 2021, Vol. 41
2021, Vol. 41文章信息
- 肖艳梅, 解婧媛, 姚义鹏, 梁士楚, 高丽娜, 张惠, 莫适祯
- XIAO Yanmei, XIE Jingyuan, YAO Yipeng, LIANG Shichu, GAO Lina, ZHANG Hui, MO Shizhen
- 桂林岩溶石山常绿落叶阔叶混交林乔木层优势物种生态位研究
- Niche of dominant species in arbor layer of evergreen deciduous broad-leaved mixed forest in karst hills of Guilin, southwestern China
- 生态学报. 2021, 41(20): 8159-8170
- Acta Ecologica Sinica. 2021, 41(20): 8159-8170
- http://dx.doi.org/10.5846/stxb202010142614
-
文章历史
- 收稿日期: 2020-10-14
- 修订日期: 2021-08-14





2. 广西师范大学生命科学学院, 桂林 541006;
3. 桂林航天工业学院, 桂林 541004
2. College of Life Science, Guangxi Normal University, Guilin 541006, China;
3. Guilin University of Aerospace Technology, Guilin 541004, China
生态位是指群落中各物种所能利用的各种资源总和, 及其与群落中其他物种的相互关系总和[1-2]。生态位测度主要包括生态位宽度和生态位重叠[3]。其中, 生态位宽度是指物种所能利用各种资源的一个综合指标[2], 反映物种对资源的适应状态和利用程度;生态位重叠则强调两个物种对同一资源的利用效率, 及不同物种或个体间的生态学特性可以相似到何种程度仍然能够共存, 即对资源利用相似性指标测度的问题。因此, 进行生态位研究时需首先对资源维度进行量化统计, 即从环境资源角度着手, 根据不同资源梯度的变化规律赋予数量等级值, 基于某一资源轴上的任意一组样方数量排序的位置关系, 进而计算并分析植物种群的生态位特征。该方法有助于探讨不同物种对同种资源的利用效率, 以及同一物种对不同资源的利用程度和物种间的共存关系。目前, 已有文献证实引起物种生态位分化的环境资源维度主要包括土壤理化性质[4-7]、地形[8]和气候[9-10]等环境因子。如王勤花[11]等研究发现, 锐齿栎林10个优势种群在土壤有机质、土壤pH值和土壤全氮3个资源维度上的生态位重叠都较大, 种群间利用资源的竞争排斥作用较为强烈, 彼此之间容易在各资源维度上形成竞争。郭平平等[12]对退化天坑物种生态位特征研究中, 发现土壤速效氮、速效钾和速效磷与植物分布关系最密切, 在3个环境下物种的生态位特征存在差异, 高重叠的种对在土壤速效磷环境梯度下最多, 速效氮次之, 速效钾相对较少。刘媛[13]在对岩溶石山土壤厚度对植物群落结构的研究中发现, 生态位互补效应随土壤厚度的增加而显著增加。
桂林岩溶石山所在的我国西南喀斯特地区是世界三大喀斯特集中连片发育区中面积最大、岩溶发育最典型和最强烈、石漠化最严重、人地矛盾最尖锐的地区, 也是景观类型复杂、生物多样性丰富、生态系统极为脆弱的地区[14-15]。长期以来, 桂林岩溶石山地区由于地质背景特殊, 景观异质性强, 岩溶作用强烈, 生态系统脆弱, 抗干扰能力差, 生态环境容量小, 但人口众多, 尖锐的人地矛盾导致植被破坏和水土流失日益严重, 石漠化等问题突出, 植被生态恢复成为桂林岩溶石山地区石漠化治理和生态重建的首要任务[16-17]。而桂林岩溶石山常绿落叶阔叶混交林作为桂林岩溶石山生态系统的主体, 在岩溶森林生态系统的恢复和保护中具有难以替代的作用和地位。因此, 研究桂林岩溶石山常绿落叶阔叶混交林的生态位特征, 明确其优势物种的生物学和生态学特性, 对岩溶地区退化生态系统植被恢复与重建具有重要意义。鉴于此, 本文以桂林岩溶石山常绿落叶阔叶混交林乔木层14个优势物种为研究对象, 分析其在6个不同土壤资源轴(土壤有机质、土壤水溶钙、土壤速效氮、土壤全氮、土壤pH值和土壤厚度)等级梯度上的生态位宽度和生态位重叠。旨在了解该地区不同生活型优势物种对资源利用的生态幅度和生态习性差异, 揭示影响桂林岩溶石山常绿落叶阔叶混交林植被分布的关键环境因子, 以期为岩溶石山植被恢复与重建、生物多样性保护等提供科学理论依据。
1 研究区概况研究区位于广西壮族自治区桂林市灵川县东部的灵田镇小水村后山, 地理坐标为25°19′04.52″—25°19′07.22″N, 110°25′10.36″—110°25′40.38″E。该地区岩溶地貌特征显著, 地貌类型主要由碳酸盐岩溶蚀为主所形成的峰丛谷地和峰丛洼地。气候属于中亚热带湿润季风气候, 雨热同期, 雨量充沛, 全年光照充足(年平均日照时数为1670 h);年平均气温19℃, 最冷的1月份平均气温8℃, 最热的8月份平均气温28℃, 全年无霜期309 d;年平均降雨量为1856.7 mm, 具有年内分配不均和秋、冬季干燥少雨等特点, 年平均蒸发量为1458.4 mm。研究区的主要植被类型为亚热带常绿落叶阔叶混交林, 主要常绿优势物种有青冈(Cyclobalanopsis glauca)、粗糠柴(Mallotus philippensis)、石山樟(Cmnamomum saxitilis)、檵木(Loropetalum chinense)、黄梨木(Boniodendron minius)、石岩枫(Mallotus repandus)、木犀(Osmanthus fragrans)等;主要落叶优势物种有南酸枣(Choerospondias axillaris)、光皮梾木(Cornus wilsoniana)、菜豆树(Radermachera sinica)、黄连木(Callicarpa bodinieri)、紫珠(Callicarpa bodinieri)、无患子(Sapindus saponaria)和大叶合欢(Archidebdron turgidum)等。
2 材料与方法 2.1 群落学调查2018年7—9月, 在桂林市灵田镇小水村的岩溶石山常绿落叶阔叶混交林内设置50个20 m×20 m的样方, 并以25°19′07.22″N, 110°25′39.44″E作为样地的坐标原点进行群落学调查。将每个20 m×20 m样方划分为4个10 m×10 m的小样方, 记录每个小样方内胸径(基径)≥1 cm的所有木本植物的物种名、相对坐标、多度、胸径、高度和健康状况以及每个样方的经纬度、海拔、坡向、坡度、岩石裸露率和土壤厚度等生境特征信息。据统计, 样地内存活的木本植物个体共有13012株, 隶属于37科77属96种, 其中常绿和落叶物种分别有53和43种。
