043 | 0.081 | 0.012 | -0.036 | -0.064 | -0.090 | -0.082 | -0.089 | |
| 4 | -0.104 | -0.004 | -0.027 | 0.018 | -0.027 | -0.124 | -0.114 | -0.157 | |
| 5 | 0.186 | 0.311 | 0.329* | -0.288 | -0.388 | 0.051 | -0.105 | 0.035 | |
| 6 | 0.192 | 0.354* | 0.325* | -0.021 | 0.146 | 0.255 | 0.237 | -0.362 | |
| 7 | 0.357* | 0.163 | 0.336* | 0.027 | 0.433* | 0.346 | 0.430 | 0.260 | |
| 8 | 0.087 | 0.032 | 0.061 | -0.181 | 0.001 | 0.341 | 0.279 | 0.152 | |
| 9 | 0.076 | 0.097 | 0.100 | 0.202 | -0.178 | 0.342 | 0.115 | -0.328 | |
| Tmin: Monthly average minimum temperature; Tmax: Monthly average maximum temperature; Tmean: Monthly average temperature; P: Monthly precipitation; * 代表显著(P < 0.05); 月份列中的“-”代表上一年的意思 | |||||||||
| 月份 Month |
1958—1994 | 1995—2015 | |||||||
| 月均最低温 Tmin |
月均最高温 Tmax |
月均温 Tmean |
月降水量 P |
月均最低温 Tmin |
月均最高温 Tmax |
月均温 Tmean |
月降水量 P |
||
| -9 | 0.049 | -0.077 | -0.061 | 0.296 | -0.128 | -0.131 | -0.181 | 0.159 | |
| -10 | 0.007 | -0.144 | -0.109 | -0.010 | 0.188 | 0.106 | 0.185 | -0.064 | |
| -11 | 0.233 | 0.447** | 0.457** | -0.131 | -0.197 | -0.036 | -0.214 | -0.120 | |
| -12 | 0.239 | 0.282 | 0.372* | -0.256 | -0.013 | -0.162 | -0.055 | -0.152 | |
| 1 | 0.094 | 0.072 | 0.156 | -0.086 | 0.054 | 0.113 | 0.072 | -0.147 | |
| 2 | -0.098 | -0.163 | -0.121 | -0.099 | 0.331 | 0.361 | 0.342 | 0.107 | |
| 3 | 0.021 | -0.197 | -0.160 | 0.028 | 0.138 | -0.210 | -0.134 | 0.098 | |
| 4 | -0.068 | -0.087 | -0.049 | -0.113 | 0.194 | -0.221 | -0.165 | -0.134 | |
| 5 | -0.007 | 0.043 | 0.070 | -0.108 | -0.504* | -0.382 | -0.493* | 0.357 | |
| 6 | 0.255 | 0.312 | 0.317 | 0.071 | 0.198 | 0.272 | 0.308 | -0.246 | |
| 7 | 0.151 | -0.006 | 0.118 | 0.176 | 0.262 | 0.403 | 0.418 | -0.176 | |
| 8 | 0.141 | 0.050 | 0.101 | -0.161 | -0.186 | 0.184 | 0.005 | 0.260 | |
| 9 | 0.142 | 0.187 | 0.229 | 0.174 | ]0.004 | 0.254 | 0.140 | -0.157 | |
| * 代表显著(P < 0.05), **代表极显著(P < 0.01) | |||||||||
本研究结果表明, 道孚林线川西云杉和鳞皮冷杉均无明显的响应分歧现象, 这与郭明明等[24]研究川西马尔康地区树木在升温突变后表现出明显的“响应分歧现象”不同, 也与李宗善等[12]研究川西卧龙地区岷江冷杉在气候变化后与温度序列有明显差异不同, 但与Shi等[40]对整个青藏高原南部的研究结果相同。林线上树木生长与迁移的变化与生态系统碳收支有关[41], 而树种结构的变化是环境变换下物种间的相互作用和促进与竞争的复杂平衡[42]。因此, 云杉和冷杉无明显的响应分歧现象一定程度体现了林线上树种结构对气候响应的动态平衡[43]。
