 2021, Vol. 41
2021, Vol. 41文章信息
- 刘深思, 徐贵青, 李彦, 吴雪, 刘杰, 米晓军
- LIU Shensi, XU Guiqing, LI Yan, WU Xue, LIU Jie, MI Xiaojun
- 5种沙地灌木对地下水埋深变化的响应
- Difference and consistency of responses of five sandy shrubs to changes in groundwater level in the Hailiutu River Basin
- 生态学报. 2021, 41(2): 615-625
- Acta Ecologica Sinica. 2021, 41(2): 615-625
- http://dx.doi.org/10.5846/stxb202003310762
-
文章历史
- 收稿日期: 2020-03-31
- 网络出版日期: 2020-11-27





2. 中国科学院阜康荒漠生态国家野外科学观测研究站, 阜康 831505;
3. 中国科学院大学, 北京 100049;
4. 新疆大学资源与环境科学学院, 乌鲁木齐 830046
2. Fukang Station of Desert Ecology, Chinese Academy of Sciences, Fukang 831505, China;
3. University of China Academy of Sciences, Beijing 100049, China;
4. College of Resource and Environment Sciences, Xinjiang University, Urumqi 830046, China
在干旱、半干旱生态环境脆弱区, 水是限制植物种类、数量和生长发育的关键因素之一[1-2]。由于年降水量少、蒸发剧烈, 地下水成为植物生存所依赖的重要甚至是唯一的水分来源[3]。地下水的时空分布直接或间接影响生态系统过程、植物群落组成及植物用水策略[4-5]。流域尺度的地形地貌变化会影响地表到地下水的距离(地下水埋深), 而地下水埋深变化会约束植物土壤水分获取相关的生物量分配[4, 6-7]、功能性状表达及调控机制[8-10]。目前, 基于个体水平研究植物生理性状与地下水位变化之间的关系较为普遍, 如Gries等[11]在塔克拉玛干沙漠南缘发现胡杨叶水势、气孔导度与地下水位变化关系密切;张晓蕾等[12]研究表明地下水位的变化影响骆驼刺幼苗的蒸腾速率、光合作用等生理特性。然而, 相关研究多集中于探讨特定植物的生理性状对地下水位变化的响应和适应, 包括叶水势、气孔导度、净光合速率、叶绿素荧光参数、根系分布及生物量分配等指标的变化[13-17]。少有研究扩展到流域尺度, 基于地形地貌分异引起的地下水埋深变化, 认识不同灌丛植物对地下水埋深变化的响应差异。
内陆干旱区的地下水依赖型生态系统(Groundwater Dependent Ecosystems, GDEs)包含深根系和浅根系植被群落, 植物部分或全部依赖地下水[18]。地下水位和浅层地下水的空间分布直接影响着自然植被的生长和群落的空间分布结构[5]。改变地下水资源量将会导致地下水位的波动, 进而控制着水分对植物的有效性[17]。植物可以利用的水分不仅取决于生境的气候、土壤条件, 同时与植物根深、侧根分布及根系系统的重叠程度密切相关[19-20], 这些条件决定了植物吸收水分的范围[21]和对水分条件变化的响应及水分利用策略[22]。然而, 植被短期或长期对地下水埋深变化的响应仍然存在不确定性, 关于不同功能型植物汲取水分深度和生理调整的知识仍然不足[23]。
海流兔河流域属于我国典型的西北内陆干旱、半干旱地区, 土地沙漠化严重[24]。沙柳(Salix psammophila)、乌柳(Salix cheilophila)、柠条(Caragana korshinskii)、油蒿(Artemisia ordosica)、杨柴(Hedysarum laeve)等植物是海流兔河流域草原生态系统中的优势固沙灌木[25-26], 在维持生态系统稳定性和促进土地荒漠化防治中起关键作用。近年来, 由于城镇化水平的提高与能源工业产业的发展, 对地下水开采的需求增大, 导致区域内地下水位下降, 造成自然植被退化和局部荒漠化[26-28], 严重威胁GDEs的生态服务功能和系统稳定性[29-30]。该研究利用海流兔河流域产流区地形地貌变化和地势抬升形成的不同地下水埋深条件, 初步认识灌木生物量分配、根系分布和生理性状对地下水埋深变化的响应特征, 探讨不同灌木对地下水埋深条件变化的生理调整策略, 理论上有助于揭示流域乃至半干旱区内陆盆地中植物应对地下水埋深增加的抗旱生存维持机制, 实践上为沙生植被恢复的地下水资源管理提供参考。
1 研究区概况与研究方法 1.1 研究区域概况海流兔河流域位于鄂尔多斯盆地毛乌素沙漠与陕北黄土高原的过渡地带, 地处38°01′—38°42′N, 108°56′—109°36′E之间, 北起乌审旗的巴彦柴达木乡, 南达韩家峁, 西起乌审旗的查汗敖包, 东至榆林大海则三队。南北长91.3km, 东西宽109.8km, 流域面积达2603km2。流域内河网稀疏, 海流兔河为区内主要河流, 发源于乌审旗南部的新庙巴嘎, 自西北向东南汇入无定河, 全长120.9km, 此区内河长45.2km[31]。气候属温带大陆性季风气候, 夏季炎热多雨, 冬季寒冷干燥, 年平均气温为6.6—8.6℃, 年平均降水量为334.0—364.7mm, 年平均蒸发量约为2000mm, 属典型干旱、半干旱草原环境。
