 2021, Vol. 41
2021, Vol. 41文章信息
- 向杏信, 黄宗胜, 王志泰
- XIANG Xingxin, HUANG Zongsheng, WANG Zhitai
- 喀斯特多山城市空间形态结构与植物群落物种多样性的耦合关系——以安顺市为例
- Relationship between spatially morphology structure and species diversity of plant communities in Karst mountainous cities:a case study of Anshun city
- 生态学报. 2021, 41(2): 575-587
- Acta Ecologica Sinica. 2021, 41(2): 575-587
- http://dx.doi.org/10.5846/stxb201909191959
-
文章历史
- 收稿日期: 2019-09-19
- 网络出版日期: 2020-11-27





2. 贵州大学风景园林规划设计研究中心, 贵阳 550025;
3. 贵州大学建筑与城市规划学院, 贵阳 550025
2. Research Center for Landscape Architecture Planning and Design of Guizhou University, Guiyang 550025, China;
3. Building and Urban Planning College of Guizhou University, Guiyang 550025, China
当前城市化进程速度越来越快, 我国的城镇化率从2013年的53.7%增至2017年的58.52%[1], 土地资源利用开发强度越来越大, 土地利用效率在不同地区表现出较大的不均衡[2], 随之产生的生态环境、社会经济可持续发展等相关问题日益受到政府和众多学者的关注[3-5], 2018年组建自然资源部将履行国土空间用途管制、建立空间规划体系、生态保护修复等多项职责, 由此协调城市空间规划、生物多样性保护规划显得尤为重要[6]。城市空间形态结构是特定的地理环境下, 人类活动与自然环境相互作用的结果[7], 城市空间研究和城市生物多样性研究已成为当前城市快速发展背景下人居环境科学和生态学的研究热点[8]。城市空间形态结构研究主要集中在城市形态[9]、结构[10]、特征[11]、演变及动力学机制[12]等方面, 其中山地城市亦开展了空间结构模型[13]、空间格局规划[14]等研究, 但基于喀斯特这一生态脆弱与人文历史独特的山地城市空间形态结构及其量化的研究未见报道;城市生物多样性研究方面主要集中在植物多样性[15]、绿地景观格局[16]、植物群落物种多样性[17]、城市绿地系统多样性保护[18]等, 并且有向个体小的生物发展的趋势, 如草本植物多样性[19-20]、浮游植物多样性研究[21]等;在城市生物多样性影响因素方面主要开展了城市化[22]、绿地景观格局[23]对生物多样性的影响研究, 在山地城市涉及生物多样性的研究主要有生物多样性保护规划[24]、绿地系统构建[25]、景观格局[26]、城市森林物种多样性[27]。虽然, 关于城市空间和城市生物多样性两方面的研究已非常广泛, 但将城市空间形态结构与城市生物多样性二者结合在一起的研究并不多见, 尤其是针对特殊的多山城市的相关研究鲜见报道。城市生物多样性的形成、维持等是城市空间中的主要生态过程, 二者之间是否存在必然的关系, 城市空间形态结构对城市生物多样性的空间格局有没有影响, 这些问题对于多山城市的空间规划与生物多样性保护具有重要意义。基于此, 本研究选取山地城市中生态环境脆弱的喀斯特山地城市为研究对象, 以典型喀斯特山地城市—安顺市为例, 采用空间句法、生物多样性理论及其研究方法对喀斯特山地城市空间形态结构、绿地植物群落物种多样性及二者的关系进行初步研究, 旨在揭示城市空间形态结构与生物多样性的互作关系, 为城市空间规划、生物多样性保护规划与维持提供理论依据, 同时对城市国土空间规划上的“多规合一”途径具有重要参考意义。
1 材料和方法 1.1 研究区概况安顺市(25°21′—26°37′N, 105°13′—106°33′E), 位于贵州省中西部, 处于长江水系乌江流域和珠江水系北盘江流域的分水岭地带, 是世界上典型的喀斯特地貌集中地区, 喀斯特地貌占全域总面积84.9%, 平均气温在13.2—15.0℃, 属于“东部季风农业气候区”, 四季分明, 降水多集中于5至8月, 年平均气温14—16℃, 年平均无霜期280 d, 年平均相对湿度80%, 少日照、低辐射。境内大多为亚热带岩溶化高原山地, 地表山岭纵横, 孤立山体较多, 山体分布散乱无序, 且人为开垦的坡耕地较多, 土地荒漠化情况严重。本研究选取范围为安顺市主城区规划区(图 1), 面积约为264.61 km2。区域内地势北高南低, 海拔在1250—1560 m之间, 区域内较多建筑依山而建, 道路坡度较大, 且由于山体分隔道路较为曲折。

|
| 图 1 研究区地理区位图 Fig. 1 Location map of study area |
以安顺市2017年Pleiades高分辨率卫星影像图(0.5 m空间分辨率)为基础数据源, 通过人工解译及相关软件(Auto CAD、UCL Depth map)处理, 得到安顺市城区道路轴线模型, 以供空间句法相关指标测定。划定1000 m×1000 m网格覆盖整个研究区域, 选定网格交界处为取样点, 在取样点上设置植物群落调查样地, 通过实地调查研究区内样地植物群落及其物种多样性来获得基础数据。