一般来说, 寒冷地区的树木生长与升温呈现为正相关[44]。这是因为当温度在阈值范围内时, 温度升高有利于打破植物的休眠状态, 提高光合作用和增加碳水化合物的积累, 促进木质部的生长, 从而有利于树木的径向生长[45]。但随着温度达到或超过地区林线树木的生长阈值, 较高的气温会导致该地区林线树木呼吸消耗增加, 碳水化合物积累减少, 从而进一步导致年轮变窄[46], 同时也更易发生干旱胁迫使得树木生长下降的情况[47-48]。而申静霞等[49]也发现随着土壤温度的下降, 川西云杉幼苗各器官可溶性糖、淀粉和NSC浓度并未降低, 反而使其呈现出升高或不变的趋势。因而本文中云杉和冷杉生长下降可能为林线树木生长的限制因子存在区域差异, 道孚林线云杉和冷杉的生长仍主要受温度限制[50]。
在本研究中, 升温突变后, 林线上的川西云杉和鳞皮冷杉的抵抗力和恢复力下降的趋势加快, 云杉和冷杉整体的弹性下降, 且无响应分歧现象。高海拔地区的植物经受着干旱、低温、强紫外辐射等多种逆境因子影响, 而植物则能通过调节自身的生理状况来加强抵抗或恢复能力, 并以此来适应逆境[51]。一般认为, 低温是限制高山林线分布的限制因子[3], 但温度升高并不一定促进高山植物生长, 也有可能会抑制其生长[52]。同时, 在生态系统中抵抗力和恢复力的变化一般呈现相反的关系, 即抵抗力增加则恢复力降低, 反之亦然[32]。因而, 弹性下降可能是温度升高加快了树木的蒸腾速率, 同时也影响了高寒森林的光合作用、碳平衡以及水分与养分的利用情况, 使得树木内部调节机制发生变化[51]。另一方面, 不断升温可能加剧的干旱胁迫, 或产生更多的极端事件[53], 也可能使得树木生长弹性下降。并且不同树种对干扰的响应也存在差别, 如Fang等[17]发现刺柏对干旱的响应时间和对干旱前状态的恢复时间均短于冷杉。而在本研究中, 云杉的抵抗力下降速率快于冷杉, 而冷杉的恢复力下降速率则快于云杉。因而, 持续升温可能使得林线上云杉和冷杉生长的抵抗力超过了阈值, 使得稳定性产生了变化[54], 即云杉和冷杉的生长弹性都下降。而今, 有研究者认为在不久的将来, 由于演替动态和气候变暖可能有利于被子植物而不利于裸子植物[55]。在不同森林类型中, 量化森林弹性对气候变化的响应依然是一大片知识空白区, 也是将来的一个研究挑战的方向[56]。
道孚林线年轮指数与月气候要素的相关性存在着树种间差别。升温突变前, 川西云杉生长主要受夏季温度控制, 而鳞皮冷杉生长主要受冬季温度控制;1995年升温突变后, 川西云杉生长仍受夏季温度限制;而鳞皮冷杉则受到春季干旱胁迫的影响。这可能是气候变暖导致生长季提前和延长, 使得鳞皮冷杉在生长季早期受到低温冻害和干旱事件增加[57]。
在以往的研究中, 人们认为干旱胁迫会造成响应分歧现象。如Driscoll等[58]推断由于干旱胁迫, 同时位点生境差异使得阿拉斯加南部白云杉树轮发生响应分歧现象。而Lloyd和Fastie[59]对阿拉斯加树轮产生响应分歧现象也有着相同的解释, 他们发现干暖位点树轮生长速率下降更为普遍和显著, 并由此推断升温引起的干旱胁迫可能是造成响应分歧现象的重要原因。同时在北美西部White Mountains地区刺果松(Pinus longaeva)响应分歧现象研究中, Salzer等[60]发现响应分歧现象与树木生理形态特征、去趋势方法和大气污染等都没有关系, 因此, 20世纪后半期升温最有可能引起该地区刺果松的响应分歧现象。而在Jiao等[61]研究发现阿拉泰山和天山的西伯利亚落叶松受到不同月气候要素的影响, 这主要是由于区域温度差异导致西伯利亚落叶松生长存在响应分歧现象。本研究中鳞皮冷杉尚未出现明显的响应分歧现象, 但在升温突变后已受到春季干旱胁迫的影响, 未来如果该区随着升温出现干暖化现象, 林线鳞皮冷杉的生长则可能受到进一步抑制并出现明显的响应分歧现象。
4 结论本文以1995年作为温度突变点, 在年尺度上, 升温突变前后川西云杉和鳞皮冷杉年轮指数和生长弹性的时间变化趋势均不显著, 没有出现显著的响应分歧现象, 一定程度反应了林线群落结构的稳定性。在树木弹性的研究中, 升温突变后, 云杉和冷杉的抵抗力和恢复力的下降速度加快, 表明现阶段温度升高可能已成为道孚地区林线上云杉和冷杉生长的不利条件。而在月尺度上, 升温突变前林线云杉和冷杉生长分别受夏季和冬季温度控制;升温突变后, 川西云杉生长依然受夏季温度控制, 而鳞皮冷杉生长则受到春季干旱胁迫的影响。因而, 温度变化会对川西道孚地区林线植物的生长和结构稳定性产生影响, 但在后期研究中还需加入更多因素进行综合探讨。
致谢: 本文部分野外取样工作由曲昌蕾完成。| [1] |