流域内地势特征表现为北高南低、东西高、中间低, 海拔高程在982.0—1479.5m之间[31]。在流域产流区以风沙地貌为主, 分布着滩地、沙地、沙丘、沙梁等地貌类型。滩地主要由河、湖相沉积形成, 地形平坦, 受较浅的地下水埋深和盐碱化影响;沙地分布面积广, 地形起伏小;沙丘地形起伏较大, 高度随固定程度发生变化, 滩地边缘的固定沙丘高度为1—4m, 半固定沙丘为7m;沙梁分布在流域分水岭地带, 相对高差达15—30m[32]。主要优势灌木有沙柳、杨柴、油蒿、乌柳、柠条、白沙蒿(Caryopteris mongholica)、沙棘(Hippophae rhamnoides)等。海流兔河流域地下水埋深的空间变化与灌木种类的多样性, 为认识不同灌木对地下水埋深变化的响应特征提供了理想的实验场地。
1.2 样地设置和地下水位调查在流域上游沿地形抬升设置1条样带, 间隔一定距离选取4个样地, 分别记为S1-1、S1-2、S1-3和S1-4。每个样地随机选取2—3个10m×10m的灌木样方, 沿海拔梯度跨越滩地、沙地、沙丘、沙梁等地貌类型。S1-1油蒿、杨柴、旱柳和沙柳分布;S1-2油蒿、柠条、沙柳和乌柳分布;S1-3油蒿、沙柳分布, S1-4仅出现油蒿。本研究选取沙柳、乌柳、柠条、油蒿、杨柴5种灌木作为研究对象。利用GPS定位系统, 确定样地中心经纬度位置与海拔高度。当地下水埋深小于3m时, 采用人工土钻法测定自然存在的地下水埋深;当地下水埋深超过3m时, 采用克里格空间插值的方法确定地下水埋深[31]。
1.3 根系分布与生物量测定选取高度和冠幅适中的沙柳、乌柳、柠条、油蒿和杨柴, 在植物生长末期进行全根系挖掘。记录植物主根深度、各级侧根数量、长度、深度分布范围及根直径。将每一吸收根视为圆柱体, 通过测得的直径和长度计算其表面积。以10cm为一个深度区间, 计算每个土壤深度区间内吸收根表面积, 绘制植物吸收根表面积垂直分布条形图[33]。同时, 对地上、地下生物量进行取样, 将样品带回实验室, 置于烘箱中以65℃烘烤24h, 称其干重, 全部生物量以烘干计重, 计算根冠比。
1.4 植物生理生态指标测定 1.4.1 小枝水势测定利用Model 3500植物水分压力室(PMS Instrument Company, Albany, NY, USA)分别于黎明前40min和正午测定黎明前小枝水势(predawn shoot water potential, ψpd)和正午小枝水势(midday shoot water potential, ψm)。每株植物上随机选取1个长势良好的枝条, 重复5次, 取数据平均值记为小枝水势值。
1.4.2 光响应曲线测定利用LI-6400便携式光合作用系统(Li-Cor, Lincoln, NE, USA)在晴天测量植株的光响应曲线。分别使用2×3cm2自带红蓝光源叶室和CO2注射器提供不同的光强和CO2浓度。叶室控温为30℃, 参照CO2浓度设为400μmol/mol, 流量设为400μmol/s。选择生长状况良好的成熟叶片进行测量, 测定光响应曲线时, 将叶平铺于叶室内, 尽量避免叶片重叠, 使叶表面接受同等的光照, 所有叶片的表面积可视为光合有效面积。测定结束后, 采集叶室内测定的叶, 计算叶面积, 再根据算出的实际叶面积折算光合速率。油蒿叶片狭窄而短小, 操作不便, 因而没有测量光响应曲线。
采用MnMolecular指数函数关系式对光响应曲线进行拟合, 公式如下[33]:
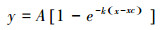
|
(1) |
式中, y是净光合速率(Pn), x是叶室内光量子通量密度(PPFDi), A为光饱和点的Pn(Ps), xc为光补偿点的PPFDi(Ic), k×A为光合作用表观量子效率(Φ)。通过拟合获得Ps、Ic和Φ等参数以衡量植物的光合能力。
1.4.3 气孔导度测定利用SC-1稳态气孔计(Model SC-1, Decagon, USA)测定植株的气孔导度(gs)。选取植株长势良好的成熟叶片, 将通道夹子夹在叶片上, 采取手动模式测量其气孔导度并记录数据, 实验重复3—9次。
1.4.4 叶绿素荧光测定利用Pocket PEA植物效率分析仪(Pocket PEA, PE 32 1JL, Hansatech Instruments Ltd, King′s Lynn Norfolk, UK)测定灌木叶片的初始荧光(F0)、最大荧光(Fm)和光化学最大量子产量(Fv/Fm)。每次测量选取数量相当的成熟叶片, 将其摊开均匀置于叶夹内, 选择冠层中有充足光照的成熟叶片进行测量, 重复6—15次。
1.5 数据处理利用SPSS 21.0软件(SPSS for Windows, Version 21.0, Chicago, IL, USA)对数据进行统计分析, 对5种灌木小枝水势、光合参数、气孔导度及叶绿素荧光数据进行单因素方差分析(One-way ANOVA)、独立样本T检验和Duncan多重比较;对沙柳各项生理指标与地下水埋深进行相关性分析, P < 0.05时表示差异显著。利用Sigmaplot 12.5 (Sigmaplot for windows, Version 12.5, Systat Software, USA)软件中的非线性回归模块对光合数据进行拟合及绘图。
