 2021, Vol. 41
2021, Vol. 41文章信息
- 宋永鹏, 唐希颖, 贾亚敏, 刘小燕, 崔耀平
- SONG Yongpeng, TANG Xiying, JIA Yamin, LIU Xiaoyan, CUI Yaoping
- 甘南地区二氧化碳施肥效应对生态系统的影响
- Fertilization effect of carbon dioxide on ecosystems in Gannan, China
- 生态学报. 2021, 41(18): 7301-7311
- Acta Ecologica Sinica. 2021, 41(18): 7301-7311
- http://dx.doi.org/10.5846/stxb202005091163
-
文章历史
- 收稿日期: 2020-05-09
- 修订日期: 2021-08-21






2. 中国林业科学研究院湿地研究所, 北京 100091;
3. 太原理工大学水利科学与工程学院, 太原 030000;
4. 黄河中下游数字地理技术教育部重点实验室(河南大学), 开封 475004
2. Institute of Wetland Research, Chinese Academy of Forestry, Beijing 100091, China;
3. School of Hydro Science and Engineering, Taiyuan University of Technology, Taiyuan 030000, China;
4. Key Laboratory of Geospatial Technology for the Middle and Lower Yellow River Regions (Henan University), Ministry of Education, Kaifeng 475004, China
陆地生态系统对地球碳循环非常重要[1]。定量评估陆地生态系统的碳收支情况是进行区域气候调节、碳汇管理的重要依据[2], 也是目前在全球气候变化研究中较为前沿和热门的前沿领域[3-4]。植被在碳循环中扮演着极为重要的角色, 植被通过光合作用储存的碳量是陆地生态系统碳库的重要组成部分。其中, 在森林、草地和农田中储存的碳量占陆地生态系统碳库的27.2%[5], 对全球碳收支具有及其重要的意义, 对全球碳循环和碳平衡也起着重要作用[6]。陆地生态系统总初级生产力(Gross primary productivity, GPP)是指植被通过光合作用等一系列生物活动, 将大气中的CO2吸收同化到自身体内, 从而转化累计的有机物[7];净初级生产力(Net primary productivity, NPP)是植被通过自养消耗剩余的有机碳量[8], 它反映了生态功能状况[9-11], 是生物地球化学碳循环的关键因素[12-13], 也是量化气候变化和人类活动对陆地植被综合影响的重要指标之一[14];净生态系统生产力(Net ecosystem productivity, NEP)是植被经历过自养呼吸后, 再次经历异养消耗而剩余的碳量净贮存。当NEP>0时, 表明生态系统为碳汇, 反之则为碳源[15-16]。
目前, 有大量的研究证明植被GPP和NPP能够被用来很好地量化生态系统的碳汇/源过程[17], 也有很多研究采用GPP和NPP来研究CO2的施肥效应, 例如Terrer等提出CO2的增长可以直接为植被的固碳量带来施肥效应[18];Girardin等通过研究发现CO2含量增长一倍可以促进NPP增长23%[19]。但是, 利用GPP和NPP来研究评估生态系统固碳能力, 没有充分考虑植被异养的呼吸消耗, 而从碳收支原理上来看, 采用NEP来评估生态系统固碳能力是更准确和更有价值的, 然而, 利用NEP来评价区域植被固碳量的成果目前还是相对不足。此外, 在分析CO2对植被固碳能力的影响机制中, 有研究发现, CO2浓度的升高会持续增加森林的碳汇[20];也有研究指出提出CO2浓度的增加有利于NPP的积累[21]。但目前就CO2对植被固碳能力的反馈效应的研究相对较少, 相关案例支撑也相对不足。
基于以上研究背景, 本研究利用遥感数据, 采用光能利用率模型C-FIX, 来模拟分析2019甘南地区的碳循环, 包括GPP、NPP和NEP, 对比三者关系;并利用敏感性实验揭示CO2施肥效应因子的有无对碳循环的影响。