Körner C. A re-assessment of high elevation treeline positions and their explanation. Oecologia, 1998, 115(4): 445-459. DOI:10.1007/s004420050540 |
| [2] |
戴君虎, 崔海亭. 国内外高山林线研究综述. 地理科学, 1999, 19(3): 243-249. DOI:10.3969/j.issn.1000-0690.1999.03.010 |
| [3] |
丛毓, 贺红士, 谷晓楠, 徐文茹, 刘凯, 宗盛伟, 杜海波. 高山林线形成机理研究进展. 应用生态学报, 2016, 27(9): 3035-3041. |
| [4] |
赵小祥, 王根绪, 杨凯, 冉飞, 杨阳, 杨燕. 高山植物种子萌发、幼苗存活和定居对气候变化的响应. 草业科学, 2020, 37(2): 213-225. |
| [5] |
Wilmking M, Myers-Smith I. Changing climate sensitivity of black spruce (Picea mariana Mill.) in a peatland-forest landscape in Interior Alaska. Dendrochronologia, 2008, 25(3): 167-175. DOI:10.1016/j.dendro.2007.04.003 |
| [6] |
Wilmking M, Juday G P. Longitudinal variation of radial growth at Alaska's northern treeline-recent changes and possible scenarios for the 21st century. Global and Planetary Change, 2005, 47(2/4): 282-300. |
| [7] |
Lloyd A H, Fastie C L. Spatial and Temporal Variability in the Growth and Climate Response of Treeline Trees in Alaska. Climatic Change, 2002, 52: 481-509. DOI:10.1023/A:1014278819094 |
| [8] |
Porter T J, Pisaric M F J. Temperature-growth divergence in white spruce forests of Old Crow Flats, Yukon Territory, and adjacent regions of northwestern North America. Global Change Biology, 2011, 17(11): 3418-3430. DOI:10.1111/j.1365-2486.2011.02507.x |
| [9] |
Büntgen U, Frank D C, Schmidhalter M, Neuwirth B, Seifert M, Esper J. Growth/climate response shift in a long subalpine spruce chronology. Trees, 2006, 20(1): 99-110. DOI:10.1007/s00468-005-0017-3 |
| [10] |
Büntgen U, Frank D, Wilson R, Carrer M, Urbinati C, Esper J. Testing for tree-ring divergence in the European Alps. Global Change Biology, 2008, 14(10): 2443-2453. DOI:10.1111/j.1365-2486.2008.01640.x |
| [11] |
段建平, 王丽丽, 徐岩, 孙毓, 陈津. 贡嘎山东坡不同海拔高度树轮宽度对气候变化的响应. 地理研究, 2010, 29(11): 1940-1949. |
| [12] |
李宗善, 刘国华, 傅伯杰, 张齐兵, 胡婵娟, 罗淑政. 川西卧龙国家级自然保护区树木生长对气候响应的时间稳定性评估. 植物生态学报, 2010, 34(9): 1045-1057. DOI:10.3773/j.issn.1005-264x.2010.09.005 |
| [13] |
Fritts H C. Tree Rings and Climate. London: Academic Press, 1976.
