 2021, Vol. 41
2021, Vol. 41文章信息
- 王欢, 张萍, 王朔, 郭佳茹, 王博, 张昱, 秦海琴, 靳磊
- WANG Huan, ZHANG Ping, WANG Shuo, GUO Jiaru, WANG Bo, ZHANG Yu, QIN Haiqin, JIN Lei
- 柠条锦鸡儿和沙拐枣主根暴露后的生理响应及抗逆机理
- Physiological response and stress tolerance mechanisms of Caragana korshinskii and Calligonum mongolicum suffering from taproot exposure
- 生态学报. 2021, 41(2): 535-542
- Acta Ecologica Sinica. 2021, 41(2): 535-542
- http://dx.doi.org/10.5846/stxb201901020015
-
文章历史
- 收稿日期: 2019-01-02
- 修订日期: 2020-08-13





壤风蚀是发生于干旱、半干旱及部分半湿润地区土地沙漠化过程的首要环节, 也是固定沙丘活化, 植物种群更新与演替的影响因素之一[1]。合理选择搭配防风固沙植物可有效控制风沙活动、维持当地生态稳定[2]。沙拐枣和柠条锦鸡儿是流动半流动沙丘(沙地)上草方格沙障内常套种的固沙植物种[3-5], 揭示固沙植物的抗风蚀生理生化调节机制, 对今后的防风固沙选种及抚育工作具有指导意义。
风蚀对植物的影响主要有两方面, 一是风沙流对植物的拍打、磨蚀和劈裂等物理伤害;另一方面是风蚀带走地表沙土使植物根系暴露, 甚至使其丧失固着能力。在发生风沙活动的地区沙埋、风蚀和风沙流磨蚀是同时存在的, 目前有关沙埋对植物生长和生理的影响研究较多, 而风沙流对植物地上部分机械性损伤以及风蚀导致植物根系暴露后生长及生理响应的研究较少。大量研究发现沙埋后植物生理指标表现为, 植物体内活性氧升高, 致超氧化物歧化酶(SOD, Superoxide Dismutase)和过氧化物酶(POD, Peroxidase)活性增强, 并及时清除过氧化物, 保护细胞膜不被氧化;渗透调节系统中的脯氨酸含量也升高, 降低细胞的渗透势, 防止细胞脱水;植物生产低于消耗, 作为储存物质的可溶性糖被分解利用维持生长, 含量呈下降趋势[6-12]。沙下叶片遭遇黑暗和缺氧胁迫, 丙二醛(MDA, Malondialdehyde)含量增幅越小代表植物抗沙埋特性越强[13]。与沙埋相反, 风蚀将植物根系大量暴露在地表, 使植物受到高温、水分缺乏和营养匮乏的威胁, 显著降低植物叶水势和气孔导度, 抑制光合作用, 限制植物的生长[14-16], 另一方面风蚀带走沙土从而改变了植物的生长环境, 进而影响植物的更新和繁殖方式[17]。我们通过野外考察也发现, 大量固沙植物因沙丘活化或处在风蚀强烈区, 致使其根系逐渐裸露, 直至植株死亡。本试验以常见典型固沙植物柠条锦鸡儿和沙拐枣为研究对象, 采样测量主根暴露不同处理深度后三个时间点的各项生理指标, 从而揭示柠条锦鸡儿和沙拐枣遭受风蚀导致根系暴露后的生理动态响应过程及抗逆机理。
1 研究区概况及研究方法宁夏灵武白芨滩国家级自然保护区(E106°20′22″—106°37′19″, N37°49′05″—38°20′54″)位于我国西部鄂尔多斯台地西南缘, 常见人工固沙植物有柠条锦鸡儿、沙拐枣、花棒(Hedysarum scoparium)等。气候属中温干旱气候区, 具有典型的大陆性气候特征, 全年日照时数3080 h, 平均无霜期157 d, 植物生长期持续170 d, 年平均≥10℃积温3351.3℃, 年平均气温8.8℃, 年均降水量206.2—255.2 mm。年平均风速5.1 m/s, 东南风较多, 西北西南风次之, 4级风天数70.9 d, 冬春季风沙多。保护区南部以沙地丘陵为主, 北部以山地荒漠为主, 最高海拔为1650 m。