1 数据与方法 1.1 研究区概况甘南地区(100°46′E—104°44′E, 33°06′N—36°10′N), 位于中国甘肃省西南部, 地处青藏高原东北边缘与黄土高原西部之间, 东部和北部与陇南市、临夏州、定西市毗连, 南与四川阿坝州相接, 西南与青海黄南州、果洛州接壤, 整体板块呈现出西北高, 东南低的地势特点, 其总面积大致为38521 km2。甘南地区具有大陆性季节气候的特点, 年平均气温在1—13℃之间, 地域差异很大, 总体温度分布趋势呈自东南向西北逐渐递减(图 1)。

|
| 图 1 甘南地区 Fig. 1 Study area: Gannan |
太阳辐射数据。太阳辐射数据是来自韩国国立首尔大学环境生态实验室的全球陆表下行短波辐射产品, 其原始时间分辨率为月, 空间分辨率为0.05°×0.05°。本研究假设一定范围内的太阳辐射基本一致, 在具体分析时对该数据的空间分辨率重采样为30 m×30 m。
温度数据。该数据原始值来自于国家气象信息中心的全国国家级台站(基本、基准和一般站)的气温日值观测资料。本研究利用30 m×30 m的DEM数据通过ANUSPLIN [22]插值将其处理为栅格温度数据。
归一化植被指数数据(NDVI)。该数据基于Google Earth Engine遥感大数据云计算平台, 由2019年所有可用的Landsat 8红波段和近红外波段数据通过去云等处理之后在归一化, 生成NDVI的月数据。其时间分辨率为月, 空间分辨率为30 m×30 m。
1.3 研究方法 1.3.1 碳循环遥感模型模拟基于Monteith理论, Veroustraete等建立了光能利用率模型C-FIX[23]。该模型可以在区域尺度上实现对GPP、NPP及NEP三个基本碳循环参数的估算, 并对地理空间的每个栅格像元进行以日为单位的碳循环量化。本研究转换至月的计算, 并进行季与年的实际尺度分析。模型公式表示如下:
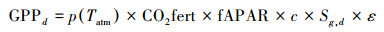
|
(1) |
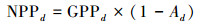
|
(2) |
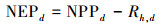
|
(3) |
其中, 下标d代表日值。p(Tatm)是归一化气温依赖因子;CO2fert是归一化CO2施肥效应因子;fAPAR是植被光合作用可吸收的有效辐射比例;c为气候效率因子, 取值为0.48;Sg, d为地面辐射日总量, 其单位是MJ m-2d-1;ε是光能利用率, 取值为1.10 gC/MJ;Ad是植被自养呼吸率;Rh, d是土壤异养呼吸通量, 其单位为gC m-2d-1。其中, 归一化CO2施肥效应因子及其余各因子的公式表示为:
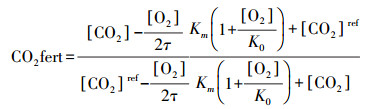
|
(4) |
式中, [CO2]代表目前测定的大气CO2混合浓度, 取值为410 mg/m3;[O2]代表目前测定的大气O2混合浓度, 取值为20.90 mg/m3;τ为CO2对O2的浓度比率;Km为Rubisco与CO2亲和力常数;K0为O2的阻止力常数;[CO2]ref是基准大气中CO2混合浓度, 取值为工业革命前期的浓度值281。
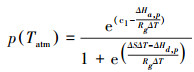
|
(5) |
式中, c1为常数21.77;∆Hd, p为惰性分子能量, 取值为211000 J/mol;Rh, d是土壤一样呼吸通量, 其单位为gC m-2d-1;∆S是CO2动态熵平衡, 取值为704.98 J K-1mol-1;T代表甘南地区日平均气温;Rg为普适气体常数, 其值为8.31 J K-1mol-1。
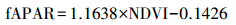
|
(6) |
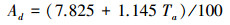
|
(7) |
其中, Ta为日平均气温, 单位是℃。
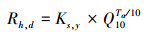
|
(8) |
其中, Ks, y是全年日平均异养呼吸效率, 其单位是gC m-2d-1;Q10代表温度每升高10℃, 植物异养呼吸相对增加的倍数。
对于公式中的Km、K0、Ks, y及τ需要进一步用下列公式运算:
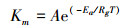
|
(9) |
其中, 当T < 15℃时, Ea= 109600 J/mol, A= 1.976 x 1022;当T ≥ 15℃时, Ea=59400 J/mol, A=2.419×1013。
K0也按照公式(9)计算, 其中A0=8240, Ea0 = -13913.5 J/mol。
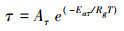
|
(10) |