2 结果 2.1 生物量分配与根系分布从表 1中可以看出, 不同样地灌木生物量的大小存在差异。S1-1—S1-4的灌木层地上生物量、地下生物量和总生物量呈降低的趋势, 表明灌木层生物量可能受到由地势抬升而引起的地下水埋深增加的限制。S1-2地上生物量小于地下生物量, 其它3个样地地上生物量均大于地下生物量, 这表明S1-2的灌木把更多的光合产物投入到地下部分, 这可以增强植物对地下水资源的获取能力[34]。4个样地的标准误均偏大, 表明不同样方生物量的变化非常大, 这可能是由于地下水埋深变化、人类活动如放牧、资源开采所导致[28, 32]。
| 调查点Sample plot | S1-1 | S1-2 | S1-3 | S1-4 |
| 经纬度 Longitude and Latitude |
38°44′32.2″ N108°48′38.3″ E | 38°46′38.1″ N108°48′48.1″ E | 38°47′17.9″ N108°45′10.5″ E | 38°48′02.2″ N108°42′33.9″ E |
| 海拔Altitude/m | 1326 | 1330 | 1380 | 1412 |
| 地貌类型Landform | 滩地 | 沙地、沙丘 | 沙丘 | 沙梁 |
| 地下水埋深groundwater level/m | 0.6 | 1.1、2.6 | 2.8 | 11.0 |
| 地上生物量 Aboveground biomass/(g/m2) |
432.80±390.62 | 235.04±57.97 | 112.39±34.47 | 69.68±59.39 |
| 地下生物量 Belowground biomass/(g/m2) |
362.83±332.64 | 281.24±167.44 | 77.46±16.54 | 40.90±31.76 |
| 总生物量 Total biomass/(g/m2) |
795.62±723.26 | 516.28±224.10 | 189.85±51.02 | 110.58±91.15 |
| 样本数n=2—3, 生物量数据采用平均值±标准误(Mean±SE)表示 | ||||
根系调查结果表明(图 1), 沙柳主根伸展的深度达500cm, 吸收根主要分布在地表以下10—120cm之间, 其吸收根表面积占总吸收根表面积的50%以上;在300—500cm深的土层中仍有13.21%的侧根分布。乌柳和柠条所有根系主要分布在地表下60cm深的土层中, 其中50%以上的吸收根表面积分布在0—20cm深的土层中。油蒿与杨柴50%以上的吸收根表面积分布在10—20cm的土壤深度。根据5种灌木的根系分布情况可知(图 1), 乌柳、柠条、油蒿和杨柴均为浅根系植物, 根系垂直深度不超过60cm, 侧根集中分布于浅层土壤;沙柳为深根系植物, 其主根发达, 侧根发育, 分布面积广。

|
| 图 1 5种灌木的吸收根表面积垂直分布与根冠比 Fig. 1 Vertical distribution of surface area of the feeders roots and root-shoot ratio |
由5种灌木的根冠比可知(图 1), 沙柳的根冠比最大, 乌柳、柠条、油蒿及杨柴根冠比均较小。这表明, 在典型的干旱、半干旱草原环境中, 沙柳更多的生物量投入地下以生长和发育庞大的根系, 从而在更大的土壤体积中获取水分, 相应的深根也有助于适应更广的地下水埋深范围。乌柳、柠条、油蒿及杨柴对地下生物量的投资较小, 其浅根系主要利用由降水转化而来的浅层土壤水。
2.2 滩地环境中灌木的生理性状特征小枝水势、净光合速率、气孔导度和最大光化学量子产量等生理指标可以反映植物在各种环境胁迫下的生理健康状态[35-38]。沙柳、乌柳、油蒿和杨柴在滩地共生环境中的生理性状特征如图 2所示。沙柳和乌柳黎明前小枝水势(ψpd)没有明显差异, 油蒿和杨柴ψpd也没有明显差异, 而前者的水分条件明显好于后者(P < 0.05)。油蒿正午小枝水势(ψm)最高, 为-0.94MPa, 其次是杨柴, 沙柳和乌柳ψm较低且不存在显著差异。与之相应地, 油蒿气孔导度(gs)大于另三种植物, 其中乌柳和杨柴的gs明显小于油蒿(P < 0.05)。从4种灌木的最大光化学量子产量(Fv/Fm)发现, 杨柴Fv/Fm显著高于其它三种灌木(P < 0.05), 为0.83;沙柳、乌柳和油蒿Fv/Fm差异不显著。在滩地共生环境中沙柳与乌柳的光合能力差异不显著, 3种植物表观量子效率(Φ)和光饱和点净光合速率(Ps)均不存在明显差异, 仅杨柴光补偿点光量子通量密度(Ic)显著大于沙柳与乌柳(P < 0.05)。

|
| 图 2 滩地共生环境中沙柳、乌柳、油蒿和杨柴小枝水势、气孔导度、最大光化学量子产量及光响应参数 Fig. 2 Shoot water potential, stomatal conductance, maximum quantum yield and light response parameters of Salix psammophila , Salix cheilophila , Artemisia ordosica and Hedysarum laeve in the flat land |