于2018年4月14日, 在保护区南部的固定沙丘迎风侧(西北朝向), 选取以8年生的柠条锦鸡儿和沙拐枣为建群种的样地。沙拐枣平均株高1.5 m, 处在展叶期, 柠条锦鸡儿平均株高2 m, 处在开花期。两个植物种各选取长势相近的12株作为供试材料, 供试材料中的3株为正常生长的对照, 剩余9株分成3组进行主根暴露处理, 3组的暴露深度分别为10、20、30 cm, 每组3个重复。考虑到不同植物种和各植株间的侧根根系生长均有差异, 因此尽可能只暴露主根(挑选主根暴露段上侧根较少的植株), 揭示仅主根暴露后对植株生理的影响。首先用记号笔在供试植株主根的土表处做一记号, 之后人为用手将主根周围的沙土(面积约10 cm×10 cm)扒掉使其暴露至处理深度, 在处理后的土表处再做标记。处理后每隔7d采集一次叶片, 共采集3次。采集时将植株按东西南北四个方向分为四部分, 每个方向上在植株的上、中、下采集叶片, 采集后的叶片混合均匀后用锡箔纸包住放入盛有干冰的保温桶内带回实验室, 放入-80℃的超低温冰箱保存待测。
采用分光光度法测叶绿素含量, 茚三酮法测脯氨酸含量, 硫代巴比诺酸显色法, 蒽酮法测可溶性糖含量, 愈创木酚法测过氧化物酶(POD)活性, NBT还原法测SOD活性, 紫外吸收法测过氧化氢酶(CAT)的活性[18]。
各生理指标值变化率的计算公式:
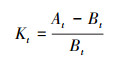
|
式中, Kt为植物处理第t天的某一生理指标的变化率;At为第t天处理组的某一生理指标的平均值, Bt为第t天对照组的某一生理指标的平均值。
采用Microsoft Excel 2010进行数据处理, 以及基础统计分析和绘图;采用SPSS18.0进行单因素方差分析。
2 结果与分析 2.1 叶绿素含量及叶绿素(a/b)对根系暴露深度的响应柠条锦鸡儿在3种不同根系暴露深度(10、20、30 cm)处理下, 其叶绿素含量及Chl(a/b)相较对照组的变化如图 1、2所示。柠条锦鸡儿在根系暴露10 cm深度后的3个观测时间点(7、14、21 d), 其叶绿素含量和Chl(a/b)与对照组相比均无显著性差异;20 cm深度处理下随暴露时间延长叶绿素含量和Chl(a/b)先低于对照组后高于对照组(叶绿素含量高于对照时有显著差异(P < 0.05, 下同), Chl(a/b)低于对照时有显著差异);30 cm深度处理下第7天叶绿素含量和Chl(a/b)均显著低于对照组, 第14天时处理组与对照组间的叶绿素含量和Chl(a/b)差异逐渐减小(叶绿素含量已无显著差异, Chl(a/b)仍显著低于对照), 第21天叶绿素含量和Chl(a/b)与对照组间均无显著差异。
如图 1、2所示, 沙拐枣根系暴露10 cm深度处理下, 3个观测期的叶绿素含量与对照间均无显著性差异, 20、30cm深度处理在第7天与对照间无显著性差异, 第14天和第21天均与对照间有显著性差异;沙拐枣3个深度处理的Chl(a/b)在根系暴露至第7天和第14天时与对照间均存在显著性差异(第7天显著高于对照, 第14天时20 cm和30 cm处理显著低于对照), 至第21天时3个处理与对照间均无显著性差异。

|
| 图 1 不同根系暴露深度下柠条锦鸡儿和沙拐枣的叶绿素含量及其变化率 Fig. 1 Chlorophyll content and its change rate of Caragana korshinskii and Calligonum mongolicum at different root exposure depth 相同的字母表示各处理之间差异不显著; 不同的字母表示差异显著(P < 0.05) |

|
| 图 2 不同根系暴露深度下柠条锦鸡儿和沙拐枣的Chl(a/b)及其变化率 Fig. 2 Chl(a/b) and its change rate of Caragana korshinskii and Calligonum mongolicum at different root exposure depth |
如图 3所示, 柠条锦鸡儿根系暴露10、20 cm深度的MDA含量在观测期内与对照间均无显著差异, 30 cm深度的MDA含量在第7天、第14天低于对照值(P < 0.05), 第21天恢复至与对照无显著差异;沙拐枣3个处理的MDA含量在观测期内均与对照间无显著性差异。

|