式中, Aτ=7.87x10-5, Eaτ = -42896.9 J/mol。
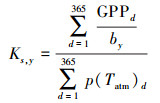
|
(11) |
式中, by是土壤异养消耗年平均标定系数, 通常以距离较近、生态类型相近的通量观测数据标定。
1.3.2 CO2敏感性检验参照C-FIX光能利用率模型, 在去除CO2的条件下, 对甘南地区GPP、NPP及NEP三个基本碳循环参数的模拟运算。具体公式如下:
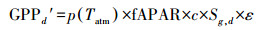
|
(12) |
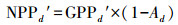
|
(13) |
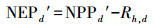
|
(14) |
为模拟CO2对甘南地区植被各碳循环参数的贡献情况, 设立CO2贡献度因子量化CO2对碳循环各因子产生的影响。

|
(15) |
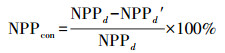
|
(16) |
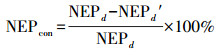
|
(17) |
GPP、NPP及NEP三者的月变化趋势基本一致, 上半年呈持续递增态势, 下半年呈持续递减趋势。由甘南地区2019年GPP、NPP及NEP的月变化趋势图可知, GPP、NPP及NEP的峰值出现在7月, 其值为分别为164.67 gC m-2月-1、121.33 gC m-2月-1和58.19 gC m-2月-1;最小值出现在1月份, 分别为0.82 gC m-2月-1、0.82 gC m-2月-1和-20.76 gC m-2月-1。其中, 对于NEP, 5、6、7、8、9月份, 其值为正, 即植被最终所固定的剩余有机碳量为正;而在其余月份的值为负, 即生态系统呈现碳源作用。
GPP、NPP、NEP随季节变化明显。在本研究时段内, 研究区夏季的GPP、NPP、NEP明显高于其他季节, 其值分别为422.98 gC m-2月-1、315.08 gC m-2月-1和141.32 gC m-2月-1;春季和秋季的GPP、NPP、NEP差异较小, 春秋季的GPP约为夏季的0.28倍, 其NPP约为夏季的0.3倍;最小值出现在冬季, 其值分别为4.55 gC m-2月-1、4.46 gC m-2月-1和-57.38 gC m-2月-1(图 2)。

|
| 图 2 甘南地区2019年GPP、NPP、NEP月趋势变化图和季度统计图 Fig. 2 Monthly trend change and quarterly statistical chart of GPP, NPP and NEP in Gannan in 2019 GPP: 陆地生态系统总初级生产力; NPP: 净初级生产力; NEP: 净生态系统生产力 |
本研究同时对比了甘南地区下辖地区的植被年GPP、NPP、NEP。由2019年甘南地区各下辖县、市年均GPP、NPP、NEP统计表可知, 迭部县、临潭县和舟曲县的GPP较大, 其年均GPP都在840 gC m-2a-1以上;迭部县、临潭县和卓尼县的NPP较大, 其年均NPP都在600 gC m-2a-1以上;临潭县和卓尼县的NEP较大, 其值都在100 gC m-2a-1以上。其中, 临潭县占地面积最小, 占地面积仅为1394.25km2, 但是其GPP和NEP最大, 分别为1050.53 gC m-2a-1和122.59 gC m-2a-1。年均GPP、NPP最小值出现在玛曲县, 分别为465.30 gC m-2a-1和391.47 gC m-2a-1, 其次是碌曲县和夏河县, GPP、NPP分别围绕550 gC m-2a-1和450 gC m-2a-1上下浮动。甘南地区固碳总量为54.91 gC m-2a-1, 其中, 舟曲县的NEP最小, 其值为负, -237.43 gC m-2a-1(图 3)。

|
| 图 3 甘南地区各下辖县2019年的GPP、NPP、NEP和甘南地区年固碳总量 Fig. 3 Annual GPP, NPP, NEP in each county of Gannan and total carbon sequestration in 2019 |
甘南地区2019年GPP、NPP、NEP的州总量分别为31524 tC/a、24310 tC/a和2611 tC/a。其中, GPP和NPP空间分布表现为东部高于西部的显著特征, 东部地区主要是舟曲县、迭部县、卓尼县、临潭县, 西部地区主要是玛曲县、夏河县。NEP空间分布主要表现为北部高于南部的特征, 即较大的NEP主要分布在卓尼县和临潭县, 较小值主要集中在舟曲县(图 4)。对于各下辖县、市的区域年固碳总量(单位: tC/a)进行对比, 得出结果: 卓尼县(811.93)>玛曲县(771.57)>夏河县(659.66)>碌曲县(432.98)>迭部县(338.72)>合作市(272.46)>临潭县(222.42)>舟曲县(-898.81)(图 3)。