2.2 土壤采样与测定采用“梅花五点法”, 在每个10 m×10 m小样方的4个顶点和中心位置, 采用插钎法测定土壤厚度(soil depth, SD, cm), 然后去除表层凋落物和腐殖质后, 用直径5 cm的土钻钻取土壤表层(0—20 cm)土样, 将采集的土壤样品装入密封袋带回实验室, 将其混合均匀置于室内阴凉通风处自然风干, 剔除其中的根系、石块、钙核及动植物残体等杂质后研细, 过不同孔径(0.15、0.25、0.9、1.7 mm)的土壤筛, 对预处理后的土样进行土壤有机质、土壤水溶钙、土壤速效氮、土壤全氮、土壤pH值、土壤速效钾和土壤速效磷等7个土壤化学性质测定, 测定方法参照《土壤农业化学常规分析方法》[18]。其中, 土壤有机质(soil organic matter, SOM, g/kg)含量采用重铬酸钾容量法(外加热法)测定;土壤水溶钙(water-soluble calcium, Ca, g/kg)含量采用氯化锶-火焰分光光度法测定;土壤速效氮(available nitrogen, AN, mg/kg)含量采用碱解扩散法测定;土壤全氮(total nitrogen, TN, g/kg)含量采用全自动凯氏定氮仪(KJELTECTM8400)测定;土壤pH值(pH value, pH)采用电位计法测定(水土比2.5∶1);土壤速效钾(available potassium, AK, mg/kg)含量采用乙酸铵-火焰分光光度法测定;土壤速效磷(available phosphorus, AP, mg/kg)含量采用碳酸氢钠提取-钼锑抗比色法测定。每个土壤样品重复测定3次, 取其平均值作为本研究分析的数据。
2.3 数据分析 2.3.1 优势物种的确定基于重要值=(相对显著度+相对多度+相对频度)/3[19]公式计算50个样方内各物种的重要值, 确定样地内出现的常绿和落叶优势物种名录。然后以其重要值作为排序依据, 筛选出重要值排序的前14个物种作为研究对象, 其中, 常绿和落叶优势物种各7种, 详见表 1。
| 编号 No. |
常绿Evergreen | 编号 No. |
落叶Deciduous | |||
| 优势物种 Dominant species |
重要值/% Importance value |
优势物种 Dominant species |
重要值/% Importance value |
|||
| 1 | 青冈Cyclobalanopsis glauca | 13.62 | 8 | 南酸枣Choerospondias axillaris | 14.20 | |
| 2 | 粗糠柴Mallotus philippensis | 9.94 | 9 | 光皮梾木Cornus wilsoniana | 8.94 | |
| 3 | 石山樟Cmnamomum saxitilis | 9.72 | 10 | 菜豆树Radermachera sinica | 5.51 | |
| 4 | 檵木Loropetalum chinense | 4.91 | 11 | 黄连木Callicarpa bodinieri | 2.75 | |
| 5 | 黄梨木 Boniodendron minius | 4.34 | 12 | 紫珠Callicarpa bodinieri | 1.76 | |
| 6 | 石岩枫Mallotus repandus | 2.31 | 13 | 无患子Sapindus saponaria | 0.94 | |
| 7 | 木犀Osmanthus fragrans | 1.97 | 14 | 大叶合欢Archidebdron turgidum | 0.86 | |
采用冗余分析(Redundancy analysis, RDA)结合蒙特卡罗随机置换检验(Monte Carlo random replacement test)筛选出对常绿和落叶优势物种分布影响显著的土壤资源轴。物种矩阵由50个20 m×20 m样方内14个优势物种的多度数据组成, 土壤资源轴矩阵由50个样方的8个土壤理化性质(土壤有机质、土壤水溶钙、土壤速效氮、土壤全氮、土壤pH值、土壤速效钾、土壤速效磷和土壤厚度)组成。排序结果结合蒙特卡罗999次随机置换检验筛选出对优势物种分布具有显著影响的土壤资源轴(土壤有机质、土壤水溶钙、土壤速效氮、土壤全氮、土壤pH值和土壤厚度)。然后, 初步采取等距间隔法[12]将土壤有机质、水溶钙、全氮、速效磷、pH值和厚度6个土壤资源轴, 从小到大划分为8个资源等级;但这样的划分没有充分考虑到每个资源轴等级下资源位分配样方数量的不均衡导致对生态位宽度和重叠统计结果的影响。因此, 为了避免不合理统计结果的产生, 进一步采取了将包含样方个体数较少的相邻等级进行适当合并, 将之前的8个资源等级最终划分为5个相对较均匀的等级区间(表 2)。
| 资源等级 Resource gradients |
有机质/(g/kg) Soil organic matter |
水溶钙/(g/kg) Water-soluble calcium |
速效氮/(mg/kg) Available nitrogen |
全氮/(g/kg) Total nitrogen |
pH值 pH value |
土壤厚度/cm Soil depth |
| 1 | 20.64—29.66 | 2.08—3.78 | 214.20—260.62 | 2.58—3.12 | 5.76—6.44 | 20.00—37.64 |
| 2 | 29.66—34.16 | 3.78—4.63 | 260.62—307.03 | 3.12—3.65 | 6.44—6.78 | 37.64—46.45 |
| 3 | 34.16—38.67 | 4.63—5.49 | 307.03—353.45 | 3.65—4.19 | 6.78—7.11 | 46.45—55.27 |
| 4 | 38.67—47.69 | 5.49—6.34 | 353.45—399.87 | 4.19—4.73 | 7.11—7.45 | 55.27—64.09 |
| 5 | 47.69—56.70 | 6.34—8.89 | 399.87—492.70 | 4.73—6.87 | 7.45—7.45 | 64.09—90.54 |
采用Levins生态位宽度指数(Bi)[20]测定物种间生态位宽度。
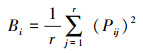
|
式中, Bi表示物种i的生态位宽度;Pij=nij/Ni, nij为物种i在资源位j上的重要值, Ni为物种i在所有资源位上重要值的总和, 即Pij表示物种i在资源位j上的重要值占所有资源位重要值总和的比例[19];r代表资源等级。