| 图 3 不同根系暴露深度下柠条锦鸡儿和沙拐枣的丙二醛(MDA)及其变化率 Fig. 3 Malondialdehyde (MDA) and its change rate of Caragana korshinskii and Calligonum mongolicum at different root exposure depth |
如图 4所示, 柠条锦鸡儿3个处理的可溶性糖含量在观测期内均与对照间无显著性差异。沙拐枣3个处理的可溶性糖含量在第7天均与对照间存在显著性差异(P < 0.05)(各处理间无显著差异), 第14天、21天则与对照间无显著性差异。综上, 30cm深度内的根系暴露处理, 对柠条锦鸡儿的可溶性糖含量无影响, 沙拐枣的可溶性糖含量随暴露时间增长表现为先上升后恢复至对照水平。

|
| 图 4 不同根系暴露深度下柠条锦鸡儿和沙拐枣的可溶性糖含量及其变化率 Fig. 4 Soluble sugar content and its change rate of Caragana korshinskii and Calligonum mongolicum at different root exposure depth |
如图 5所示, 柠条锦鸡儿30 cm深度处理的Pro含量仅在第7天相比对照显著升高(P < 0.05), 至第21天各处理间及其与对照间均无显著性差异;10 cm深度处理下的沙拐枣Pro含量在第21天时相比对照显著升高, 其他处理在观测期内相比对照均无显著性差异或未显著上升。

|
| 图 5 不同根系暴露深度下柠条锦鸡儿和沙拐枣的脯氨酸(Pro)含量及其变化率 Fig. 5 Proline (Pro) content and its change rate of Caragana korshinskii and Calligonum mongolicum at different root exposure depth |
如图 6、图 7所示, 柠条锦鸡儿3个深度处理在观测期内的SOD活性均与对照间无显著性差异, 20、30 cm两个处理的CAT活性在第7天和14天与对照存在显著性差异(P < 0.05);沙拐枣10 cm深度处理的SOD活性和柠条锦鸡儿10 cm深度处理的CAT活性在观测期也均与对照间无显著性差异;沙拐枣20、30 cm两个处理下, SOD活性在第14天和21天, CAT活性在观测期内均与对照间存在显著性差异(P < 0.05)。综上, 柠条锦鸡儿的SOD活性对3个深度处理均无显著响应, CAT活性先升后降, 在处理21 d后也恢复至对照水平;沙拐枣在20、30 cm两个处理下, SOD活性和CAT活性至第21天时仍显著低于对照(P < 0.05), 10 cm深度处理的SOD和CAT活性与对照间基本无显著差异。

|
| 图 6 不同根系暴露深度下柠条锦鸡儿和沙拐枣的超氧化歧化酶(SOD)及其变化率 Fig. 6 Superoxide dismutase (SOD) and its change rate of Caragana korshinskii and Calligonum mongolicum at different root exposure depth |

|
| 图 7 不同根系暴露深度下柠条锦鸡儿和沙拐枣的过氧化氢酶(CAT)及其变化率 Fig. 7 Catalase (CAT) and its change rate of Caragana korshinskii and Calligonum mongolicum at different root exposure depth |
与其他单因素对植物造成的逆境胁迫不同, 风蚀导致植物根系大量暴露在地表, 使其可能受到高温、水分和营养缺乏等多因素共同作用的胁迫。沙拐枣遭受风蚀致根系出露后的母株和分株的生长和繁殖均受到影响, 其母株和分株的叶绿素含量、净光合速率、水分利用效率和同化枝长度、数量等只有沙埋分株的50%左右[19]。超氧化物歧化酶(SOD), 过氧化物酶(POD)和过氧化氢酶(CAT)是抗氧化酶系统的重要组成, SOD催化氧自由基转变为H2O2, POD与CAT则一起分解由SOD作用产生的H2O2, 生成无害的H2O, 3种酶相互协调能有效清除代谢过程产生的活性氧[20-21]。