|
| 图 4 甘南地区2019年GPP、NPP、NEP空间分布图和总量统计图 Fig. 4 Spatial distribution of GPP, NPP, and NEP and total statistical chart in Gannan in 2019 |
在去除CO2因子影响的条件下, 模拟甘南地区2019年的碳循环情况。由月趋势变化图可知, GPP、NPP、NEP的月变化趋势仍基本一致, 上半年呈持续递增态势, 下半年呈持续递减趋势。GPP、NPP、NEP的最大值均出现在7月, 对应的值分别为131.53 gC m-2a-1、97.76 gC m-2a-1和48.52 gC m-2a-1;最小值出现在1月, 其值分别为0.81 gC m-2a-1、0.81 gC m-2a-1和-18.34 gC m-2a-1。对于NEP, 在5、6、7、8、9月份其值大于0, 当月植被最终的固碳总量为正;而在其余月份NEP都小于0, 即当月区域整体表现为碳源。
去除CO2条件下的GPP、NPP、NEP也具有明显的季节性规律。甘南地区夏季的GPP、NPP、NEP明显高于其他三个季节, 其值分别是340.58 gC m-2a-1、255.93 gC m-2a-1和119.74 gC m-2a-1;春、秋季节相差不大, 其值分别围绕100 gC m-2a-1、82 gC m-2a-1和-7 gC m-2a-1上下浮动, 其中, 春、秋季节的GPP约为夏季总数的0.3倍, NPP约为夏季总数的0.32倍;最小值出现在冬季, 分别为4.46 gC m-2a-1、4.38 gC m-2a-1和-50.65 gC m-2a-1。对于NEP, 除夏季, 其余季节的NEP均为负(图 5)。

|
| 图 5 去除CO2效应下甘南地区2019年GPP、NPP、NEP月趋势变化图和季度统计图 Fig. 5 Monthly trend change and quarterly statistical chart of GPP, NPP and NEP without the effect of CO2 in Gannan in 2019 |
比较研究区各下辖地区去除CO2影响条件下的GPP、NPP及NEP, 可以得到: 甘南地区的临潭县、迭部县及卓尼县的GPP和NPP较大, 其值分别都处于650 gC m-2a-1和500 gC m-2a-1以上, 而玛曲县的GPP和NPP较小, 其值分别低于420 gC m-2a-1和350 gC m-2a-1。对于NEP, 研究区固碳总量是55.07 gC m-2a-1, 其中最大的下辖县是临潭县, 为147.36 gC m-2a-1, 其次是卓尼县和迭部县, 其值分别为138.70 gC m-2a-1和120.76 gC m-2a-1, 而剩余下辖县2019年的总固碳量都小于100 gC m-2a-1, 其中舟曲县的NEP最小, 其值为-138.68 gC m-2a-1(图 6)。

|
| 图 6 去除CO2效应下甘南地区各下辖县2019年年均GPP、NPP、NEP和甘南地区年固碳总量 Fig. 6 Annual GPP, NPP, and NEP in each county and total carbon sequestration in Gannan in 2019 without CO2 |
在没有CO2效应的情况下, 甘南地区2019年GPP、NPP、NEP的州总量分别为26100 tC/a、20283 tC/a、2619 tC/a。从空间分布图中可以看出, 此条件下, 2019年甘南地区年均GPP和NPP表现出南部高于北部, 东部高于西部的特征, 具有较大GPP、NPP的下辖地区主要是卓尼县、迭部县以及临潭县, 而玛曲县、碌曲县和夏河县植被的GPP、NPP较小。对于NEP, 较小的NEP集中分布在舟曲县, 较大的NEP集中分布在卓尼县, 而对于迭部县, 该下辖县小部分区域的NEP呈现为负, 大部分区域的NEP为正, 且数值较大(图 7)。对各下辖区域单位面积的固碳量进行对比(单位: tC/a), 得出其值大小次序: 卓尼县(945.29)>迭部县(730.15)>玛曲县(364.02)>夏河县(343.17)>碌曲县(297.93)>临潭县(267.36)>合作市(204.86)>舟曲县(-525.00)(图 6)。

|