1.3 研究方法 1.3.1 城市空间形态结构测定采用空间句法表征城市空间形态结构, 空间句法通过对包括建筑、聚落、城市甚至景观在内的人居空间形态结构的量化描述, 来表征空间及其组织与人类社会之间相互影响、相互作用的关系[28]。这种空间与人类社会之间的关系自然而然地会涉及空间内的物种多样性, 本研究选择空间句法中相关空间形态结构指标, 探讨空间形态结构对植物物种多样性的影响。选取指标如下[29], 见表 1。
| 指数Index | 计算方法Method of calculation | 指标含义Indicator meaning |
| 连接度(Ci) Connectivity value | 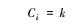
|
k为与节点i直接相连的节点个数, 在实际空间系统中, 一个空间的连接度越高, 表明该空间渗透性越好[30]。 |
| 整合度(Ii) Integration value | 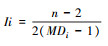
|
n为网络中总节点数(轴线数);MDi为平均深度值, 表征特定区域整体的空间属性, 反映了一个空间的可达性, 空间的整合度越高, 其可达性越高[30-32]。 |
| 平均深度值(MDi) Mean depth | 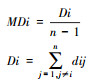
|
Di为全局深度值, n为网络中总节点数(轴线数);dij为i到j的深度值, MDi是一种相对的深度值, 数值越大, 该空间节点的便捷程度越低。 |
| 选择度Choice value | 反映某个空间出现在系统中其它任意两个空间的最短路径上的次数之和, 最短路径是指空间中任意一个元素到另一个元素的最短路径[30]。 | |
| 控制值(Ctrl) Control value | 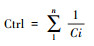
|
k表示与第i个节点相连的节点数;Ci表示道路i的连接度, 表示某空间节点对与之相交的空间的控制程度, 可反映局部空间之间的聚集程度[33-34]。 |
| 可理解度(R 2) Intelligible degree | 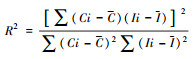
|
Ci为道路i的连接值;Ii为整合度;C为所有单元空间连接度的均值;I为所有单元空间整合度的均值。其反映了局部空间结构与整体空间结构的耦合程度, 可理解度越高, 局部空间与整体空间一致性越高, 越容易被认知理解[30] |
基于遥感影像, 为便于与空间结构形态有较高的吻合度, 通过空间尺度分析, 在整个研究区划定1000 m×1000 m网格, 网格交界处设取样点, 若取样点落在建筑、道路等无植物的硬化设施以及农田上, 则去除, 共获得有效取样点93处(图 1), 在样点上设置40 m×40 m的样地, 样地内设置10 m×10 m乔木样方4个, 每个乔木样方设5个4 m×4 m灌木样方、5个1 m×1 m草本样方[35]。
采用植物群落社会学调查方法[35]获得基础数据。在调查过程中发现人为干扰对物种数量和种类数目影响较大, 故选取物种多样性指数有:①Margalef指数(R), 仅考虑群落的物种数量和总个体数, 将一定大小的样本中的物种数量定义为多样性指数;②Shannon-Wiener(H′)指数, 主要体现区域内种类数目和种类之间个体分配均匀程度;③Pielou(Jh)指数, 反映区域中各物种在数量上的一致程度;④Simpson(D)指数, 表示样本中两个不同种之间个体相遇的概率[36]。计算公式为:
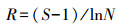
|
(1) |
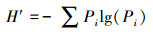
|
(2) |
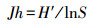
|
(3) |
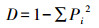
|
(4) |
式中, N为总个体数量;S为总物种数量;lg为以10为底的对数;ln为以e为底的自然对数;Pi=ni/N, ni表示第i个物种的个体数量。
1.3.3 城市空间形态结构与植物群落物种多样性相关关系测定采用相关性双变量分析法对城市空间形态结构指标与植物群落物种多样性指数进行相关性分析, 通过上述研究得到城市空间形态结构指标与植物群落物种多样性指数, 取植物群落样地内空间形态结构指标数据(因空间形态结构指数具有临近空间区域管辖性, 因此样点内如没有空间形态结构数据, 则取离样地最近的空间形态结构指标数据)与样地内物种多样性指标数据进行相关关系分析。
1.3.4 数据处理安顺市空间形态结构指标通过UCL Depth map 10软件实现;植物群落物种多样性Margalef指数、Shannon-Wiener指数、Pielou指数及Simpson指数等数据在Excel软件进行整理与处理;采用Pearson相关检验分析空间形态结构与植物群落物种多样性之间的相关性, 采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)比较不同绿地类型和不同方位植物群落物种多样性的差异, 显著性水平设定为α=0.05, 上述处理在SPSS Statistics 20软件中实现。