2.3.4 生态位重叠测定采用Pianka生态位重叠指数(Oik)[21]测定物种间的生态位重叠程度。
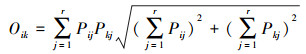
|
式中, Oik表示物种i和物种k的生态位重叠指数, 取值范围为[0, 1], 其值越大表示生态位重叠程度越高。Pij和Pkj分别表示物种i和k在资源位j上的重要值占所有资源位重要值总和的比例。
本研究中所有数据处理与分析均在Excel 2010和R 4.00软件中完成, 其中生态位宽度和生态位重叠分别由种间联结分析程序包spaa[22]中的niche.width()和niche.overlap()函数计算完成。
3 结果与分析 3.1 土壤资源轴对常绿落叶优势物种分布的影响RDA排序结合Monte Carlo随机置换检验结果表明:土壤有机质和土壤速效氮(P < 0.001), 土壤水溶钙和土壤pH值(P < 0.01), 土壤全氮和土壤厚度(P < 0.05)6个土壤资源轴对常绿落叶优势物种的分布产生显著影响(图 1)。其中, 常绿物种檵木和落叶物种紫珠倾向分布于土层较厚的生境;无患子、大叶合欢和黄连木等落叶物种倾向分布于土壤有机质、速效氮和全氮养分含量较高的生境;而青冈、粗糠柴等常绿物种倾向分布于土壤pH值、水溶钙含量及其它土壤养分含量较低的生境;光皮梾木和南酸枣落叶物种等则趋向分布于养分含量居中的生境。

|
| 图 1 土壤资源轴对常绿和落叶优势物种分布影响的RDA排序图 Fig. 1 RDA ordination diagram of the influence of soil resource axis on the distribution of dominant species RDA: 冗余分析Redundancy analysis; SOM: 土壤有机质Soil organic matter; AN: 速效氮Available nitrogen; Ca: 水溶钙Water-soluble calcium; pH: pH值pH value; TN: 全氮Total nitrogen; SD: 土壤厚度Soil depth; ***表示在0.001水平上显著;**表示在0.01水平上显著;*表示在0.05水平上显著 |
由表 3可看出, 在土壤有机质资源轴上, 常绿优势物种生态位宽度最大为檵木(0.89), 最小是木犀(0.56);落叶优势物种光皮梾木的生态位宽度最大(0.88), 紫珠和大叶合欢的生态位宽度最小(0.46)。在土壤水溶钙资源轴上, 常绿优势物种生态位宽度最大为檵木(0.94), 最小是木犀(0.53);落叶优势物种生态位宽度最大为菜豆树(0.80), 最小是黄连木(0.40)。
| 编号 No. |
常绿优势物种 Evergreen dominant species |
编号 No. |
落叶优势物种 Deciduous dominant species |
|||||||||||
| 有机质 SOM |
水溶钙 Ca |
速效氮 AN |
全氮 TN |
pH值 pH |
厚度 SD |
有机质 SOM |
水溶钙 Ca |
速效氮 AN |
全氮 TN |
pH值 pH |
厚度 SD |
|||
| 1 | 0.79 | 0.61 | 0.85 | 0.92 | 0.87 | 0.88 | 8 | 0.86 | 0.78 | 0.87 | 0.99 | 0.92 | 0.97 | |
| 2 | 0.85 | 0.86 | 0.83 | 0.95 | 0.96 | 0.83 | 9 | 0.88 | 0.76 | 0.82 | 0.88 | 0.79 | 0.89 | |
| 3 | 0.75 | 0.85 | 0.76 | 0.91 | 0.90 | 0.86 | 10 | 0.73 | 0.80 | 0.80 | 0.81 | 0.74 | 0.79 | |
| 4 | 0.89 | 0.94 | 0.72 | 0.62 | 0.85 | 0.76 | 11 | 0.48 | 0.40 | 0.53 | 0.45 | 0.25 | 0.56 | |
| 5 | 0.85 | 0.84 | 0.83 | 0.85 | 0.64 | 0.71 | 12 | 0.46 | 0.58 | 0.46 | 0.50 | 0.49 | 0.80 | |
| 6 | 0.81 | 0.65 | 0.70 | 0.82 | 0.67 | 0.62 | 13 | 0.56 | 0.59 | 0.48 | 0.48 | 0.53 | 0.77 | |
| 7 | 0.56 | 0.53 | 0.86 | 0.90 | 0.74 | 0.85 | 14 | 0.46 | 0.65 | 0.40 | 0.40 | 0.40 | 0.65 | |
| 均值 Mean |
0.79 | 0.75 | 0.79 | 0.85 | 0.80 | 0.79 | 均值 Mean |
0.63 | 0.65 | 0.62 | 0.64 | 0.59 | 0.78 | |
| SOM: 土壤有机质Soil organic matter;Ca: 水溶钙Water-soluble calcium;AN: 速效氮Available nitrogen;TN: 全氮Total nitrogen;pH: pH值pH value;SD: 土壤厚度Soil depth。1: 青冈Cyclobalanopsis glauca; 2: 粗糠柴Mallotus philippensis; 3: 石山樟Cmnamomum saxitilis; 4: 檵木Loropetalum chinense; 5: 黄梨木Boniodendron minius; 6: 石岩枫Mallotus repandus; 7: 木犀Osmanthus fragrans; 8: 南酸枣Choerospondias axillaris; 9: 光皮梾木Cornus wilsoniana; 10: 菜豆树Radermachera sinica; 11: 黄连木Callicarpa bodinieri; 12: 紫珠Callicarpa bodinieri; 13: 无患子Sapindus saponaria; 14: 大叶合欢Archidebdron turgidum | ||||||||||||||
在土壤速效氮资源轴上, 常绿优势物种生态位宽度最大为木犀(0.86), 最小是石岩枫(0.70);落叶优势物种生态位宽度最大为南酸枣(0.87), 最小是大叶合欢(0.40)。在土壤全氮资源轴上, 常绿优势物种生态位宽度最大为粗糠柴(0.95), 最小是檵木(0.62);落叶优势物种生态位宽度最大为南酸枣(0.99), 最小是大叶合欢(0.40)。