| 图 7 去除CO2效应下甘南地区2019年GPP、NPP、NEP空间分布图和总量统计图 Fig. 7 Spatial distribution of GPP, NPP and NEP and total statistical chart without CO2 in Gannan in 2019 |
将具有CO2条件下和去除CO2条件下的GPP、NPP、NEP空间分布图叠加相减, 量化甘南地区2019年CO2对植被光合作用的影响。由2019年甘南地区GPP差值空间分布图可知, 甘南地区整体GPP的差值为正, 极少像元值被统计出为负数。这表明CO2对植被的总初级生产力具有正向反馈作用, 即对植被进行有机碳总量的固定过程具有施肥效应, 其总贡献率为14.4%;且该效应在甘南地区明显表现为东高西低的特征, 其中, CO2对临潭县植被固碳的施肥效应最为明显, 为242.18 gC m-2a-1, 其贡献率是23.4%, 其次是迭部县和舟曲县, 其施肥效应都在190 gC m-2a-1以上, CO2对玛曲县植被的施肥效应最弱, 为51.99 gC m-2a-1, 贡献率为9.4%。
由NPP差值空间分布图可知, CO2对甘南地区植被NPP同样具有施肥效应, 其总贡献率为14.3%, 在空间分布上呈现东高西低的特点。CO2对NPP的施肥效应整体上要小于对GPP的施肥效应, 在各下辖县、市中, 对植被NPP施肥效应最明显的是临潭县, 为168.24 gC m-2a-1, 其贡献率为22.8%;其次是迭部县、舟曲县和卓尼县, 施肥效应都在100 gC m-2a-1以上, 舟曲县的CO2贡献率也颇高, 为22.3%;对玛曲县的施肥效应较弱, 仅为43.90 gC m-2a-1, 其贡献率为9.2%。
CO2对NEP的施肥效应在空间上大致表现为北高南低的特征。其中, CO2对NEP存在正向施肥效应的下辖县区主要是夏河县、玛曲县、碌曲县和合作市, 其施肥效应大致围绕30 gC m-2a-1上下浮动, CO2对夏河县和玛曲县NEP的贡献率最大, 接近于50%;CO2对NEP存在负向反馈效应的下辖县区主要是舟曲县、卓尼县、临潭县和迭部县, 其中, 舟曲县的负反馈效应最大, 为-98.75 gC m-2a-1, 其次是迭部县, 负反馈效应为-64.76 gC m-2a-1, 其贡献率为-115.6%(图 8)。

|
| 图 8 有无CO2因子下甘南地区GPP、NPP、NEP差值空间分布图和统计图 Fig. 8 Spatial distribution and statistical charts of difference values of GPP, NPP, and NEP with and without CO2 factor in Gannan |
本研究求算出甘南地区2019年单位面积内GPP年总量为662.88 gC m-2a-1, 同MODIS发布的2019年GPP产品的结果在空间、时间与具体值上(642.95 gC m-2a-1)保持基本一致, 其R2达到0.95, 但本研究GPP受益于分辨率而可以体现出更精细的空间特征。本研究的结果还证实了CO2施肥效应对提升GPP和NPP具有重要作用, 这与之前研究结论也较为一致[24-25]。另外, 本研究结果显示, 就整个碳循环过程来说, 在一些区域CO2并非总是起到施肥效应, 会出现NEP差值为负的现象, 这其实是由于土壤呼吸的变化所导致, 而土壤呼吸是陆地生态系统异养呼吸的重要部分[26]。有研究表明大气中CO2的升高可以促使植被根系微生物活动能力的增强[27], 使得土壤中微生物呼吸释放的CO2增大, 进而抬升了陆地生态系统的异养消耗[28-29]。当陆地生态系统异养消耗的CO2大于CO2对NEP的施肥效应时, 会出现NEP值为负的现象(图 9)。

|
| 图 9 甘南地区GPP与MODIS GPP产品年均值空间对比和甘南地区2019年NEP和温度的生态地理区划分布 Fig. 9 Comparison of annual average value of GPP between MODIS GPP product and C-FIX result and Spatial distribution of ecosystem division and temperature in Gannan |
甘南地区东南部的草地植被覆盖度低于中西部和西南部地区[30], 有学者在研究青藏高原草地碳源/汇时空变化及其与气候因子的关系时, 发现由于温度增长会引起土壤温度的升高[31-32], 进而加大了以异养呼吸为主要形式的土壤呼吸[26], 使得草地生态系统NEP与温度呈负相关性[2]。在本研究区中, 地跨北亚热带、高原温带的区域年均温度较高, 分别为9.4℃和3.9℃, 而位于暖温带和高原亚寒带的区域年均温度较低, 分别是2.4℃和1.8℃, 这也在一定程度上解释了地处甘南地区自治州东南部地区温带、亚热带植被年均固碳量呈现为负的现象(图 9)。