2 结果与分析 2.1 安顺市空间形态结构分析 2.1.1 安顺市整体空间形态结构特征基于空间句法理论, 通过分析轴线模型得到安顺市整体空间形态结构如表 2、图 2和图 3。表 2可知安顺市全局整合度、局部整合度均偏低, 表明安顺市整体可达性较差, 空间联系程度不高。图 2可以看出整体空间形态结构呈东南—西北向条块状分布(东南条块状由红色高整合度轴线组成, 西北条块状由西北方向黑色轴线组成), 东南向以黄果树大街与二环路所夹区域为主体, 其整合度高于西北向区域, 这两大区域主要由迎宾大道、工业大道相联系;安顺市整体空间形态结构中心性不强且较为分散, 中心性较高区域为:二环路、迎宾大道、黄果树大街、工业大道以及安顺环城高速东段区域;其中二环路不仅是整合度核心区域, 同时还连接了迎宾大道、工业大道和安顺环城高速这些中心性较强区域, 说明二环路在整个城市中有着举足轻重的作用, 是连接城市的交通枢纽;迎宾大道、工业大道作为连接城市两大块区域的主要交通道路, 其本身也是整合度核心区域, 说明在推动城市向北发展的过程中, 这两大区域作为重点开发对象, 在建设中优先于其它区域进行发展;安顺环城高速尽管在西北向区域整合度较高, 但仅有东段区域为城市整合度核心, 说明在城市发展过程中此区域发展强度不大, 在未来城市建设中仍是开发重点;安顺市平均道路密度较低, 城市道路密集区域集中在中部, 西部的迎宾大道与东部的工业大道为道路稀疏区域, 总体上安顺市道路空间分布不均衡, 道路网络集聚于较为发达的中心城区, 且道路密度在中心城区与东西部区域间有明显差异;轴线颜色由红色向蓝色过渡, 反映了空间整合度由高到低, 全局整合度最高区域主要集中在二环路、迎宾大道、黄果树大街, 说明在一定程度上这三条道路的人车流量更大, 可达性最高, 全局整合度较低区域集中在城市北部, 可能是因为北部尚处于在建的过程中, 在规划上保留有许多山体绿地, 可达性相对较低;局部整合度(半径=3)较高区域集中在黄果树大街, 其次为二环路, 城市北部区域局部整合度较低, 表明人流主要集中在黄果树大街与二环路区域。以上整体反映了喀斯特山地城市自然山体保留较多, 内部空间可达性较差, 联系程度较低, 因山体影响呈两个条块状发展趋势等特征。
| 轴线数量Number of axes | 全局整合度Global integration | 局部整合度Local integration | 连接度Connectivity value | 控制值Control value | 深度值Mean depth | 可理解度Intelligible degree | 选择度Choice value | 道路密度Road density |
| 13943 | 0.225 | 0.931 | 2.077 | 0.999 | 53.908 | 0.138 | 737642 | 3.92 |

|
| 图 2 安顺市空间形态结构整合度核心 Fig. 2 The core of integration degree of spatially morphology structure in Anshun City |

|
| 图 3 安顺市整体空间形态结构特征 Fig. 3 Characteristics of the overall spatially morphology structure in Anshun City |
图 4是通过分析轴线模型得到关于空间形态结构局部特征图, 由图可以看出:整体上安顺市的空间渗透性、聚集度、交通潜力低, 这反映了安顺市自然空间占较大比例, 人文社会空间不发达, 选择度较高区域与连接度较高区域、控制值较高区域三者之间存在高度重合, 表明此区域空间渗透性较好, 空间聚集程度较高, 其空间吸引穿越交通的潜力较大, 并且选择度、连接度、控制值较高区域仍然集中在黄果树大街和二环路, 进一步说明黄果树大街和二环路是主要的交通枢纽中心, 是连接城市其它区块的便捷通道, 其对周边区域的控制程度也是最大的, 同时也说明除黄果树大街和二环路外, 其他研究区域内空间渗透性、聚集度、交通潜力均较低;在迎宾大道区域也出现选择度较高的情况, 在城市向北扩张过程中, 人们在穿行城市活动中选择此区域通行的频率逐渐提高;中心城区和西南区域便捷程度较高, 在东部的东关办以及东北方向的蔡官镇区域内, 平均深度值远大于其它区域, 交通相对较为闭塞, 主要原因是与中心城区连接的区域为保留的山体绿地和农用地, 规划把此区域部分划分为生态廊道, 导致该区域与其他区域连接不便捷。

|
| 图 4 安顺市空间形态结构局部关系特征 Fig. 4 Characteristics of local relationship between spatially morphology structure in Anshun City |
可理解度主要是用来表征城市空间形态结构的认知程度, 由图 5可知:安顺市全局整合度和局部整合度拟合系数低, 可理解度数值仅为0.138, 说明安顺市整体可理解度不高, 人们行走于市区内的道路中, 难以通过几条街道或几片区域形成整体城市印象, 说明喀斯特山地城市空间多样性和异质性高、社会空间与自然空间交融, 使得空间丰富多样, 进一步反映了喀斯特山地城市因遗留自然山体较多, 城山交错, 从而形成了变化多样的、复杂的自然与社会融合的空间网络, 因而其可理解度较低;图中红色点表示全局整合度和局部整合度都较高的区域, 这些区域主要为黄果树大街、二环路以及这两条街道中间的众多小街道, 如顾府街等;红点聚集图像近似于一条回归直线, 其斜率远大于局部整合度和全局整合度形成的回归直线, 说明这些区域不仅是城市核心所在, 自身也具有较为鲜明的特色;安顺市山体较多, 道路走向、分布易受地形地势改变, 故而城市空间形态结构易受影响, 局部空间与整体空间拟合程度不高, 反映出安顺市保存有许多生态环境良好的自然与半自然空间。