对丘间、丘顶同种植物的生理性状进行对比研究, 可以促进我们对植物策应地下水埋深变化的机理认识。图 3显示了丘间、丘顶分布下沙柳、乌柳、柠条和油蒿的ψpd、ψm、gs和Fv/Fm。沙柳丘间ψpd明显低于丘顶(P < 0.05), 预示着生长在丘顶的沙柳水分条件好于丘间;乌柳丘间ψpd明显高于丘顶(P < 0.05);柠条与油蒿丘间、丘顶ψpd差异不显著。正午时丘顶沙柳、柠条和油蒿小枝水势均显著高于丘间(P < 0.05);而乌柳丘间、丘顶ψm没有明显差异。沙柳、柠条和油蒿的gs对地形分异表现出一致性, 三者均没有明显变化。相比之下, 乌柳丘间、丘顶的gs则显示出显著差异(P < 0.05), 其丘间的gs明显大于丘顶。在两种地貌类型中, 4种植物的Fv/Fm值差异性均不显著。丘间沙柳的Ps和Ic显著高于丘顶, Φ没有明显差异;乌柳和柠条的光合能力没有受到微地形分异的限制, Ps、Ic和Φ均没有显著差异(图 4)。

|
| 图 3 典型丘间、丘顶分布下沙柳、乌柳、柠条和油蒿小枝水势、气孔导度及最大光化学量子产量 Fig. 3 Shoot water potential, stomatal conductance and maximum quantum yield of Salix psammophila , Salix cheilophila , Caragana korshinskii and Artemisia ordosica at the lowland and sandy dune |

|
| 图 4 典型丘间、丘顶分布下沙柳、乌柳和柠条表观量子效率、光补偿点光量子通量密度及光饱和点净光合速率 Fig. 4 Apparent quantum efficiency of photosynthesis, photosynthetic photon flux density at light compensation point and net photosynthetic rate at light saturation point of Salix psammophila , Salix cheilophila and Caragana korshinskii at the lowland and sandy dune |
在地势抬升条件下, 沙柳的水分状态受到地下水埋深变化的影响, 其结果如图 5所示。当地下水埋深为1.1m时, 沙柳ψpd和ψm较低;而当地下水埋深位于2.6m时, 沙柳ψpd和ψm均显著升高(P < 0.05)。沙柳gs与Ps对地下水埋深变化的响应一致, 线性回归分析的结果表明, 随着地下水埋深的增加, gs与Ps逐渐减小(P < 0.05)。当地下水埋深为0.6m时, 沙柳Fv/Fm最小, 为0.78;当地下水埋深为1.1m、2.6m、2.8m时, Fv/Fm不存在明显差异。沙柳Φ与地下水埋深的关系则呈现出显著的正相关, 即Φ随着地下水埋深的增加逐渐升高(P < 0.05), 而地下水埋深的变化并没有引起沙柳Ic的显著差异。

|
| 图 5 不同地下水埋深沙柳小枝水势、气孔导度、最大光化学量子产量及光合指标 Fig. 5 Shoot water potential, stomatal conductance, maximum quantum yield and photosynthetic parameters of Salix psammophila under different groundwater level |
地下水埋深是控制干旱区植被生长和分布的关键因素之一[26]。在降水稀少、蒸发剧烈的干旱区, 当地下水埋深较浅, 小于或等于植物根系深度时, 植物可能由于土壤盐渍化和土壤透气性差[10, 14, 32], 导致植被分布稀疏。当根系深入毛细水带时, 植物可以利用沿毛管上升而存在于土壤毛管孔隙中的地下水[39], 以维持正常生长。当地下水埋深过深, 植物根系不能触及毛管上升区时, 植物会因水分匮缺而影响生存[26, 32]。根据植物根系分布调查的结果(图 1), 这5种沙地灌木的根系分别属于两种功能型:深根型和浅根型。沙柳属于深根系植物, 它不仅利用表层土壤水, 而且能获取深层地下水, 这与前人的研究结果相似[40-41]。Zunzunegui等[42]研究表明, 在干旱季节, 这类植物的吸收根能够吸收深层水源以满足水分需求。相比之下, 乌柳、柠条、油蒿与杨柴均为浅根系植物, 其生存依赖于由降水转换而来的浅层土壤水和较浅的地下水。吕京京在海流兔河流域的研究结果表明[24], 这类灌木尽管不完全依赖地下水, 但对地下水很敏感。
植物功能性状能够响应生存环境的变化, 在一定程度上起到塑造植物生态策略的作用。由于植物功能性状的差异性, 使物种间形成不同获取、利用和保存资源的策略, 导致同一生活型植物具有不同的竞争优势和适应机制[43]。在地势平缓的滩地共生条件下, 油蒿和杨柴的生理性状存在显著差异, 油蒿ψm与gs明显高于杨柴, Fv/Fm明显低于杨柴(图 2)。任昱[44]等研究也表明, 相比杨柴, 油蒿具有较高的相对含水量和较低的叶绿素含量。而且根据张翠霞等[45]对沙生植物的研究表明, 油蒿在正午时气孔并未完全关闭, 其蒸腾作用仍保持较高水平, 这可能与油蒿叶片的形态结构密切相关。油蒿的叶表面具有表皮绒毛、角质膜蜡质层, 且气孔密度较高、气孔深陷, 这不仅能阻止水分蒸腾[46], 而且有利于叶片吸水[47]。此外, 肖春旺等人[48]发现, 无论水分的丰缺, 油蒿的株高、总枝长、叶数、以及叶、茎枝、根、总生物量干重均大于杨柴。因此, 油蒿对环境的适应能力更强、分布范围更广;而杨柴的适应能力较弱, 分布范围相对狭窄。