|
| [14] |
Willis K J, Jeffers E S, Tovar C. What makes a terrestrial ecosystem resilient?. Science, 2018, 359(6379): 988-989. DOI:10.1126/science.aar5439 |
| [15] |
Walker B, Hollin C S, Carpenter S R, Kinzig A P. Resilience, adaptability and transformability in social-ecological systems. Ecology and Society, 2004, 9(2): 5. DOI:10.5751/ES-00650-090205 |
| [16] |
Rahman M, Islam M, Bräuning A. Species-specific growth resilience to drought in a mixed semi-deciduous tropical moist forest in South Asia. Forest Ecology and Management, 2019, 433: 487-496. DOI:10.1016/j.foreco.2018.11.034 |
| [17] |
Fang O Y, Qiu H Y, Zhang Q B. Species-specific drought resilience in juniper and fir forests in the central Himalayas. Ecological Indicators, 2020, 117: 106615. DOI:10.1016/j.ecolind.2020.106615 |
| [18] |
Huang M T, Wang X H, Keenan T F, Piao S L. Drought timing influences the legacy of tree growth recovery. Global Change Biology, 2018, 24(8): 3546-3559. DOI:10.1111/gcb.14294 |
| [19] |
Zhang Y D, Guo M M, Wang X C, Gu F X, Liu S R. Divergent tree growth response to recent climate warming of Abies faxoniana at alpine treelines in east edge of Tibetan Plateau. Ecological Research, 2018, 33(2): 303-311. DOI:10.1007/s11284-017-1538-0 |
| [20] |
庞瑞, 顾峰雪, 张远东, 侯振宏, 刘世荣. 西南高山地区净生态系统生产力时空动态. 生态学报, 2012, 32(24): 7844-7856. |
| [21] |
Myers N, Mittermeier R A, Mittermeier C G, da Fonseca G A B, Kent J. Biodiversity hotspots for conservation priorities. Nature, 2000, 403(6772): 853-858. DOI:10.1038/35002501 |
| [22] |
余莲, 封彩云. 青藏高原近期气候变化研究进展. 高原山地气象研究, 2012, 32(3): 84-88. DOI:10.3969/j.issn.1674-2184.2012.03.018 |
| [23] |
吴成启, 唐登勇. 近50年来全球变暖背景下青藏高原气温变化特征. 水土保持研究, 2017, 24(6): 262-266, 272-272. |
| [24] |
郭明明, 张远东, 王晓春, 黄泉, 杨素香, 刘世荣. 升温突变对川西马尔康树木生长的影响. 生态学报, 2015, 35(22): 7464-7474. |
| [25] |
四川森林编辑委员会. 四川森林. 北京: 中国林业出版社, 1992.
|
| [26] |
谢学强, 李云松, 张世萍, 吴晓东. 道孚县青稞生产现状与技术对策. 黑龙江农业科学, 2014(9): 160-163. DOI:10.3969/j.issn.1002-2767.2014.09.039 |
| [27] |
王亚军, 张永, 邵雪梅. 河北太行山南段树木年轮指示的167年来相对湿度变化研究. 生态学报, 2019, 39(12): 4570-4578. |
| [28] |
Stokes M A, Smiley T L. An Introduction to Tree-Ring Dating. Chicago: The University of Chicago Press, 1968, 56-57. |
| [29] |
刘立斌, 许海洋, 郭银明, 梁辉, 芦晓明, 张慧, 梁尔源, 倪健. 基于树木年轮定量重建过去50年贵州典型森林优势树种的地上生物量与生产力变化. 生态学报, 2020, 40(10): 3441-3451. |
| [30] |
Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983, 43: 69-78. |
| [31] |
Cook E R, Holmes R L. Users Manual for Program ARSTAN. Tucson: Laboratory of Tree-Ring Research, University of Arizona, 1986.