有研究表明植物在轻度干旱胁迫下, SOD、POD、CAT活性会上升, 但在中重度干旱胁迫下, 其活性则会下降[22-24], 同时干旱胁迫会抑制叶绿素的生物合成并促使其降解[25]。可溶性糖和脯氨酸是植物组织内理想的渗透调节物, 当其含量大幅度增加时, 细胞渗透势降低, 植物组织的持水能力提高, 维持体内水分, 保护植物生理稳态[26]。叶绿素a的含量与土壤水分呈正相关, 相同的干旱胁迫对叶绿素a比叶绿素b影响大[27]。本实验中的沙拐枣在根系暴露20—30 cm深度的后两周, 其SOD、CAT活性和叶素绿含量相比对照均显著下降, MDA含量相比对照有所升高, 加之观测期是在沙丘土壤水分含量整体偏少的春季, 由此推断在春季干旱期根系暴露20 cm以上即对沙拐枣的抗氧化酶系统和光合作用造成影响, 使其处于中重度的干旱胁迫状态, 但沙拐枣根系暴露20 cm以内的处理第一周, 其SOD、CAT活性、Pro含量、叶绿素含量与对照间无显著差异, 而其可溶性糖含量、Chl(a/b)相比对照显著升高, 表明沙拐枣在此阶段通过渗透调节功能和提高光能利用率来维持生理稳态[14-17]。综上判断春季干旱期的沙拐枣成株, 根系暴露深度和暴露时长均是决定其受胁迫程度的重要影响因素, 根系暴露后的多因素作用中, 以干旱胁迫作用为主。
3.2 柠条锦鸡儿主根暴露后的抗逆机理柠条锦鸡儿根系暴露10 cm深度所测的各项生理指标在观测期内与对照间均无显著差异, 意味着根系暴露10 cm深度对柠条锦鸡儿未造成任何环境胁迫。MDA是膜脂过氧化作用的最终产物[28], 当植物处于干旱、高温、盐碱等逆环境胁迫时, 细胞内活性氧的产生与消除的平衡就会受到破坏, 导致活性氧大量积累, 从而引发膜脂过氧化作用, 造成MDA大量积累[29]。柠条锦鸡儿根系暴露30 cm深度时其MDA含量在前两周相比对照显著下降, 这与风蚀沙埋后一些沙生植物幼苗MDA含量的变化趋势一致[13-14, 28], 表明根系暴露后的逆境胁迫, 未引发膜脂过氧化。柠条锦鸡儿的叶绿素含量和Chl(a/b)在前期均存在不同程度下降, 此结果与马洋等发现风蚀导致幼苗叶绿素含量降低的结果一致[30], 植物在受干旱胁迫时叶绿素b的波动较小[31], 因此推测叶绿素含量及Chl(a/b)的下降是由叶绿素a合成变缓或分解过快所致, 但随着暴露时间的延长, 其叶绿素含量及Chl(a/b)均恢复至对照水平。有研究表明弱干旱胁迫时CAT活性对植物抗性起主要作用, 而强干旱胁迫时SOD和POD起主要作用[32], 本研究中20 cm和30 cm深度处理的柠条锦鸡儿CAT活性和Pro含量在处理后7 d内显著升高, 21 d后又恢复至对照水平, 而SOD活性始终与对照间无差异, 因此判断CAT活性增强和以游离脯氨酸为主的渗透调节系统共同作用使其逐渐适应根系暴露所致的环境胁迫。综上, 20—30 cm深度根系的出露造成柠条锦鸡儿受轻度胁迫, 但因柠条锦鸡儿根系主要分布在60 cm层内, 且其根系生物量密度显著大于其他植物种, 获取水分和养分的能力较强[33], 因此主根暴露30 cm深度对其并不能造成不可修复的生理损伤。
4 结论在春季干旱期, 不同物候期的柠条锦鸡儿和沙拐枣根系暴露30 cm深度, 两者的生理响应过程和适应机理存在明显的差异性。
(1) 根系暴露10 cm深度对开花期柠条锦鸡儿未造成胁迫, 20—30 cm深度的根系暴露, 其通过CAT活性快速升高和游离脯氨酸为主的渗透调节系统的共同作用, 从而在21 d后恢复至正常水平。
(2) 沙拐枣根系暴露20 cm深度以内的初期, 其通过积累可溶性糖及提高Chl(a/b)来维持生理稳态, 根系暴露20 cm以上的两周后削弱了其抗氧化酶系统和光合作用, 使其处于中重度的干旱胁迫状态, 且在21 d后仍未恢复至对照水平, 根系暴露深度和暴露时长协同决定其受胁迫的程度。
| [1] |
Barchyn T E, Hugenholtz C H. Winter variability of aeolian sediment transport threshold on a cold-climate dune. Geomorphology, 2012, 177-178: 38-50. DOI:10.1016/j.geomorph.2012.07.012 |
| [2] |
赵哈林. 沙漠生态学. 北京: 科学出版社, 2012.