长期处于相同的选择压力下, 同一生活型植物在形态性状、生理性状与繁殖性状等方面表现出相似性, 从而呈现出趋同适应的现象[49]。Callaway[50]认为, 植物群落中的物种对特定物理环境具有相似的功能性状。本研究结果表明, 滩地共生环境中沙柳和乌柳具有相似的生理性状, ψpd、ψm、gs、Fv/Fm、Φ、Ps和Ic等生理生态指标均表现出一致性(图 2), 说明这两种灌木在滩地生境采取相同的生态策略。这一现象产生的原因与沙柳、乌柳之间的亲缘关系和形态结构有关, 根据刘玉冰等人[46]对干旱半干旱区荒漠植物叶片形态特征的研究表明, 尽管沙柳和乌柳属于不同的种, 但两种灌木同为柳属, 且都是典型的异面叶, 叶片上表皮覆被绒毛, 下表皮为柳属植物特有的蜡质层, 如沙柳的膜片状蜡质晶体、乌柳的壳状蜡质层。
丘间低地到沙丘的过渡, 除了海拔高度的直接变化, 地下水埋深变化最为明显。已有的研究表明, 地下水位变化会影响植物的生长发育和生理性状[51-53]。丘间、丘顶沙柳与乌柳的生理性状表现出差异性, 二者ψpd、ψm、gs、Ps和Ic的变化均不一致, 仅Fv/Fm和Φ相似(图 3、4), 这说明沙柳和乌柳的生理活动对不同地下水埋深的响应存在差异。乌柳ψpd和gs受到水分的限制, 但光合作用与荧光效率并没有表现对地下水埋深变化的显著响应(图 3、4), 表明地下水埋深对乌柳水分生理性状的影响更为强烈。柠条和油蒿的生理活动对地下水埋深变化的响应呈现出一致性, 生长在不同地貌类型的2种灌木ψpd、gs和Fv/Fm均没有明显变化, 但丘间ψm明显低于丘顶(图 3), 这一现象可能与浅地下水埋深盐分表聚限制根土界面土壤水分传输过程有关[54]。
沙柳在丘间-丘顶的生理特征, 可以间接反映其生长状态与地下水埋深之间的关系。然而, 关于沙柳的生理性状对地下水埋深变化的响应仍不明确。研究表明(图 5), 沙柳多项生理指标受到地下水埋深的显著影响。前人对多枝柽柳[10]的研究表明, ψpd随地下水埋深的增加而显著降低。本研究发现, 当地下水埋深从0.6m增加到1.1m时, 沙柳ψpd与ψm小枝水势显著降低;然而, 当地下水埋深从1.1m增加到2.6m时, ψpd与ψm明显升高(图 5)。这种现象出现的原因可能是:(1)在海流兔河流域, 当地下水埋深较浅时, 地下水在强烈的蒸发作用下可直接通过毛细上升作用携带矿物质到达地表, 导致表土层盐分积累[32], 沙柳受到盐分胁迫的限制, ψpd与ψm显著降低;(2)当地下水埋深介于0.6—1.1m时, 沙土土壤水分的贡献率占总蒸发量的80%以上[55], 土壤水分因蒸发散失而引起植物水势下降。此外, 随着地下水埋深的增加, gs和 Ps呈降低趋势, Φ则呈升高趋势, 这与前人的研究结果相似[12, 14, 51]。在自然环境下, 沙柳通过利用弱光进行光合作用、提高光能利用效率、减小光饱和时的光合能力和降低气孔导度的生态适应策略应对地下水埋深的增加(图 5)。
地下水位与植物功能性状的关系并不是简单的线性统计关系, 而是植物、土壤、地下水综合作用的复杂系统[13]。本文主要是研究5种沙地灌木对地下水埋深变化的生理响应, 还应充分考虑地下水水质、地下水位波动、土壤含水量及植物可塑性等影响因素, 加强高光谱遥感技术和同位素示踪技术的应用, 采用长期定位研究, 明确植物对地下水的吸收、利用情况。
4 结论(1) 沙柳为深根系植物, 根冠比大, 生理活动的维持不仅依赖浅层土壤水, 而且吸取地下水;乌柳、柠条、油蒿和杨柴为浅根系植物, 根冠比小, 主要水源是降水形成的浅层土壤水。
(2) 在滩地共生环境中, 沙柳和乌柳具有相同的适应机制, ψpd、ψm、gs、Fv/Fm、Φ、Ps和Ic等生理生态指标均表现出一致性;油蒿和杨柴的生理性状存在显著差异, 油蒿ψm与gs明显大于杨柴, Fv/Fm明显低于杨柴。
(3) 在丘间-丘顶分布下, 沙柳与乌柳对地下水埋深变化的生理响应表现出差异性, 二者ψpd、ψm、gs、Ps和Ic的变化均不一致, 仅Fv/Fm和Φ相同;柠条和油蒿对地下水位变化的生理响应呈现出一致性, ψpd、gs和Fv/Fm均没有明显变化, 丘间ψm都明显低于丘顶。
(4) 地下水埋深的变化显著地影响沙柳的生理性状。线性回归分析的结果表明, 随着地下水埋深的增加, 沙柳gs和Ps呈降低趋势, Φ呈升高趋势。
| [1] |
Dube O P, Geoff P. Effects of rainfall variability and communal and semi-commercial grazing on land cover in southern African rangelands. Climate Research, 2001, 17(2): 195-208. |
| [2] |
Watson R. Common themes for ecologists in global issues. Journal of Applied Ecology, 1999, 36(1): 1-10. DOI:10.1046/j.1365-2664.1999.00390.x |
| [3] |
Rodriguez-Iturbe I. Ecohydrology:A hydrologic perspective of climate-soil-vegetation dynamies. Water Resources Research, 2000, 36(1): 3-9. DOI:10.1029/1999WR900210 |