|
| 图 5 可理解度 Fig. 5 Intelligible degree |
植物群落物种多样性测定结果见表 3, 可以看出:安顺市中心城区规划区内植物群落物种多样性4种指数均偏低, 安顺市正处于快速的城市发展建设过程中, 区域内植被生态系统可能受此影响, 导致植物群落物种多样性指数较低;不同样点内多样性指数差距较大, 某些样点所在生境被隔离开, 易造成生态孤岛, 或其生境本身为喀斯特岩石裸露较严重区域, 植物群落物种多样性指数相对较低, 有些则其所在区域或周边区域用地性质发生改变, 城市空间形态结构相应发生变化, 由此人为活动在此区域也更加频繁, 对区域内生态环境影响较大, 故多样性指数比其他未受干扰的样点低, 从而形成明显差异。
| 指数Index | R | H′ | D | Jh |
| 均值Mean value | 2.83±0.07 | 0.87±0.02 | 0.78±0.01 | 0.32±0.004 |
| 极小值Minimum | 0.58 | 0.25 | 0.30 | 0.18 |
| 极大值Maximum | 5.23 | 1.21 | 0.91 | 0.43 |
| R:马格列夫, Margalef;H′:香浓-维纳, Shannon-Wiener;D:辛普森, Simpson;Jh:均匀度指数, Pielou | ||||
通过对不同绿地类型中植物群落物种多样性进行分析得到结果如表 4:安顺市内不同绿地类型中植物群落物种多样性存在有差异性, 总体上区域绿地和公园绿地植物群落物种多样性数值相差不大, 且高于附属绿地、广场绿地和防护绿地;在Margalef指数上, 区域绿地高于附属绿地、广场绿地、防护绿地, 公园绿地也大于广场绿地;在Shannon-Wiener指数上, 区域绿地高于附属绿地、广场绿地和防护绿地;在Simpson指数上, 区域绿地高于附属绿地和防护绿地;在Pielou指数上, 各绿地类型之间并无显著差异。结果表明:区域绿地受人为活动影响较小, 其植物群落物种多样性指数较高;附属绿地、广场绿地、防护绿地受人工种植因素影响较大, 且大多数种植模式较为单一, 植物多样性指数偏低;安顺市公园大多为山体公园或依山而建的综合性公园, 人工种植绿化偏少, 故其植物多样性指数偏高。
| 绿地类型Type of green space | R | H′ | D | Jh |
| 公园绿地Park green space | 2.996±0.987ac | 0.875±0.076ab | 0.785±0.035ab | 0.300±0.016a |
| 防护绿地Protection green space | 2.018±0.288bc | 0.686±0.074b | 0.693±0.046b | 0.315±0.017a |
| 广场绿地Square green space | 1.605±0.506b | 0.580±0.117b | 0.693±0.048ab | 0.278±0.053a |
| 附属绿地Attached green space | 2.376±0.252bc | 0.743±0.055b | 0.700±0.038b | 0.295±0.014a |
| 区域绿地Regional green space | 2.911±0.127a | 0.898±0.030a | 0.792±0.015a | 0.316±0.006a |
| 具有相同字母表示无显著差异, 无相同字母表示具有显著差异 | ||||
表 5是对不同方位中植物群落物种多样性进行分析得到结果, 可以看出:安顺市不同方位之间植物群落物种多样性指数存在显著性差异, 整体上东北、东南方向植物多样性指数高于其他区域, 中心城区与西南区域植物群落物种多样性指数较低, 中心城区为政治、经济、文化的中心, 许多样地内的植物为园林绿化树种, 人们为了营造较为美观的植物绿化景观, 通常会进行人为干预, 如杀虫、除杂草、施肥等, 这些活动打破了生境内的自然演替, 是造成植物群落物种多样性较低的重要原因之一;其中Pielou指数仅在东北区域与中心城区存在有明显差异, 在一定程度上反映出人为活动对生境的影响表现在整体水平上, 这导致样点内均匀度指数可能会下降, 但相比其他指数并不十分明显, 其中变化较大的为Margalef指数, 表明人为活动对物种数量的影响最为明显。
| 方位Position | R | H′ | D | Jh |
| 东北Northeast | 3.29±0.20a | 0.97±0.04a | 0.83±0.02a | 0.33±0.01a |
| 东南Southeast | 2.95±0.27ac | 0.90±0.07ac | 0.78±0.04ac | 0.32±0.01ab |
| 西北Northwest | 2.62±0.16bc | 0.84±0.04ac | 0.76±0.02ac | 0.31±0.01ab |
| 西南Southwest | 2.13±0.23b | 0.76±0.05bc | 0.73±0.03bc | 0.30±0.01ab |
| 中心Center | 2.29±0.27bc | 0.67±0.06b | 0.68±0.03b | 0.29±0.02b |
| 具有相同字母表示无显著差异, 无相同字母表示具有显著差异 | ||||