CO2与其他因子相互作用, 改变CO2的同时, 其他参数也会发生变化。比如CO2的变化本身可能已经体现在NDVI的变化上了, 单纯去除CO2其实仅为一个敏感性试验模拟, 而复杂的过程模型对分析机理尤为重要, 这也是本研究需要继续探讨的方向[33-34]。但是, 纵然是复杂如地球系统模型(Earth system model, ESM), 也有研究揭示出因模型对于CO2过度敏感及对这种施肥效应缺乏条件约束而可能严重高估了CO2的施肥效应[35]。从研究区来说, 甘南地区的温湿度、植被类型以及人类活动等都是影响碳循环的重要因素[36]。可以证实的是, 在不同的地理环境下, CO2在碳循环过程中的作用有很大的差异性, 因而, 不能简单认为CO2只单方向起施肥效应。
4 结论本文利用C-FIX光能利用率模型模拟分析了甘南地区植被的GPP、NPP和NEP月变化及空间分布特征,并探究了CO2对GPP、NPP和NEP的施肥效应存在与否。研究结果表明甘南地区2019年植被固碳总量约为2611 tC/a。由于研究区跨生态地理分区较多, 受温度、降水以及地表植被种类多元化的影响, 使得GPP、NPP在空间分布上表现为东高西低的显著特征, NEP呈现出北高南低的特征;其中, 卓尼县是甘南地区植被年固碳量最大的县, 其固碳量为811.93 tC/a, 唯舟曲县为碳源, 其固碳量为-898.81 tC/a。通过分析CO2的敏感性, 发现CO2对GPP和NPP存在正向的施肥效应, 使其分别增加了14.4%、14.3%;而对于NEP, CO2对夏河县、玛曲县、碌曲县和合作市存在正向施肥效应, 而舟曲县、卓尼县、临潭县和迭部县存在负向施肥效应, 整体而言, CO2对NEP的贡献率微乎其微。这说明在不同温湿度等气候背景的区域, CO2是否起到施肥作用更不能一概而论。
本研究从生态地理分区角度做了探讨, 得出不同区域气候因子可能会对植被固碳造成影响, 这对揭示陆地生态系统碳循环的动态机制提供了一定的理论依据。
| [1] |
Tian H Q, Lu C Q, Ciais P, Michalak A M, Canadell J G, Saikawa E, Huntzinger D N, Gurney K R, Sitch S, Zhang B W, Yang J, Bousquet P, Bruhwiler L, Chen G S, Dlugokencky E, Friedlingstein P, Melillo J, Pan S F, Poulter B, Prinn R, Saunois M, Schwalm C R, Wofsy S C. The terrestrial biosphere as a net source of greenhouse gases to the atmosphere. Nature, 2016, 531(7593): 225-228. DOI:10.1038/nature16946 |
| [2] |
周夏飞, 於方, 曹国志, 杨威杉, 周颖. 2001-2015年青藏高原草地碳源/汇时空变化及其与气候因子的关系. 水土保持研究, 2019, 26(1): 76-81. |
| [3] |
Ciais P, Peylin P, Bousquet P. Regional biospheric carbon fluxes as inferred from atmospheric CO2 measurements. Ecological Applications, 2000, 10(6): 1574-1589. |
| [4] |
Li N, Cui Y P, Fu Y M, Liu X Y, Run Y D, Li M D, Chen L Y, Xia H M, Lu H L. Contribution of anthropogenic CO2 in China to global radiative forcing and its offset by the ecosystem during 2000-2015. Annals of the New York Academy of Sciences, 2021, 1488(1): 56-66. DOI:10.1111/nyas.14505 |
| [5] |
王旭峰. 黑河流域陆地生态系统生产力模拟研究[D]. 兰州: 中国科学院寒区旱区环境与工程研究所, 2012: 2-8.