在土壤pH值资源轴上, 常绿优势物种生态位宽度最大为粗糠柴(0.96), 最小是黄梨木(0.64);落叶优势物种生态位宽度最大为南酸枣(0.92), 最小是黄连木(0.25)。在土壤厚度资源轴上, 常绿优势物种生态位宽度最大为青冈(0.88), 最小是石岩枫(0.62);落叶优势物种生态位宽度最大为南酸枣(0.97), 最小是黄连木(0.56)。总体而言, 在6个土壤资源轴上, 常绿优势物种的生态位宽度平均值均大于落叶优势物种, 说明常绿优势物种对6个土壤资源轴的利用能力更强, 适应范围更广。在土壤有机质、土壤水溶钙资源轴上, 生态位宽度最大值为常绿物种檵木;在土壤pH值资源轴上, 生态位宽度最大值为常绿物种粗糠柴;在土壤速效氮、土壤全氮和土壤厚度资源轴上, 生态位宽度最大值均为落叶物种南酸枣。
3.3 不同土壤资源轴上常绿落叶优势物种的生态位重叠在土壤有机质资源轴上, 生态位重叠值最大(0.99)的种对有常绿物种青冈与粗糠柴、常绿物种粗糠柴与石山樟、落叶物种黄连木与无患子、常绿物种石山樟与落叶物种菜豆树;生态位重叠值最小(0.10)的种对有落叶物种紫珠与黄连木、大叶合欢, 此外, 常绿物种木犀与落叶物种黄连木、大叶合欢、无患子(0.16、0.19、0.20)的生态位重叠值较小。在土壤水溶钙资源轴上, 生态位重叠值最大(0.98)的种对有落叶物种光皮梾木与无患子、常绿物种粗糠柴与落叶物种南酸枣、落叶物种光皮梾木与常绿物种石山樟、黄梨木;落叶物种黄连木与紫珠(0.08)的生态位重叠值最小(表 4)。
| Ca | SOM | |||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
| 1 | 0.99 | 0.97 | 0.91 | 0.65 | 0.65 | 0.89 | 0.96 | 0.94 | 0.95 | 0.34 | 0.90 | 0.41 | 0.29 | |
| 2 | 0.93 | 0.99 | 0.92 | 0.71 | 0.72 | 0.85 | 0.96 | 0.93 | 0.97 | 0.41 | 0.85 | 0.48 | 0.36 | |
| 3 | 0.94 | 0.97 | 0.86 | 0.63 | 0.67 | 0.80 | 0.92 | 0.87 | 0.99 | 0.33 | 0.84 | 0.41 | 0.25 | |
| 4 | 0.72 | 0.88 | 0.84 | 0.82 | 0.74 | 0.89 | 0.88 | 0.95 | 0.86 | 0.46 | 0.76 | 0.52 | 0.58 | |
| 5 | 0.83 | 0.89 | 0.95 | 0.83 | 0.96 | 0.50 | 0.74 | 0.78 | 0.63 | 0.85 | 0.41 | 0.88 | 0.91 | |
| 6 | 0.72 | 0.85 | 0.86 | 0.67 | 0.89 | 0.38 | 0.76 | 0.71 | 0.66 | 0.85 | 0.45 | 0.90 | 0.83 | |
| 7 | 0.91 | 0.84 | 0.82 | 0.77 | 0.70 | 0.50 | 0.80 | 0.90 | 0.79 | 0.16 | 0.78 | 0.20 | 0.19 | |
| 8 | 0.93 | 0.98 | 0.97 | 0.79 | 0.89 | 0.88 | 0.77 | 0.95 | 0.87 | 0.55 | 0.85 | 0.60 | 0.42 | |
| 9 | 0.90 | 0.90 | 0.98 | 0.77 | 0.98 | 0.88 | 0.74 | 0.92 | 0.83 | 0.55 | 0.77 | 0.59 | 0.49 | |
| 10 | 0.83 | 0.92 | 0.87 | 0.86 | 0.74 | 0.68 | 0.74 | 0.91 | 0.76 | 0.28 | 0.80 | 0.37 | 0.26 | |
| 11 | 0.36 | 0.53 | 0.60 | 0.52 | 0.79 | 0.84 | 0.20 | 0.54 | 0.70 | 0.30 | 0.10 | 0.99 | 0.83 | |
| 12 | 0.86 | 0.86 | 0.79 | 0.82 | 0.62 | 0.46 | 0.94 | 0.79 | 0.65 | 0.87 | 0.08 | 0.15 | 0.10 | |
| 13 | 0.86 | 0.81 | 0.92 | 0.65 | 0.95 | 0.83 | 0.70 | 0.84 | 0.98 | 0.62 | 0.71 | 0.55 | 0.85 | |
| 14 | 0.64 | 0.69 | 0.82 | 0.69 | 0.92 | 0.75 | 0.47 | 0.72 | 0.90 | 0.60 | 0.77 | 0.39 | 0.89 | |
| 右上角为优势物种在土壤有机质资源轴上的生态位重叠, 左下角为优势物种在土壤水溶钙资源轴上的生态位重叠 | ||||||||||||||
在土壤速效氮资源轴上, 生态位重叠值最大(0.99)的种对有常绿物种檵木与石岩枫、常绿物种青冈与落叶物种南酸枣、落叶物种光皮梾木与常绿物种粗糠柴、石山樟;生态位重叠值较小的种对有落叶物种紫珠与大叶合欢、无患子、黄连木(0.09、0.10、0.14)。在土壤全氮资源轴上, 生态位重叠最大(0.99)的种对有常绿物种青冈与石山樟、常绿物种木犀与落叶物种光皮梾木;生态位重叠值最小(0.50)的种对有落叶物种菜豆树与黄连木、常绿物种檵木与落叶物种紫珠(表 5)。
| TN | AN | |||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
| 1 | 0.97 | 0.97 | 0.81 | 0.85 | 0.76 | 0.82 | 0.99 | 0.97 | 0.88 | 0.58 | 0.51 | 0.55 | 0.44 | |
| 2 | 0.95 | 0.98 | 0.83 | 0.81 | 0.78 | 0.84 | 0.95 | 0.99 | 0.92 | 0.59 | 0.51 | 0.61 | 0.42 | |
| 3 | 0.99 | 0.93 | 0.87 | 0.84 | 0.82 | 0.84 | 0.94 | 0.99 | 0.92 | 0.61 | 0.38 | 0.62 | 0.43 | |
| 4 | 0.78 | 0.85 | 0.81 | 0.93 | 0.99 | 0.96 | 0.84 | 0.85 | 0.96 | 0.72 | 0.42 | 0.69 | 0.54 | |
| 5 | 0.85 | 0.97 | 0.82 | 0.85 | 0.94 | 0.93 | 0.85 | 0.84 | 0.85 | 0.85 | 0.39 | 0.78 | 0.76 | |