表 6为采用相关性双变量分析法对城市空间形态结构指标与植物群落物种多样性指数进行相关性分析, 图 6为植物群落物种多样性总体分布图, 结果显示:在安顺市整体规划区域内, 选择度仅与Margalef指数呈显著负相关, 空间吸引人到达的潜力越大, 物种多度越小, 说明交通潜力对物种数量的影响相对较强, 对物种丰富度、均匀度影响较弱;连接度与各植物群落物种多样性指数都呈显著负相关, 城市空间渗透性越好的区域, 各植物群落物种多样性指数越低, 说明路网密集区域, 人类活动较为频繁, 对植物生境的干扰强度也较大;整合度与Margalef指数、Shannon-Wiener指数、Simpson指数呈显著负相关, 可达性越高的区域, 植物群落物种多样性指数越低(Pielou指数显著性不大), 说明人流和车流穿行量较大、城市空间聚集区域, 植物群落物种多样性指数一般较低;平均深度值仅与Shannon-Wiener指数呈显著正相关, 平均深度值越大, Shannon-Wiener指数越大, 进一步说明在便捷程度较低区域, 物种丰富度较高;而控制值与各植物群落物种多样性指数均无显著相关性。
| 指标Index | R | H′ | D | Jh |
| 选择度Choice | -0.213* | -0.137 | -0.125 | -0.053 |
| 连接度Connectivity | -0.333** | -0.325** | -0.348** | -0.223* |
| 控制值Control | -0.185 | -0.152 | -0.199 | -0.108 |
| 整合度Integration | -0.317** | -0.351** | -0.295** | -0.151 |
| 平均深度MeanDepth | 0.201 | 0.220* | 0.143 | 0.070 |
| n=93;* *. P < 0.01;*. P < 0.05 | ||||

|
| 图 6 安顺市植物群落物种多样性指数 Fig. 6 Plant community species diversity index in Anshun City |
采用相关性双变量分析法对不同绿地类型中二者之间的相关关系进行分析, 结果如表 7:在不同绿地类型中仅附属绿地在空间形态结构与植物群落物种多样性间存在显著相关关系, 其他绿地类型无显著相关性。其中在附属绿地上, Margalef指数与选择度、连接度、整合度呈显著负相关与平均深度值呈显著正相关, 说明在可达性较高、选择度较高的空间区域, Margalef指数越低, 而在便捷程度越低区域, Margalef指数越高, 这反映出不同区域内附属绿地受人为活动干扰程度不同, 在可达性较低区域, 生境内植被受人为影响较弱, 能进行群落自然演替, 相较于人类聚集强度较大区域, 其多样性指数相对较高, Pielou指数与连接度呈显著负相关, 由于不同物种在外界干扰下所呈现的反应不同, 人为活动对不同物种数量影响不一致, 故空间聚集程度越高, Pielou指数越低。在公园绿地中, 公园的选址建设大多是为了满足人类的需求而建立在人相对集中的地方, 大多位于中心城区, 所在区域的城市空间形态结构指数差异不大, 另外安顺市大多数公园为山体公园, 其植被类型由人工与自然混合组成, 有些公园的植被组成存在有一定的同质性, 因而其城市空间形态结构与植物群落物种多样性仅有一定的相关关系。区域绿地大多分布于建成区外围, 其周围空间形态结构指数较低, 区域绿地主要由自然绿地组成, 为保障其在城市中的生态和环保作用, 一般区域绿地受严格保护并限制开发, 故植物群落物种多样性受人为干扰不大, 所以二者之间相关关系较弱。广场绿地受人为活动影响严重, 其本身所处区域一般为城市较为聚集区域, 空间形态结构较规则而统一, 区域内植物大多由人工种植, 植物群落物种多样性指数存在有太大的人为干扰性, 故空间形态结构与植物群落物种多样性之间的相关性较低。在防护绿地中, 为使得植被起到较好的卫生、隔离和防护等作用, 植被大多由人工种植而成, 其组成形式较为单一, 植物多样性指数在不同区域差异性不大, 其本身所在区域一般为工厂、铁路、公路等需要防护地段, 空间形态结构受这些用地分布影响表现出局部无规律状态, 故空间形态结构与植物群落物种多样性之间的相关关系较低。总体而言, 不同绿地类型因其性质和作用不同, 植物群落物种多样性差异明显, 有些绿地由于自身特性或依附于周边用地等原因, 其地理分布较为无序, 空间形态结构也表现出无规律状态, 因此二者之间相关关系较低, 但在附属绿地中, 空间形态结构对人为活动影响较大, 两者相关关系显著。综上表明, 尽管表 6表明城市绿地系统整体水平上植物群落物种多样性与城市空间形态结构关系显著, 但在表 7反映的绿地类型水平上, 由于局部城市人为活动干扰强烈、或城市用地性质不同、或喀斯特生境自身生态脆弱等因素, 导致空间形态结构与物种多样性关系欠佳, 因此在局部尺度上应该减少人为干扰、确定好适当的用地类型、维护好喀斯特生态系统平衡, 这将是未来喀斯特山地城市空间规划与管理应当重视的重要举措。
3 讨论 3.1 喀斯特山地城市空间形态结构与植物群落物种多样性关系特征有研究[37-38]显示城市空间形态结构、社会职能、生态环境质量之间存在某些映射关系, 本研究结果也表明了这一点。本研究表明, 从整体而言, 在城市周边区域, 城市空间形态结构指数数值较中心城区低, 但其植物群落物种多样性远高于中心城区(图 6), 由此可知在连接度、整合度、选择度和控制值较高区域, 人口聚集、干扰频度和强度较高, 区域内物种数量较少, 种类越来越简单, 物种组成分布渐趋混乱[39-40], 植物群落物种多样性也较低, 整体上两者之间呈负相关关系。