|
| [3] |
杨小林, 张希明, 李义玲, 张永领. 基于分形理论的塔克拉玛干沙漠腹地自然植物根系构型特征分析. 干旱区资源与环境, 2015, 29(8): 145-150. |
| [4] |
石坤, 贾志清, 张洪江, 张友焱, 李清雪, 冯莉莉, 何凌仙子. 青海共和盆地典型固沙植物根系分布特征. 中国水土保持科学, 2016, 14(6): 78-85. |
| [5] |
郑士光, 贾黎明, 庞琪伟, 李锐. 平茬对柠条林地根系数量和分布的影响. 北京林业大学学报, 2010, 32(3): 64-69. |
| [6] |
赵哈林, 曲浩, 周瑞莲, 云建英, 李瑾, 王进. 沙埋对小麦(Triticum aestivum)生长的影响及其生理响应. 中国沙漠, 2014, 34(3): 689-695. |
| [7] |
赵哈林, 曲浩, 赵学勇, 周瑞莲, 云建英, 李瑾, 王进. 差巴嘎蒿幼苗对沙埋的生态适应和生理响应. 生态学报, 2014, 34(20): 5832-5839. |
| [8] |
王进, 周瑞莲, 赵哈林, 赵彦宏, 侯玉萍. 海滨沙地砂引草对沙埋的生长和生理适应对策. 生态学报, 2012, 32(14): 4291-4299. |
| [9] |
周瑞莲, 王进, 杨淑琴, 杨树德. 海滨沙滩单叶蔓荆对沙埋的生理响应特征. 生态学报, 2013, 33(6): 1973-1981. |
| [10] |
解卫海, 周瑞莲, 梁慧敏, 曲浩, 董龙伟, 强生斌. 海岸和内陆沙地砂引草(Messerschmidia sibirica)对自然环境和沙埋处理适应的生理差异. 中国沙漠, 2015, 35(6): 1538-1548. |
| [11] |
周瑞莲, 杨淑琴, 黄清荣, 左进城, 陈娟丽, 李赞. 小叶锦鸡儿抗沙埋生长与抗氧化酶及同工酶变化的关系. 生态学报, 2015, 35(9): 3014-3022. |
| [12] |
高洁, 李从娟, 徐新文, 孙永强. 罗布泊地区引种植物生理生化指标的主成分及其聚类分析. 干旱区研究, 2018, 35(1): 199-206. |
| [13] |
赵哈林, 曲浩, 周瑞莲, 李瑾, 潘成臣, 王进. 沙埋对两种沙生植物幼苗生长的影响及其生理响应差异. 植物生态学报, 2013, 37(9): 830-838. |
| [14] |
樊宝丽, 马全林, 郭树江, 张剑挥, 张晓娟, 张卫星, 杜鹃, 尉秋实, 赵长明. 克隆植物沙拐枣的母株和分株对风蚀沙埋的生理生态响应. 西北植物学报, 2016, 36(12): 2491-2497. |
| [15] |
马洋, 王雪芹, 张波, 刘进辉, 韩章勇, 唐钢梁. 风蚀和沙埋对塔克拉玛干沙漠南缘骆驼刺水分和光合作用的影响. 植物生态学报, 2014, 38(5): 491-498. |
| [16] |
杨显基, 杜建会, 秦晶, 陈志华, 杨林, 林萍. 海岸沙生植物老鼠艻(Spinifex littoreus)叶水势对风蚀、沙埋和风沙流磨蚀的响应. 生态学杂志, 2017, 36(5): 1215-1223. |
| [17] |
张萍, 哈斯, 王帅, 张素红. 呼伦贝尔沙质草原风蚀坑积沙区的植被分带性. 自然资源学报, 2008, 23(2): 237-244. DOI:10.3321/j.issn:1000-3037.2008.02.008 |
| [18] |
王学奎. 植物生理生化实验原理和技术 (第二版). 北京: 高等教育出版社, 2006.