|
| [32] |
Fang O Y, Zhang Q B. Tree resilience to drought increases in the Tibetan Plateau. Global Change Biology, 2019, 25(1): 245-253. DOI:10.1111/gcb.14470 |
| [33] |
Lloret F, Keeling E G, Sala A. Components of tree resilience: effects of successive low-growth episodes in old ponderosa pine forests. Oikos, 2011, 120(12): 1909-1920. DOI:10.1111/j.1600-0706.2011.19372.x |
| [34] |
Zhang Y X, Wilmking M, Gou X H. Changing relationships between tree growth and climate in Northwest China. Plant Ecology, 2009, 201(1): 39-50. DOI:10.1007/s11258-008-9478-y |
| [35] |
Zhang Q B, Cheng G D, Yao T D, Kang X C, Huang J G. A 2326-year tree-ring record of climate variability on the northeastern Qinghai-Tibetan Plateau. Geophysical Research Letters, 2003, 30(14): 1739. |
| [36] |
邵雪梅, 范金梅. 树轮宽资料所指示的川西过去气候变化. 第四纪研究, 1999(1): 81-89. DOI:10.3321/j.issn:1001-7410.1999.01.010 |
| [37] |
吕少宁, 李栋梁, 文军, 王磊, 刘蓉, 王欣. 全球变暖背景下青藏高原气温周期变化与突变分析. 高原气象, 2010, 29(6): 1378-1385. |
| [38] |
丁一汇, 张莉. 青藏高原与中国其他地区气候突变时间的比较. 大气科学, 2008, 32(4): 794-805. DOI:10.3878/j.issn.1006-9895.2008.04.08 |
| [39] |
梁慧敏, 魏江生, 贺敏, 周梅, 赵鹏武. 大兴安岭南段华北落叶松人工林径向生长对气候变化的响应. 温带林业研究, 2019, 2(3): 31-36, 62-62. DOI:10.3969/j.issn.2096-4900.2019.03.006 |
| [40] |
Shi C M, Schneider L, Hu Y, Shen M G, Sun C, Xia J Y, Forbes B C, Shi P L, Zhang Y D, Ciais P. Warming-induced unprecedented high-elevation forest growth over the monsoonal Tibetan Plateau. Environmental Research Letters, 2020, 15(5): 054011. DOI:10.1088/1748-9326/ab7b9b |
| [41] |
Hansson A, Dargusch P, Shulmeister J. A review of modern treeline migration, the factors controlling it and the implications for carbon storage. Journal of Mountain Science, 2021, 18(2): 291-306. DOI:10.1007/s11629-020-6221-1 |
| [42] |
Calderaro C, Cocozza C, Palombo C, Lasserre B, Marchetti M, Tognetti R. Climate-growth relationships at the transition between Fagus sylvatica and Pinus mugo forest communities in a Mediterranean mountain. Annals of Forest Science, 2020, 77(3): 63. DOI:10.1007/s13595-020-00964-y |
| [43] |
杨婧雯. 东北林线偃松径向生长特征及其对气候变暖的响应[D]. 哈尔滨: 东北林业大学, 2020.