本研究显示, 安顺市整体空间形态结构呈东南向与西北向两大条块状, 不同于一般的平原城市, 喀斯特山地城市空间形态结构受地形地貌特征影响严重, 区域内地形起伏繁复, 平地较少, 总体用地高差变化不大, 有些城市建设用地为山体丘陵遭平整后所用, 部分建筑依山而建[41], 这也是喀斯特山地城市量化空间形态结构指标数值较低的一个重要原因。在城市扩张过程中, 不仅城市空间形态结构发生改变, 植物群落物种多样性在此过程中也有着明显差异性。表 7显示, 比较不同绿地类型植物群落物种多样性差异及与城市空间形态之间的关系可知, 不同绿地类型中多样性差异明显, 其中与城市空间形态结构相关性最大的为Margalef指数, 空间形态结构指数越高, 物种数量相对较低, 因植物多样性受影响因素较多, 尽管两者之间存在一定的相关性, 但在其它多样性指数之间相关性较弱, 由此可知, 城市空间形态结构深受地形地貌等自然条件和人类干扰活动这两大因素影响, 在喀斯特这一特殊地貌环境背景下, 局部人类活动较易改变城市生态环境及其空间形态结构, 并同时对植物群落物种多样性造成巨大影响, 因此越是局部尺度, 在进行空间规划设计时越要注意多重影响因素。
| 绿地类型Type of green space | 指标Index | R | H′ | D | Jh |
| 附属绿地 | 选择度 | -0.444* | -0.264 | -0.167 | -0.157 |
| Attached green space | 连接度 | -0.450* | -0.401 | -0.444 | -0.461* |
| 控制值 | -0.326 | -0.284 | -0.327 | -0.387 | |
| 整合度 | -0.507* | -0.398 | -0.346 | -0.038 | |
| 平均深度 | 0.498* | 0.397 | 0.349 | 0.040 | |
| 公园绿地 | 选择度 | -0.387 | -0.244 | -0.070 | -0.019 |
| Park green space | 连接度 | -0.533 | -0.564 | -0.572 | -0.332 |
| 控制值 | 0.179 | 0.117 | 0.126 | -0.152 | |
| 整合度 | -0.358 | 0.014 | 0.450 | 0.443 | |
| 平均深度 | 0.332 | -0.049 | -0.506 | -0.456 | |
| 区域绿地 | 选择度 | -0.030 | -0.060 | -0.048 | -0.047 |
| Regional green space | 连接度 | 0.032 | 0.039 | 0.039 | 0.025 |
| 控制值 | 0.196 | 0.244 | 0.167 | 0.229 | |
| 整合度 | -0.056 | -0.114 | -0.053 | -0.101 | |
| 平均深度 | -0.041 | -0.017 | -0.119 | -0.021 | |
| 广场绿地 | 选择度 | 0.526 | 0.628 | 0.532 | 0.068 |
| Square green space | 连接度 | 0.741 | 0.147 | -0.138 | -0.532 |
| 控制值 | 0.726 | 0.327 | 0.075 | -0.353 | |
| 整合度 | 0.317 | -0.302 | -0.297 | -0.790 | |
| 平均深度 | -0.314 | 0.303 | 0.295 | 0.790 | |
| 防护绿地 | 选择度 | 0.149 | 0.315 | 0.195 | -0.013 |
| Protection green space | 连接度 | -0.101 | 0.083 | 0.004 | 0.187 |
| 控制值 | -0.114 | 0.091 | 0.045 | 0.210 | |
| 整合度 | -0.084 | -0.169 | -0.357 | -0.262 | |
| 平均深度 | 0.058 | 0.164 | 0.353 | 0.250 | |
| n=93;* *. P < 0.01;*. P < 0.05. | |||||
Matthew Carmona等曾在《城市设计的维度》中提出我们需要从广泛的视野进行城市空间的设计[42], 本文仅探讨城市空间形态规划和植物群落物种多样性规划两者之间的协调关系, 旨在为喀斯特山地城市规划建设提供一定的参考。对城市空间形态结构合理规划, 能提高空间可达性、便捷性、渗透力、集聚性等社会属性[43], 同样, 植物群落物种多样性保护与建设规划合理, 有利于城市生物多样性保护, 能更好的保护城市的生态环境。喀斯特脆弱生态系统退化的一个重要的驱动力是强烈的人类活动干扰[44], 由本研究的相关性分析结果可知, 整体上城市空间形态结构与植物群落物种多样性呈显著负相关性, 因此在喀斯特山地城市生态环境脆弱区如何协调好两者之间的关系显得尤为重要。城市空间形态结构规划的主要目的之一就是让人类社会活动更为便捷, 不仅使空间形态结构符合美学原则, 同时注重人们在此活动中的感受, 但在此过程中区域内自然生境会受到不同程度的影响, 因此在城市空间形态结构规划中应适当加强对自然生境的保护, 在城市绿地建设中多采用乡土物种, 尊重植物生长客观规律和群落演替规律。