| [4] |
许皓, 李彦, 谢静霞, 程磊, 赵彦, 刘冉. 光合有效辐射与地下水位变化对柽柳属荒漠灌木群落碳平衡的影响. 植物生态学报, 2010, 34(4): 376-386. DOI:10.3773/j.issn.1005-264x.2010.04.003 |
| [5] |
Xu W, Su X S. Challenges and impacts of climate change and human activities on groundwater-dependent ecosystems in arid areas-A case study of the Nalenggele alluvial fan in NW China. Journal of hydrology, 2019, 573: 376-385. DOI:10.1016/j.jhydrol.2019.03.082 |
| [6] |
Horton J L, Clark J L. Water table decline alters growth and survival of Salix gooddingii and Tamarix chinensis seedlings. Forest Ecology and Management, 2001, 140(2/3): 239-247. |
| [7] |
Xu G Q, Yu D D, Li Y. Patterns of biomass allocation in Haloxylon persicum woodlands and their understory herbaceous layer along a groundwater depth gradient. Forest Ecology and Management, 2017, 395: 37-47. DOI:10.1016/j.foreco.2017.03.037 |
| [8] |
Horton J L, Kolb T E, Hart S C. Physiological response to groundwater depth varies among species and with river flow regulation. Ecological Applications, 2001, 11(4): 1046-1059. DOI:10.1890/1051-0761(2001)011[1046:PRTGDV]2.0.CO;2 |
| [9] |
Máguas C, Rascher K G, Martins-Loução A, Carvalho P, Pinho P, Ramos M, Correia O, Werner C. Responses of woody species to spatial and temporal ground water changes in coastal sand dune systems. Biogeosciences, 2011, 8(12): 3823-3832. DOI:10.5194/bg-8-3823-2011 |
| [10] |
张佩, 袁国富, 庄伟, 薛沙沙. 黑河中游荒漠绿洲过渡带多枝柽柳对地下水位变化的生理生态响应与适应. 生态学报, 2011, 31(22): 6677-6687. |
| [11] |
Gries D, Zeng F, Foetzki A, Arndt S K, Bruelheide H, Thomas F M, Zhang X, Runge M. Growth and water relations of Tamarix ramosissima and Populus euphratica on Taklamakan desert dunes in relation to depth to a permanent water table. Plant, Cell & Environment, 2003, 26(5): 725-736. |
| [12] |
张晓蕾, 曾凡江, 刘波, 张慧, 刘镇, 安桂香. 不同地下水埋深下骆驼刺幼苗叶片生理参数光响应特性. 干旱区地理, 2011, 34(2): 229-235. |
| [13] |
赵文智, 刘鹄. 荒漠区植被对地下水埋深响应研究进展. 生态学报, 2006, 26(8): 2702-2708. DOI:10.3321/j.issn:1000-0933.2006.08.037 |
| [14] |
玛丽娅·奴尔兰, 刘卫国, 霍举颂, 李宏侠, 张雨, 刘建国, 徐悦. 旱生芦苇对地下水位变化的生态响应及适应机制. 生态学报, 2018, 38(20): 7488-7498. |
| [15] |
Wu X, Zheng X J, Li Y, Xu G Q. Varying responses of two Haloxylon species to extreme drought and groundwater depth. Environmental and Experimental Botany, 2019, 158: 63-72. DOI:10.1016/j.envexpbot.2018.11.014 |