| 6 | 0.87 | 0.95 | 0.88 | 0.96 | 0.93 | 0.96 | 0.78 | 0.80 | 0.91 | 0.80 | 0.37 | 0.76 | 0.64 | |
| 7 | 0.87 | 0.90 | 0.89 | 0.85 | 0.89 | 0.92 | 0.85 | 0.85 | 0.95 | 0.77 | 0.60 | 0.74 | 0.62 | |
| 8 | 0.95 | 0.97 | 0.94 | 0.79 | 0.93 | 0.89 | 0.96 | 0.93 | 0.91 | 0.53 | 0.62 | 0.48 | 0.39 | |
| 9 | 0.88 | 0.89 | 0.91 | 0.84 | 0.86 | 0.89 | 0.99 | 0.96 | 0.90 | 0.67 | 0.43 | 0.68 | 0.51 | |
| 10 | 0.85 | 0.87 | 0.81 | 0.57 | 0.79 | 0.77 | 0.80 | 0.86 | 0.74 | 0.60 | 0.58 | 0.59 | 0.39 | |
| 11 | 0.67 | 0.79 | 0.68 | 0.96 | 0.81 | 0.91 | 0.71 | 0.66 | 0.68 | 0.50 | 0.14 | 0.98 | 0.96 | |
| 12 | 0.67 | 0.80 | 0.58 | 0.50 | 0.84 | 0.63 | 0.54 | 0.71 | 0.52 | 0.65 | 0.55 | 0.10 | 0.09 | |
| 13 | 0.73 | 0.82 | 0.73 | 0.92 | 0.82 | 0.88 | 0.65 | 0.68 | 0.65 | 0.51 | 0.97 | 0.66 | 0.92 | |
| 14 | 0.65 | 0.78 | 0.62 | 0.82 | 0.81 | 0.82 | 0.55 | 0.61 | 0.52 | 0.54 | 0.92 | 0.74 | 0.96 | |
| 右上角为优势物种在土壤速效氮资源轴上的生态位重叠, 左下角为优势物种在土壤全氮资源轴上的生态位重叠 | ||||||||||||||
在土壤pH值资源轴上, 生态位重叠值最大(0.98)的种对是常绿物种青冈与木犀、常绿物种粗糠柴与石山樟;其次是常绿物种青冈与粗糠柴、石山樟(0.97、0.96);落叶物种紫珠与大叶合欢、黄连木(0.09、0.10)和落叶物种黄连木与常绿物种黄梨木、木犀(0.10、0.15)生态位重叠值较小。在土壤厚度资源轴上, 常绿物种青冈与石山樟的生态位重叠值最大(0.99);落叶物种黄连木与菜豆树(0.45)、落叶物种黄连木与常绿物种黄梨木(0.50)的生态位重叠值相对较小(表 6)。
| SD | pH | ||||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 1 | 0.97 | 0.96 | 0.80 | 0.71 | 0.60 | 0.98 | 0.82 | 0.82 | 0.71 | 0.28 | 0.74 | 0.46 | 0.39 | ||
| 2 | 0.94 | 0.98 | 0.86 | 0.72 | 0.74 | 0.91 | 0.93 | 0.87 | 0.85 | 0.48 | 0.71 | 0.65 | 0.49 | ||
| 3 | 0.99 | 0.95 | 0.83 | 0.63 | 0.75 | 0.92 | 0.91 | 0.93 | 0.83 | 0.52 | 0.66 | 0.60 | 0.42 | ||
| 4 | 0.80 | 0.94 | 0.84 | 0.91 | 0.88 | 0.74 | 0.87 | 0.84 | 0.71 | 0.47 | 0.42 | 0.73 | 0.85 | ||
| 5 | 0.85 | 0.94 | 0.90 | 0.95 | 0.63 | 0.68 | 0.66 | 0.57 | 0.48 | 0.10 | 0.42 | 0.53 | 0.86 | ||
| 6 | 0.74 | 0.86 | 0.77 | 0.92 | 0.88 | 0.50 | 0.90 | 0.89 | 0.83 | 0.80 | 0.31 | 0.88 | 0.77 | ||
| 7 | 0.90 | 0.81 | 0.91 | 0.76 | 0.84 | 0.64 | 0.71 | 0.76 | 0.57 | 0.15 | 0.66 | 0.29 | 0.32 | ||
| 8 | 0.95 | 0.96 | 0.95 | 0.92 | 0.90 | 0.86 | 0.89 | 0.91 | 0.96 | 0.71 | 0.64 | 0.86 | 0.61 | ||
| 9 | 0.91 | 0.94 | 0.94 | 0.94 | 0.96 | 0.83 | 0.93 | 0.96 | 0.81 | 0.67 | 0.52 | 0.66 | 0.51 | ||
| 10 | 0.78 | 0.83 | 0.81 | 0.88 | 0.87 | 0.67 | 0.89 | 0.86 | 0.94 | 0.79 | 0.66 | 0.89 | 0.46 | ||
| 11 | 0.63 | 0.55 | 0.59 | 0.56 | 0.50 | 0.71 | 0.63 | 0.74 | 0.60 | 0.45 | 0.10 | 0.83 | 0.36 | ||
| 12 | 0.77 | 0.88 | 0.77 | 0.94 | 0.83 | 0.79 | 0.74 | 0.90 | 0.89 | 0.91 | 0.53 | 0.34 | 0.09 | ||
| 13 | 0.84 | 0.71 | 0.80 | 0.64 | 0.64 | 0.72 | 0.81 | 0.87 | 0.75 | 0.60 | 0.94 | 0.61 | 0.71 | ||
| 14 | 0.80 | 0.71 | 0.84 | 0.70 | 0.83 | 0.59 | 0.96 | 0.79 | 0.88 | 0.84 | 0.53 | 0.61 | 0.71 | ||
| 右上角为优势物种在土壤pH值资源轴上的生态位重叠, 左下角为优势物种在土壤厚度资源轴上的生态位重叠 | |||||||||||||||