植物群落物种多样性规划在对物种保护、缓冲区构建及生态廊道建立等过程中, 对城市空间形态结构的连通性、聚集度都有着较大的影响, 规划过程中应采取相应的措施以加强各用地之间的联系程度, 如构建生态型绿道等。由图 6可知:中心城区内部也出现有植物群落物种多样性较高的区域, 大多为城市内的自然山体和山体公园中, 这是因为在安顺创建国家园林城市过程中, 对城市内部的山体及山体公园规划保护措施采取得当, 自然山体与山体公园生境受干扰程度较低。在山地城市中, 城市空间发展适应于自然山水条件的引导和制约[14], 在此, 可以借鉴城市内部山体植物群落较好区域在中心城区的保护经验, 在城市扩张过程中减少人为对山体的干扰, 并对山体周围划定保护范围, 尽可能的不改变其周边的用地模式。本研究存在的不足之处在于山体及自然生境周边, 多大范围内的用地性质改变能对生境内生态环境造成影响尚不明确, 这个尺度仍需后续的深入研究与探讨。在下一步的研究工作中, 我们建议以山体或自然生境为参考的中心点, 从中心点往城市用地方向设置样带, 每隔一定距离(用半径R表示, 具体距离视具体用地性质情况而定), 测算空间形态结构指标, 并用不同半径R点上的空间形态结构指标与中心点山体或自然生境的物种多样性指标做相关性分析, 随着半径R增大, 当空间形态结构指标与物种多样性关系不再显著时为止, 这时的R值则为山体或自然生境周边R半径范围内用地性质改变对生境内生态环境造成影响的阈值, 这种阈值在不同方位可能有所不同(因为山体周边用地性质、开发强度等有所不同), 这需要在山体或自然生境周边设置多条样带, 最后绘制影响范围梯度图, 以此图为相关规划部门提供理论参考。
4 结论众多研究[18, 37-38]表明不同的城市空间形态结构会产生不同的空间物质功能, 而不同的空间物质功能又会对城市生态环境产生不同的影响, 本研究显示空间形态结构与绿地系统物种多样性存在耦合关系, 这进一步定量地论证了这种城市空间形态结构、物质功能、生态环境三者之间映射关系的存在, 也表明了本研究预期的意义与目标基本达到, 同时亦反映了本研究采用的分析方法是可行的、有效的。喀斯特山地城市受地形地貌影响城市核心较为分散, 安顺市整体空间形态结构呈东南—西北向两大条块状分布, 城市中心性不强, 整体可达性较差, 空间联系程度不高, 整合度核心较为分散。城市道路密度较低, 其密集区域在中部, 整体上城市空间形态结构可理解性不高, 空间网络结构规律性不强。城市空间渗透性、聚集度、交通潜力低。安顺市植物群落物种多样性指数偏低, 且在不同区域内多样性指数差异较大。不同绿地类型、不同方位等条件下植物群落物种多样性差异性明显。安顺市整体空间形态结构与植物群落物种多样性有显著的相关关系, 在城市空间形态结构指数较高区域, 人的聚集、活动较高, 区域内物种数量较少, 植物群落物种多样性也较低。在不同绿地类型中, 仅附属绿地在Margalef指数和Pielou指数上存在有显著相关性。在喀斯特山地城市中, 城市空间形态结构受地形地貌等自然条件和人类干扰活动影响严重, 相同的这两大因素也对植物群落物种多样性影响巨大, 因此二者之间的相关性显著。城市空间形态结构规划与植物群落物种多样性规划应协调进行, 通过采取相应措施来调节人类活动与自然环境之间的关系, 加强不同用地类型间的内在联系, 使之成为一个有机的整体。
| [1] |
党丽娟, 宋建军. 新时代全国地质调查需求研究. 中国国土资源经济, 2020, 33(1): 43-49, 55-55. |
| [2] |
刘世超, 柯新利. 中国城市群土地利用效率的演变特征及提升路径. 城市问题, 2019(9): 54-61. |
| [3] |
成文青, 陶宇, 吴未, 欧维新. 基于MSPA-连接度-空间句法的生态保护空间及优先级识别——以苏锡常地区为例. 生态学报, 2020, 40(5): 1789-1798. |
| [4] |
焦世泰, 王鹏, 陈景信. 滇黔桂省际边界民族地区土地资源可持续利用. 经济地理, 2019, 39(1): 172-181. |
| [5] |
宫聪.绿色基础设施导向的城市公共空间系统规划研究[D].南京: 东南大学, 2018.
|
| [6] |
林坚, 赵冰, 刘诗毅. 土地管理制度视角下现代中国城乡土地利用的规划演进. 国际城市规划, 2019, 34(4): 23-30. |
| [7] |
郑莘, 林琳. 1990年以来国内城市形态研究述评. 城市规划, 2002, 26(7): 59-64, 92-92. DOI:10.3321/j.issn:1002-1329.2002.07.016 |
| [8] |
刘滨谊. 走进中国人居发展新时代. 世界人居, 2017(1): 72-74. |
| [9] |
徐银凤, 汪德根, 沙梦雨. 双维视角下苏州城市空间形态演变及影响机理. 经济地理, 2019, 39(4): 75-84. |
| [10] |
张茜, 段城江, 周典, 王宇鹏, 郭莉. 深圳城市出行空间结构研究. 城市发展研究, 2019, 26(3): 16-23. |
| [11] |
聂春祺, 谷人旭, 王春萌, 许树辉. 城市空间自相关特征及腹地空间格局研究——以福建省为例. 经济地理, 2017, 37(10): 74-81. |
| [12] |
周玉璇, 李郇, 申龙. 资本循环视角下的城市空间结构演变机制研究——以海珠区为例. 人文地理, 2018, 33(4): 68-75. |
| [13] |
黄光宇. 山地城市空间结构的生态学思考. 城市规划, 2005, 29(1): 57-63. |
| [14] |
王力国.生态和谐的山地城市空间格局规划研究[D].重庆: 重庆大学, 2016.