| [16] |
陈亚鹏, 陈亚宁, 徐长春, 李卫红, 付爱红. 塔里木河下游地下水埋深对胡杨气体交换和叶绿素荧光的影响. 生态学报, 2011, 31(2): 344-353. |
| [17] |
Naumburg E, Mata-Gonzalez R, Hunter R G, Mclendon T, Martin D W. Phreatophytic vegetation and groundwater fluctuations:A review of current research and application of ecosystem response modeling with an emphasis on great basin vegetation. Environmental Management, 2005, 35(6): 726-740. |
| [18] |
陈伟涛, 孙自永, 王焰新, 马瑞. 论内陆干旱区依赖地下水的植被生态需水量研究关键科学问题. 地球科学——中国地质大学学报, 2014, 39(9): 1340-1348. |
| [19] |
Schenk H J, Jackson R B. Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems. Journal of Ecology, 2002, 90(3): 480-494. DOI:10.1046/j.1365-2745.2002.00682.x |
| [20] |
XU G Q, Li Y. Rooting depth and leaf hydraulic conductance in the xeric tree Haloxyolon ammodendron growing at sites of contrasting soil texture. Functional Plant Biology, 2008, 35(12): 1234-1242. DOI:10.1071/FP08175 |
| [21] |
Lin G H, Phillips S L, Ehleringer J R. Monosoonal precipitation responses of shrubs in a cold desert community on the Colorado Plateau. Oecologia, 1996, 106(1): 8-17. DOI:10.1007/BF00334402 |
| [22] |
Xu H, LI Y, Xu G Q, Zou T. Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation. Plant, Cell & Environment, 2007, 30(4): 399-409. |
| [23] |
Antunes C, Chozas S, West J, Zunzunegui M, Barradas M C D, Vieira S, Máguas C. Groundwater drawdown drives ecophysiological adjustments of woody vegetation in a semi-arid coastal ecosystem. Global Change Biology, 2018, 24(10): 4894-4908. DOI:10.1111/gcb.14403 |
| [24] |
吕京京.海流兔流域地下水对植被指数分布的影响研究[D].北京: 中国地质大学(北京), 2013.
|
| [25] |
张国盛, 李玉灵, 王林和, 董智, 齐锦秋. 半干旱地区臭柏造林初步研究. 内蒙古林学院学报:自然科学版, 1999, 21(1): 22-26. |
| [26] |
金晓媚, 张强, 杨春杰. 海流兔河流域植被分布与地形地貌及地下水位关系研究. 地学前缘, 2013, 20(3): 227-233. |
| [27] |
周景山, 刘利清, 吴春霞, 于咏梅, 邢喜云, 白光梅. 内蒙古毛乌素沙地局部2009-2014年动态变化及其原因分析. 内蒙古林业调查设计, 2019, 42(6): 24-27. |
| [28] |
郝新忠, 郜超, 张继平, 石长春. 煤矿开采对榆林毛乌素沙地植被与地下水的影响. 陕西林业科技, 2019, 47(7): 28-31. |
| [29] |
刘鹄, 赵文智, 李中恺. 地下水依赖型生态系统生态水文研究进展. 地球科学进展, 2018, 33(7): 741-750. |
| [30] |
Eamus D, Froend R. Groundwater-dependent ecosystems:the where, what and why of GDEs. Australian Journal of Botany, 2006, 54(2): 91-96. DOI:10.1071/BT06029 |
| [31] |
朱丽, 徐贵青, 李彦, 唐立松, 牛子儒. 物种多样性及生物量与地下水位的关系——以海流兔河流域为例. 生态学报, 2017, 37(6): 1912-1921. |
| [32] |
张强.海流兔流域生态植被变化与环境影响因素研究[D].北京: 中国地质大学(北京), 2011.