14个优势物种组成91个种对, 其生态位重叠值≥0.8、0.5≤生态位重叠值< 0.8和生态位重叠值< 0.5的种对, 在土壤有机质资源轴上所占的比例分别为46.16%、27.47%和26.37%;在土壤水溶钙资源轴上所占的比例分别为52.74%、39.56%和7.70%;在土壤速效氮资源轴上所占的比例分别为50.55%、34.07%和15.38%;在土壤全氮资源轴上所占的比例分别为57.14%、42.86%和0;在土壤pH值资源轴上所占的比例分别为35.16%、42.86%和21.98%;在土壤厚度资源轴上所占的比例分别为60.44%、38.46%和1.10%。综合6个土壤资源轴得出, 共有546个种对, 其中275个种对的生态位重叠值≥0.8, 占总种对数的50.36%;205个种对0.5≤重叠值< 0.8, 占总种对数的37.55%;66个种对的生态位重叠值< 0.5, 占总种对数的12.09%。优势物种的生态位重叠程度较大, 说明混交林中各优势物种之间的利用资源方式和生态习性较为相似, 从物种竞争的角度来看, 优势物种在6个土壤资源轴上可能产生激烈竞争。
从生态位重叠值均值来看(表 7), 6个土壤资源轴上的14个优势物种组成91个种对的生态位重叠均值由大到小依次为土壤厚度资源轴(0.81)>土壤全氮资源轴(0.79) >土壤水溶钙资源轴(0.76)>土壤速效氮资源轴(0.73)>土壤pH值资源轴(0.69)>土壤有机质资源轴(0.68)。且6个土壤资源轴上均表现为, 常绿优势物种与常绿优势物种形成的种对生态位重叠均值(0.84)>常绿优势物种与落叶优势物种(0.74)>落叶优势物种与落叶优势物种(0.66)。14个优势物种在土壤厚度资源轴上竞争最为激烈, 全氮次之, 有机质相对较小;此外, 常绿物种间的竞争最为激烈, 常绿物种与落叶物种间的竞争次之, 落叶物种间的竞争相对较小。
| 物种对(种对数) Species pairs(number of species pairs) |
有机质 SOM |
水溶钙 Ca |
速效氮 AN |
全氮 TN |
pH | 厚度 SD |
6个资源轴 6 resource axes |
| 常绿物种与常绿物种(21) Evergreen species and evergreen species |
0.79 | 0.82 | 0.88 | 0.89 | 0.80 | 0.86 | 0.84 |
| 落叶物种与落叶物种(21) Deciduous species and deciduous species |
0.58 | 0.69 | 0.59 | 0.69 | 0.62 | 0.76 | 0.66 |
| 常绿物种与落叶物种(49) Evergreen species and deciduous species |
0.68 | 0.77 | 0.73 | 0.79 | 0.64 | 0.80 | 0.74 |
| 所有种对(91) Total species pairs |
0.68 | 0.76 | 0.73 | 0.79 | 0.69 | 0.81 | 0.74 |
本研究发现土壤有机质和土壤速效氮(P < 0.001), 土壤水溶钙和土壤pH值(P < 0.01), 土壤全氮和土壤厚度(P < 0.05)6个土壤资源轴对常绿落叶优势物种的分布产生显著影响(图 1)。常绿物种檵木和落叶物种紫珠倾向分布于土层较厚的生境, 这两个物种分布在土壤资源轴第一个资源等级(20.00—37.64 cm)的数量很少, 均仅占其个体总数的4%。无患子、大叶合欢和黄连木等落叶物种倾向分布于土壤有机质、土壤氮养分含量和土壤pH值较高的生境, 而青冈和粗糠柴等常绿物种则倾向分布于土壤pH值、土壤水溶钙含量及其它土壤养分含量较低的生境。常绿和落叶物种作为常绿落叶阔叶混交林内两种重要的生活型组分, 是物种的关键功能属性, 通常在土壤养分资源轴上采取不同的生态策略和资源利用方式。如常绿物种采取降低生长速率, 提高养分利用效率的“缓慢投资-收益”型保守性策略适应环境, 光合作用产物的一部分用于根茎的生长, 另一部分投资于枝干的生长和叶片组织密度的提高, 以减小蒸腾作用, 增强储存水分和养分的能力, 从而增强对干旱和土壤贫瘠的适应能力[23]。相反, 落叶物种常分布于土壤比较肥沃的地区, 为最大限度地获取光资源, 在生长季节快速地获取更多的光照、水分和营养物质等以满足高生长和叶面积增长, 常采取提高生长速率, 降低养分利用效率的“快速投资-收益”型资源获取性策略适应环境, 以便在相对短暂的生长季内获得更多的碳水化合物, 故而采取落叶的方式回避干旱等逆境胁迫对树木生长造成的伤害和不利影响[24-25]。因此, 尽管两种生活型物种共存于同一生境中, 可能会竞争相同的资源, 但两者生态策略的差异减弱了常绿和落叶物种之间的竞争强度, 进而促进了物种共存。
4.2 不同土壤资源轴上常绿和落叶优势物种的生态位宽度分析生态位宽度是度量物种对资源利用和环境适应能力的重要指标[9]。一般来说, 物种的生态位宽度越宽, 意味着该物种在资源上可利用范围广或者对某一资源没有偏好, 表明它对生态环境适应能力越强, 对各种资源的利用越充分, 生存机会越大, 且物种在空间分布的范围更广泛[26]。本研究发现, 在土壤有机质和土壤水溶钙资源轴上, 常绿物种檵木(0.89、0.94)的生态位宽度大于落叶物种光皮梾木(0.88)和菜豆树(0.80), 且2个资源轴的生态位宽度均值表现为常绿物种(SOM 0.79、Ca 0.75)高于落叶物种(SOM 0.63、Ca 0.65)。说明桂林岩溶石山常绿落叶阔叶混交林中的常绿物种在土壤有机质和土壤水溶钙资源轴上具有较强的资源利用和环境适应能力, 分布范围较为广泛。檵木和光皮梾木在土壤有机质资源轴的5个资源等级都有分布, 且比较均匀。说明二者对土壤有机质的适应范围广, 利用能力强, 因而在有土壤机质资源轴上的生态位宽度最大;相反黄连木和大叶合欢对土壤有机质资源的可利用幅度窄, 主要分布在土壤有机质含量4和5两资源等级上, 在其他资源等级没有分布, 所以其生态位宽度最小。岩溶区具土壤钙含量高的特性, 限制了很多植物在岩溶区生长, 常绿物种檵木是岩溶区植被恢复的先锋物种, 具分布广、耐旱、适钙性等[27]适应岩溶区特殊生境的生物生态学特性, 所以在土壤水溶钙资源轴上生态位宽度最大。
在土壤全氮、速效氮、厚度和pH值4个资源轴上, 落叶物种南酸枣(0.99、0.87、0.97、0.92)的生态位宽度最大或较大, 说明南酸枣对土壤重要养分氮的利用能力强, 还对土壤厚度和土壤pH值资源轴的适应范围广。产生这样结果的原因可能是由于该混交林乔木层中南酸枣个体数量最多, 分布较广, 占总资源位的74%, 生长迅速、适应性强, 在14个优势物种中重要值排名第一, 为该群落的建群种, 在营造群落内部特殊生境和决定群落内物种构成等方面起主导作用[28], 对资源利用充分和环境适应能力最强, 故在此4个资源轴上生态位宽度最大或较大。
一些研究表明在不同环境梯度下, 重要值大小与生态位宽度存在正相关或不相关关系[29-30]。重要值大的物种一般具有较大的生态位宽度, 本研究中大部分结果也遵循此规律。但二者的大小并非完全一致, 因为重要值并不是影响生态位宽度的唯一因素[31]。如落叶物种黄连木的重要值不是最小, 但它在土壤水溶钙、pH值、厚度资源轴上的生态位宽度均为最小。这可能与频度有关, 黄连木在50个资源位仅分布于9个资源位, 占总资源位的18%, 造成了在这3个资源轴上的分布不均匀, 所以频度也是影响生态位宽度的因素之一, 这与钱逸凡等[32]对普陀山主要植物种生态位特征的研究结果一致。