|
| [15] |
赵娟娟, 宋晨晨, 刘时彦. 城市植物种类构成的特征分析——以厦门市为例. 西南大学学报:自然科学版, 2018, 40(7): 1-8. |
| [16] |
付凤杰, 刘珍环, 黄千杜. 深圳市福田区三维城市景观格局变化特征. 生态学报, 2019, 39(12): 4299-4308. |
| [17] |
何荣晓.城市化对植物多样性的影响[D].海口: 海南大学, 2016.
|
| [18] |
杨培峰, 蔡云楠. 生态问题导向的城市绿地系统规划建设模式探索——以太原城市为例. 城市发展研究, 2016, 23(9): 62-67. |
| [19] |
张楠, 董丽, 王靛, 王阔. 北京城市生态廊道草本植物组成及分布格局. 中国园林, 2018, 34(6): 94-99. |
| [20] |
张心欣, 翟俊, 吴军. 城市草本植物多样性设计研究. 中国园林, 2018, 34(6): 100-105. |
| [21] |
范小晨, 代存芳, 陆欣鑫, 范亚文. 金河湾城市湿地浮游植物功能类群演替及驱动因子. 生态学报, 2018, 38(16): 5726-5738. |
| [22] |
刘高慧, 肖能文, 高晓奇, 付刚, 闫冰, 李俊生, 白加德. 不同城市化梯度对北京绿地植物群落的影响. 草业科学, 2019, 36(1): 69-82. |
| [23] |
叶萌, 宋于洋, 田晓鸽. 新疆库尔勒孔雀公园景观格局与生态服务功能动态分析. 西北林学院学报, 2017, 32(3): 259-264. |
| [24] |
曾雨静.山地城市生物多样性保护规划策略研究[D].贵阳: 贵州大学, 2018.
|
| [25] |
刘骏. 山地城市绿地系统规划问题及对策研究. 中国园林, 2017, 33(9): 67-71. |
| [26] |
任梅, 王志杰, 王志泰, 曾雨静, 何立影. 黔中喀斯特山地城市景观格局指数粒度效应——以安顺市为例. 生态学杂志, 2018, 37(10): 3137-3145. |
| [27] |
牛丽娟.基于"森林城市"的山地中小城市空间结构优化研究探讨[D].重庆: 重庆大学, 2012.
|
| [28] |
比尔·希列尔.空间句法的发展现状与未来.盛强, 译.建筑学报, 2014, (8): 60-65.
|
| [29] |
程明洋, 陶伟, 贺天慈. 空间句法理论与建筑空间的研究. 地域研究与开发, 2015, 34(3): 45-52. |
| [30] |
张忠训, 杨庆媛, 王立, 苏康传, 匡成铭. 少数民族地区传统村落交通可达性分析——以贵州省铜仁市为例. 资源科学, 2018, 40(11): 2296-2306. |
| [31] |
张愚, 王建国. 再论"空间句法". 建筑师, 2004(3): 33-44. |
| [32] |
陈驰, 李伯华, 袁佳利, 余雯. 基于空间句法的传统村落空间形态认知——以杭州市芹川村为例. 经济地理, 2018, 38(10): 234-240. |
| [33] |
刘承良, 余瑞林, 段德忠. 基于空间句法的武汉城市圈城乡道路网通达性演化分析. 地理科学, 2015, 35(6): 698-707. |
| [34] |
周麟, 田莉, 张臻, 李薇. 基于空间句法视角的民国以来北京老城街道网络演变. 地理学报, 2018, 73(8): 1433-1448. |
| [35] |
金振洲. 植物社会学理论与方法. 北京: 科学出版社, 2009: 77-83.
|
| [36] |
钱迎倩, 马克平. 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 1994.
|
| [37] |
Desyllas J, Duxbury E. Axial maps and visibility graph analysis: a comparison of their methodology and use in models of urban pedestrian movement//Proceedings of the 3rd International Space Syntax Symposium, Georgia Institute of Technology, Atlanta, 2001.
|
| [38] |
Hillier B. Cities as movement economies. Urban Design International, 1996, 1(1): 41-60. |
| [39] |
Hillier B. The hidden geometry of deformed grids:or, why space syntax works, When it looks as though it shouldn't. Environment and Planning B:Planning and Design, 1999, 26(2): 169-191. |
| [40] |
Midgley G F. Biodiversity and ecosystem function. Science, 2012, 335(6065): 174-175. |
| [41] |
吴勇.山地城镇空间结构演变研究[D].重庆: 重庆大学, 2012.
|
| [42] |
Carmona M, Heath T, Taner T.城市设计的维度——公共场所——城市空间.冯江, 袁粤, 万谦, 傅娟, 张红虎, 译.南京: 江苏科学技术出版社, 2005.
|
| [43] |
Graham-Rowe D. Biodiversity:endangered and in demand. Nature, 2011, 480(7378): S101-S103. |
| [44] |
郭柯, 刘长成, 董鸣. 我国西南喀斯特植物生态适应性与石漠化治理. 植物生态学报, 2011, 35(10): 991-999. |