|
| [33] |
Xu H, Li Y. Water-use strategy of three central Asian desert shrubs and their responses to rain pulse events. Plant and Soil, 2006, 285(1/2): 5-17. |
| [34] |
Bell D L, Sultan S E. Dynamic phenotypic plasticity for root growth in polygonum:A comparative study American. Journal of Botany, 1999, 86(6): 807-819. DOI:10.2307/2656702 |
| [35] |
冯建灿, 胡秀丽, 毛训甲. 叶绿素荧光动力学在研究植物逆境生理中的应用. 经济林研究, 2002, 20(4): 14-18, 30-30. DOI:10.3969/j.issn.1003-8981.2002.04.004 |
| [36] |
刘欣. 植物水势研究与应用综述. 吉林林业科技, 2015, 44(4): 35-37. |
| [37] |
王碧霞, 曾永海, 王大勇, 赵蓉, 胥晓. 叶片气孔分布及生理特征对环境胁迫的响应. 干旱地区农业研究, 2010, 28(2): 122-126, 131-131. |
| [38] |
王海珍, 韩路, 徐雅丽, 牛建龙, 于军. 干旱胁迫下胡杨光合光响应过程模拟与模型比较. 生态学报, 2017, 37(7): 2315-2324. |
| [39] |
伍光和, 王乃昂, 胡双熙, 田连恕, 张建明. 自然地理学(第四版). 北京:高等教育出版社, 2008, 320-322. |
| [40] |
杨峰, 刘立, 王文科, 赵贵章, 段鹏. 毛乌素沙地不同地貌下沙柳根系分布特征研究. 安徽农业科学, 2011, 39(25): 15582-15583, 15607-15607. DOI:10.3969/j.issn.0517-6611.2011.25.143 |
| [41] |
杨峰, 王文科, 刘立, 赵贵章, 段鹏. 毛乌素沙地沙柳根系与土壤水分特征的研究. 安徽农业科学, 2011, 39(26): 16050-16052. DOI:10.3969/j.issn.0517-6611.2011.26.087 |
| [42] |
Zunzunegui M, Boutaleb S, Díaz Barradas M C, Esquivias M P, Valera J, Jáuregui J, Tagma T, Ain-Lhout F. Reliance on deep soil water in the tree species Argania spinosa. Tree Physiology, 2017, 38(5): 678-689. |
| [43] |
杜峰, 梁宗锁, 胡莉娟. 植物竞争研究综述. 生态学杂志, 2004, 23(4): 157-163. DOI:10.3321/j.issn:1000-4890.2004.04.032 |
| [44] |
任昱, 魏春光, 郭小宇. 鄂尔多斯6种植物叶片功能性状比较. 内蒙古林业科技, 2019, 45(4): 43-46. |
| [45] |
张翠霞, 张秋良, 常金宝. 库布其沙漠几种植物的光合蒸腾及水分利用效率. 南京林业大学学报:自然科学版, 2007, 31(4): 81-84. |
| [46] |
刘玉冰, 李新荣, 李蒙蒙, 刘丹, 张雯莉. 中国干旱半干旱区荒漠植物叶片(或同化枝)表皮微形态特征. 植物生态学报, 2016, 40(11): 1189-1207. |
| [47] |
杨利贞, 冯丽, 杨贵森, 黄磊. 柠条(Caragana korshinskii)、油蒿(Artemisia ordosica)、花棒(Hedysarum scoparium)叶片吸水潜力及影响因素. 中国沙漠, 2020, 40(2): 214-221. |
| [48] |
肖春旺, 周广胜, 马风云. 施水量变化对毛乌素沙地优势植物形态与生长的影响. 植物生态学报, 2002, 26(1): 69-76. |
| [49] |
马淼, 李博, 陈家宽. 植物对荒漠生境的趋同适应. 生态学报, 2006, 26(11): 3861-3869. |
| [50] |
Callaway R M. Positive interactions in plant communities and the individualistic-continuum concept. Oecologia, 1997, 112(2): 143-149. |
| [51] |
刘世荣, 赵广东, 马全林. 沙木蓼和沙枣对地下水位变化的生理生态响应Ⅱ.叶片光合作用及其对温度和光的反应. 植物生态学报, 2003, 27(2): 223-227. |
| [52] |
朱成刚, 李卫红, 马建新, 马晓东. 塔里木河下游地下水位对柽柳叶绿素荧光特性的影响. 应用生态学报, 2010, 21(7): 1689-1696. |
| [53] |
侯荣哲, 杨娜, 孟甲, 蔺成龙, 张姣姣. 鄂尔多斯高原毛乌素沙地湖盆周缘植被与地下水关系研究. 地下水, 2012, 34(3): 10-11, 49-49. |
| [54] |
Huang F, Zhang Y D, Zhang D R, Chen X. Environmental groundwater depth for groundwater-dependent terrestrial ecosystems in arid/semiarid regions:A review. International Journal of Environmental Research and Public Health, 2019, 16(5): 763. |
| [55] |
李吉祥.毛乌素沙地土壤-地下水蒸发解析研究[D].西安: 西安科技大学, 2019.
|