4.3 不同土壤资源轴上常绿和落叶优势物种的生态位重叠分析生态位重叠是衡量物种对环境资源利用能力差异性和竞争关系的重要指标[14]。从资源利用的角度来看, 生态位重叠越大, 意味着物种间生态需求越相似, 反之则越相异[33]。从竞争的角度来看, 当资源相对匮乏的时候, 生态位重叠较大的物种间可能会产生较为激烈的资源竞争[34]。一些研究表明, 生态位宽度与生态位重叠之间存在联系, 比如正相关和负相关联系[35-36]。本研究发现, 生态位宽度大的优势物种, 其生态位重叠也一般较大。如常绿物种青冈与粗糠柴、落叶物种南酸枣与光皮梾木等生态位宽度较大的物种形成的种对, 在6个土壤资源轴上的生态位重叠值均较大。说明这些种对在资源利用策略上具有高度相似性。另外, 本研究还发现, 生态位宽度较小的落叶物种黄连木与无患子, 在土壤有机质、土壤速效氮、土壤全氮和土壤厚度4个土壤资源轴上的生态位重叠值较大。出现这种情况, 一方面可能由于黄连木与无患子都具喜光、耐干旱瘠薄和深根性等生物生态学特性, 因而在土壤有机质和氮养分以及土壤厚度资源轴上的利用方式和适应范围相似;另一方面, 这还可能与桂林岩溶石山恶劣的生境条件有关, 该地区地形复杂, 岩石裸露率高, 土层浅薄, 地形破碎, 土被不连续, 土壤含水量和土壤养分等资源匮乏。在有限资源条件下, 具有相似资源利用方式的优势物种间的竞争程度增加, 某些物种为共享稀缺的环境资源, 生态位宽度相对压缩, 多倾向于聚集在局部适宜的生境斑块中, 而在适宜生境斑块以外的空间分布较少。这就导致了尽管黄连木与无患子的生态位宽度较小, 但却在土壤有机质、土壤速效氮、土壤全氮和土壤厚度4个土壤资源轴上产生了较大的生态位重叠[37]。
同一物种对在不同土壤资源轴上生态位重叠出现较大差别。如常绿物种木犀与落叶物种黄连木、无患子、大叶合欢3个种对在土壤速效氮(0.77、0.74、0.62)和土壤厚度(0.63、0.81、0.96)资源轴上的生态位重叠值较大;但在土壤有机质(0.16、0.20、0.19)和土壤pH值(0.15、0.29、0.32)资源轴上的生态位重叠值却较小。表明同一物种对在一些资源轴上发生较大的生态位重叠, 必然在另一些资源轴上发生比较明显的生态位分离, 使物种间的相互作用趋于相互补充, 尽量减少竞争形成共存机制, 充分利用环境资源。这与段后浪等[38]对恒湖农场植物种群生态位研究结果具有一致性。
根据Gause假说的竞争排斥原理[39]得出生态位重叠仅是产生竞争的一个前提假设, 且这个假设不是必然的, 而是需要在具体的生境中观察种间生态学特性可以相似到什么程度产生竞争排除。但可以得出如果两个物种的生态位重叠值越大, 它们之间的竞争可能越激烈进而排除其他物种的存在。然而事实并非如此, 在资源条件充足的情况下, 只能说明物种占据了相似的生态位空间, 而不能说明两者一定产生竞争;只有在资源不足的条件下, 生态位高度重叠才可能会产生竞争和排除。桂林岩溶石山生境恶劣、土层浅薄、地形破碎、土被不连续、土壤含水量和土壤养分等资源匮乏, 在有限资源条件下, 高生态位重叠的物种间很可能会产生激烈竞争。但群落是一个具有相互作用、生态位分化的种群系统, 这些优势种群对群落的空间、时间资源利用等方面, 都趋向于相互补充, 形成共存, 而不是直接竞争。从本研究的结果来看, 在桂林岩溶石山常绿落叶阔叶混交林中常绿物种与常绿物种间的生态位重叠均值最高, 常绿物种与落叶物种间的次之, 落叶物种间的相对较小。说明在同一生活型常绿物种之间可能会产生相对最为激烈的竞争, 常绿物种与落叶物种间次之, 落叶物种间相对较小。钟军弟等[40]对广西木论铁榄群落物种进行生态位研究, 结果表明:重叠值>0.5的种对数占总种对数的90.9%, 认为物种重叠值较高, 种间竞争较为剧烈, 群落结构可能具有一定的不稳定性。本研究结果与其相似, 在6个土壤资源轴上, 生态位重叠值≥0.5的种对高达480对, 占总种对数的87.91%, 也进一步说明桂林岩溶石山常绿落叶阔叶混交林乔木层优势物种在岩溶石山这个资源匮乏且资源异质性高的生境中存在竞争激烈, 该混交林乔木层种间关系相对不稳定。
4.4 植被恢复与保护策略针对岩溶地区的破碎化生境, 如何根据特有生境特点进行物种的筛选和配置是植被恢复与重建过程中首要解决的问题[17]。本研究发现落叶物种更倾向于生长高土壤pH值、全氮、速效氮、有机质和水溶钙的生境, 而常绿物种更倾向于生长在土壤pH值、全氮、速效氮、有机质和水溶钙含量低的生境中, 据此可根据环境类型筛选适宜生长的物种, 做到因地制宜, 如在土壤pH值、全氮、速效氮、有机质和水溶钙含量较高的生境中, 多搭配无患子、大叶合欢等落叶物种。相反, 在土壤pH值、全氮、速效氮、有机质和水溶钙含量低的生境中, 多搭配青冈和粗糠柴等常绿物种。在土壤厚度较小的生境中, 多搭配青冈、黄连木和大叶合欢等耐瘠薄的物种。此外, 根据我们对混交林的生态位特征的研究结果发现, 生态位宽度较大的檵木和南酸枣等物种适合作为先锋物种, 搭配生态位重叠度较小的黄连木和无患子等物种作为伴生种, 将不同的物种但对生境具有相似要求的物种组合一起配置在群落的相应空间位置, 充分利用有限的资源, 使群落处于一种互补互利的协调关系, 从而维护群落的稳定性和多样性, 促进群落可持续发展[14]。
5 结论本研究结论:(1)桂林岩溶石山常绿落叶阔叶混交林乔木层中, 常绿优势物种和落叶优势物种在土壤资源轴上采取不同的生态策略和资源利用方式;(2)物种生态位宽度大小受其重要值影响, 但不是唯一的决定因素, 可能还受物种分布频度的影响;(3)14个优势物种在土壤厚度资源轴上竞争最为激烈, 全氮次之, 有机质相对较小。且常绿物种间的竞争最为激烈, 常绿物种与落叶物种间的竞争次之, 落叶物种间的竞争相对较小;(4)综合6个土壤资源轴, 生态位重叠值≥0.5的种对高达480对, 占总种对数的87.91%, 暗示桂林岩溶石山常绿落叶阔叶混交林乔木层优势物种在资源匮乏且资源异质性高的特殊生境中存在激烈竞争, 且乔木层种间关系相对不稳定。
| [1] |
李燕芬, 铁军, 张桂萍, 郭华. 山西蟒河国家级自然保护区人工油松林生态位特征. 生态学杂志, 2014, 33(11): 2905-2912. |
| [2] |
刘润红, 常斌, 荣春艳, 姜勇, 杨瑞岸, 刘星童, 曾惠帆, 傅桂焕. 漓江河岸带枫杨群落主要木本植物种群生态位. 应用生态学报, 2018, 29(12): 3917-3926. |
| [3] |
郭水良, 于晶, 陈国奇. 生态学数据分析-方法、程序与软件. 北京: 科学出版社, 2015: 25-29.