|
| [44] |
张晴, 于瑞德, 郑宏伟, 杨美琳, 甘淼. 天山东部不同海拔西伯利亚落叶松对气候变暖的响应分析. 植物研究, 2018, 38(1): 14-25. |
| [45] |
Jiang P, Liu H Y, Wu X C, Wang H Y. Tree-ring-based SPEI reconstruction in central Tianshan Mountains of China since A.D. 1820 and links to westerly circulation. International Journal of Climatology, 2017, 37(6): 2863-2872. DOI:10.1002/joc.4884 |
| [46] |
Palombo C, Battipaglia G, Cherubini P, Chirici G, Garfì V, Lasserre B, Lombardi F, Marchetti M, Tognetti R. Warming-related growth responses at the southern limit distribution of mountain pine (Pinus mugo Turra subsp. mugo). Journal of Vegetation Science, 2014, 25(2): 571-583. DOI:10.1111/jvs.12101 |
| [47] |
吴燕良, 甘淼, 于瑞德. 天山东部西伯利亚落叶松径向生长对气候的响应及其物候模拟. 应用与环境生物学报, 2019, 25(6): 1301-1311. |
| [48] |
吴普, 王丽丽, 黄磊. 五个中国特有针叶树种树轮宽度对气候变化的敏感性. 地理研究, 2006, 25(1): 43-52. DOI:10.3321/j.issn:1000-0585.2006.01.006 |
| [49] |
申静霞, 李迈和, 于飞海, 贺云龙, 袁秀锦, 雷静品. 川西云杉幼苗非结构性碳水化合物对土壤温度和水分变化的响应. 生态学报, 2021, 41(2): 503-512. |
| [50] |
王婷. 天山中部不同海拔高度天山云杉林的生态学研究[D]. 武汉: 武汉大学, 2004.
|
| [51] |
李明财, 罗天祥, 朱教君, 孔高强. 高山林线形成机理及植物相关生理生态学特性研究进展. 生态学报, 2008, 28(11): 5583-5591. DOI:10.3321/j.issn:1000-0933.2008.11.042 |
| [52] |
郭滨德, 张远东, 王晓春. 川西高原不同坡向云、冷杉树轮对快速升温的响应差异. 应用生态学报, 2016, 27(2): 354-364. |
| [53] |
吴秀臣, 裴婷婷, 李小雁, 刘鸿雁. 树木生长对气候变化的响应研究进展. 北京师范大学学报: 自然科学版, 2016, 52(1): 109-116. |
| [54] |
马姜明. 元谋干热河谷地区人工林稳定性及人工生态系统管理的研究[D]. 北京: 中国林业科学研究院, 2004.
|
| [55] |
Vayreda J, Martinez-Vilalta J M, Gracia M, Canadell J G, Retana J. Anthropogenic-driven rapid shifts in tree distribution lead to increased dominance of broadleaf species. Global Change Biology, 2016, 22(12): 3984-3995. DOI:10.1111/gcb.13394 |
| [56] |
Sánchez-Salguero R, Camarero J J, Carrer M, Gutiérrez E, Alla A Q, Andreu-Hayles L, Hevia A, Koutavas A, Martínez-Sancho E, Nola P, Papadopoulos A, Pasho E, Toromani E, Carreira J A, Linares J C. Climate extremes and predicted warming threaten Mediterranean Holocene firs forests refugia. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(47): E10142-E10150. DOI:10.1073/pnas.1708109114 |
| [57] |
沈维, 张林, 罗天祥. 高山林线变化的更新受限机制研究进展. 生态学报, 2017, 37(9): 2858-2868. |
| [58] |
Driscoll W W, Wiles G C, D'Arrigo R D, Wilmking M. Divergent tree growth response to recent climatic warming, Lake Clark National Park and Preserve, Alaska. Geophysical Research Letters, 2005, 32(20): L20703. DOI:10.1029/2005GL024258 |
| [59] |
Lloyd A H, Fastie C L. Spatial and temporal variability in the growth and climate response of treeline trees in alaska. Climatic Change, 2002, 52(4): 481-509. DOI:10.1023/A:1014278819094 |
| [60] |
Salzer M W, Hughes M K, Bunn A G, Kipfmueller K F. Recent unprecedented tree-ring growth in bristlecone pine at the highest elevations and possible causes. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106(48): 20348-20353. DOI:10.1073/pnas.0903029106 |
| [61] |
Jiao L, Chen K, Liu X P, Qi C L, Xue R H. Comparison of the response stability of Siberian larch to climate change in the Altai and Tianshan. Ecological Indicators, 2021, 128: 107823. DOI:10.1016/j.ecolind.2021.107823 |