|
| [19] |
樊宝丽. 风沙环境下沙拐枣(Calligonum mongolicum)自然更新策略研究. 兰州: 兰州大学, 2017.
|
| [20] |
王沙生, 高荣孚, 吴贯明. 植物生理学 (第二版). 北京: 中国林业出版社, 1991: 87-89.
|
| [21] |
丁龙, 赵慧敏, 曾文静, 李晴, 汪洋, 王四清. 五种西北旱区植物对干旱胁迫的生理响应. 应用生态学报, 2017, 28(5): 1455-1463. |
| [22] |
朱志梅, 杨持. 沙漠化过程中四个共有种的生长和抗氧化系统酶类变化. 应用生态学报, 2004, 15(12): 2261-2266. DOI:10.3321/j.issn:1001-9332.2004.12.013 |
| [23] |
罗青红, 宁虎森, 何苗, 吉小敏, 雷春英. 5种沙地灌木对干旱胁迫的生理生态响应. 林业科学, 2017, 53(11): 29-42. DOI:10.11707/j.1001-7488.20171104 |
| [24] |
潘航, 冯缨, 王喜勇, 李岩. 荒漠环境下10种沙拐枣的生理特征比较研究. 草业学报, 2017, 26(6): 68-75. |
| [25] |
李合生. 现代植物生理学. 北京: 高等教育出版社, 2002.
|
| [26] |
马剑, 刘贤德, 孟好军, 赵维俊, 王荣新, 王艺林, 任小凤. 水分胁迫对文冠果幼苗生长及生理特性的影响. 干旱区资源与环境, 2018, 32(1): 128-132. |
| [27] |
麻云霞, 李钢铁, 张宏武, 蓝登明, 姚庆智, 冯斐. 酸枣幼苗光合特征和生理生化指标对不同强度干旱胁迫的响应. 干旱区资源与环境, 2018, 32(12): 164-169. |
| [28] |
Chaitanya K V, Sundar D, Masilamani S, Reddy A R. Variation in heat stress-induced antioxidant enzyme activities among three mulberry cultivars. Plant Growth Regulation, 2002, 36(2): 175-180. DOI:10.1023/A:1015092628374 |
| [29] |
Simon E W. Phospholipids and plant membrane permeability. New Phytologist, 1974, 73(3): 377-420. DOI:10.1111/j.1469-8137.1974.tb02118.x |
| [30] |
马洋, 王雪芹, 韩章勇, 刘进辉. 风蚀沙埋对疏叶骆驼刺(Alhagi sparsifolia)和花花柴(Karelinia caspica)幼苗的生理影响. 中国沙漠, 2015, 35(5): 1254-1261. |
| [31] |
马雪梅, 吴朝峰. 干旱胁迫对金银花叶片叶绿素含量及荧光特性的影响. 江苏农业科学, 2018, 46(17): 133-136. |
| [32] |
康太. 光呼吸代谢和抗氧化系统对柠条锦鸡儿干旱适应性的作用研究. 咸阳: 西北农林科技大学, 2016.
|
| [33] |
何凌仙子. 青海共和盆地典型固沙植物根系特征及功能研究. 北京: 中国林业科学研究院, 2018.
|