|
| [4] |
Wagner V, Chytrý M, Zelený D, von Wehrden H, Brinkert A, Danihelka J, Hölzel N, Jansen F, Kamp J, Lustyk P, Merunková K, Palpurina S, Preislerová Z, Wesche K. Regional differences in soil pH niche among dry grassland plants in Eurasia. Oikos, 2017, 126(5): 660-670. DOI:10.1111/oik.03369 |
| [5] |
刘加珍, 陈亚宁, 张元明. 塔里木河中游植物种群在四种环境梯度上的生态位特征. 应用生态学报, 2004, 15(4): 549-555. DOI:10.3321/j.issn:1001-9332.2004.04.002 |
| [6] |
龚雪伟, 吕光辉. 艾比湖流域杜加依林荒漠植物群落多样性及优势种生态位. 生物多样性, 2017, 25(1): 34-45. |
| [7] |
赵学春, 来利明, 朱林海, 王健健, 王永吉, 周继华, 姜联合, 鲁洪斌, 赵春强, 郑元润. 三工河流域琵琶柴群落特征与土壤因子的相关分析. 生态学报, 2014, 34(4): 878-889. |
| [8] |
白晓航, 张金屯. 小五台山森林群落优势种的生态位分析. 应用生态学报, 2017, 28(12): 3815-3826. |
| [9] |
吴会峰, 宋丽娟, 杜峰, 郝文芳. 黄土丘陵区草地主要种群生态位及其环境解释. 应用生态学报, 2017, 28(11): 3494-3504. |
| [10] |
吴建国. 气候变化对7种荒漠植物分布的潜在影响. 应用与环境生物学报, 2010, 16(5): 650-661. |
| [11] |
王勤花, 巨天珍, 索安宁, 张俊华. 基于养分资源轴的锐齿栎林主要种群生态位研究. 西北师范大学学报: 自然科学版, 2005, 61(6): 61-65. DOI:10.3969/j.issn.1001-988X.2005.06.016 |
| [12] |
郭平平, 税伟, 江聪, 简小枚, 朱粟锋, 张永永, 冯洁, 陈毅萍. 退化天坑倒石坡林下优势物种生态位特征. 应用生态学报, 2019, 30(11): 3635-3645. |
| [13] |
刘媛. 喀斯特土壤厚度对植物群落结构及生物量的影响[D]. 重庆: 西南大学, 2020.
|
| [14] |
刘润红, 陈乐, 涂洪润, 梁士楚, 姜勇, 李月娟, 黄冬柳, 农娟丽. 桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结. 生态学报, 2020, 40(6): 2057-2071. |
| [15] |
王克林, 陈洪松, 曾馥平, 岳跃民, 张伟, 付智勇. 生态学研究支撑喀斯特区域生态环境治理与科技扶贫. 中国科学院院刊, 2018, 33(2): 213-222. |
| [16] |
李先琨, 苏宗明, 吕仕洪, 欧祖兰, 向悟生, 区智, 陆树华. 广西岩溶植被自然分布规律及对岩溶生态恢复重建的意义. 山地学报, 2003, 21(2): 129-139. DOI:10.3969/j.issn.1008-2786.2003.02.001 |
| [17] |
刘润红, 涂洪润, 李娇凤, 梁士楚, 姜勇, 荣春艳, 李月娟. 桂林岩溶石山青冈群落数量分类与排序. 生态学报, 2019, 39(22): 8595-8605. |
| [18] |
中国土壤学会农业化学专业委员会. 土壤农业化学常规分析方法. 北京: 科学出版社, 1983: 3-193.
|
| [19] |
张金屯. 数量生态学 (第三版). 北京: 科学出版社, 2018: 21-176.
|
| [20] |
Levins R. Evolution in Changing Environments: Some Theoretical Explorations. Princeton: Princeton University Press, 1968.
|
| [21] |
Pianka E R. The structure of lizard communities. Annual Review of Ecology and Systematics, 1973, 4: 53-74. DOI:10.1146/annurev.es.04.110173.000413 |
| [22] |
张金龙, 马克平. 种间联结和生态位重叠的计算: spaa程序包//马克平. 中国生物多样性保护与研究进展X. 北京: 气象出版社, 2014: 165-174.
|
| [23] |
Reich P B, Walters M B, Ellsworth D S. Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems. Ecological Monographs, 1992, 62(3): 365-392. DOI:10.2307/2937116 |
| [24] |
Grime J P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. The American Naturalist, 1977, 111(982): 1169-1194. DOI:10.1086/283244 |
| [25] |
Osnas J L D, Lichstein J W, Reich P B, Pacala S W. Global leaf trait relationships: mass, area, and the leaf economics spectrum. Science, 2013, 340(6133): 741-744. DOI:10.1126/science.1231574 |
| [26] |
梁玉莲, 国庆喜. 基于地形汇流值的东北天然次生林主要树种的生态位. 生态学报, 2010, 30(11): 2874-2882. |
| [27] |
梁月明, 李强, 潘复静. 岩溶区不同恢复阶段檵木根际土壤生态化学计量学特征. 水土保持通报, 2017, 37(5): 123-127. |
| [28] |
陈波, 周兴民. 三种嵩草群落中若干植物种的生态位宽度与重叠分析. 植物生态学报, 1995, 19(2): 158-169. DOI:10.3321/j.issn:1005-264X.1995.02.007 |
| [29] |
潘高, 张合平, 潘登. 南方红壤丘陵区3种森林群落内主要草本植物种群生态位特征. 草业科学, 2015, 32(12): 2094-2106. DOI:10.11829/j.issn.1001-0629.2015-0246 |
| [30] |
陈丝露, 赵敏, 李贤伟, 范川, 肖宝茹. 柏木低效林不同改造模式优势草本植物多样性及其生态位. 生态学报, 2018, 38(1): 143-155. |
| [31] |
汪建华, 周先容, 尚进, 江波, 杨利平. 金佛山巴山榧树灌丛群落主要木本植物种群生态位特征. 生态学杂志, 2014, 33(5): 1135-1141. |
| [32] |
钱逸凡, 伊力塔, 胡军飞, 张超, 余树全, 沈露, 彭东琴. 普陀山主要植物种生态位特征. 生态学杂志, 2012, 31(3): 561-568. |
| [33] |
史作民, 程瑞梅, 刘世荣. 宝天曼落叶阔叶林种群生态位特征. 应用生态学报, 1999, 10(3): 265-269. DOI:10.3321/j.issn:1001-9332.1999.03.003 |
| [34] |
陈玉凯, 杨琦, 莫燕妮, 杨小波, 李东海, 洪小江. 海南岛霸王岭国家重点保护植物的生态位研究. 植物生态学报, 2014, 38(6): 576-584. |
| [35] |
许诺, 林开敏, 吴培培, 许冠军, 朱晗, 何宗明. 杉木老龄林乔木层主要树种生态位研究. 森林与环境学报, 2017, 37(3): 330-335. |
| [36] |
铁军, 李燕芬, 金山, 陈艳彬, 张桂萍, 茹文明. 山西历山国家级自然保护区猕猴栖息地优势树种生态位特征. 生态学杂志, 2015, 34(3): 634-641. |
| [37] |
韩文衡, 李先琨, 叶铎, 吕仕洪, 向悟生, 宋同清, 曹洪麟. 桂西北喀斯特区常绿落叶阔叶混交林种群种间联结性与相关性. 山地学报, 2009, 27(6): 719-726. DOI:10.3969/j.issn.1008-2786.2009.06.012 |
| [38] |
段后浪, 赵安, 姚忠. 恒湖农场茶叶港草洲枯水期湿地植物与土壤关系及种群生态位分析. 生态学报, 2017, 37(11): 3744-3754. |
| [39] |
Gause G F. The Struggle for Existence. Baltimore: Williams & Wilkins, 1934.
|
| [40] |
钟军弟, 李先琨, 叶铎, 向悟生, 吕仕洪, 张建亮. 广西木论国家级自然保护区铁榄群落优势种群的生态位研究. 植物资源与环境学报, 2009, 18(3): 38-43. DOI:10.3969/j.issn.1674-7895.2009.03.006 |