|
| [6] |
胡海清, 魏书精, 孙龙, 王明玉. 气候变化、火干扰与生态系统碳循环. 干旱区地理, 2013, 36(1): 57-75. |
| [7] |
Woodwell G M, Whittaker R H, Reiners W A, Likens G E, Delwiche C C, Botkin D B. Where has all the carbon gone?. Science, 1979, 204(4399): 1346-1346. DOI:10.1126/science.204.4399.1346 |
| [8] |
刘晓雪, 张丽娟, 李文亮, 张学珍, 姜春艳. 中国陆地植被氧气生产量变化模拟及其影响因素. 生态学报, 2015, 35(13): 4314-4325. |
| [9] |
Melillo J M, McGuire A D, Kicklighter D W, Moore B, Vorosmarty C J, Schloss A L. Global climate change and terrestrial net primary production. Nature, 1993, 363(6426): 234-240. DOI:10.1038/363234a0 |
| [10] |
Running S W, Nemani R R, Heinsch F A, Zhao M S, Reeves M, Hashimoto H. A continuous satellite-derived measure of global terrestrial primary production. Bioscience, 2004, 54(6): 547-560. DOI:10.1641/0006-3568(2004)054[0547:ACSMOG]2.0.CO;2 |
| [11] |
Shvidenko A Z, Schepashchenko D G, Vaganov E A, Nilsson S. Net primary production of forest ecosystems of Russia: a new estimate. Doklady Earth Sciences, 2008, 421(2): 1009-1012. DOI:10.1134/S1028334X08060330 |
| [12] |
Crabtree R, Potter C, Mullen R, Sheldon J, Huang S L, Harmsen J, Rodman A, Jean C. A modeling and spatio-temporal analysis framework for monitoring environmental change using NPP as an ecosystem indicator. Remote Sensing of Environment, 2009, 113(7): 1486-1496. DOI:10.1016/j.rse.2008.12.014 |
| [13] |
Zhao M S, Running S W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science, 2010, 329(5994): 940-943. DOI:10.1126/science.1192666 |
| [14] |
张镱锂, 祁威, 周才平, 丁明军, 刘林山, 高俊刚, 摆万奇, 王兆锋, 郑度. 青藏高原高寒草地净初级生产力(NPP)时空分异. 地理学报, 2013, 68(9): 1197-1211. |
| [15] |
Sitch S, Smith B, Prentice I C, Arneth A, Bondeau A, Cramer W, Kaplan J O, Levis S, Lucht W, Sykes M T, Thonicke K, Venevsky S. Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Global Change Biology, 2003, 9(2): 161-185. DOI:10.1046/j.1365-2486.2003.00569.x |
| [16] |
Mack M C, Schuur E A G, Bret-Harte M S, Shaver G R, Chapin III F S. Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization. Nature, 2004, 431(7007): 440-443. DOI:10.1038/nature02887 |
| [17] |
Zhang L, Guo H D, Jia G S, Wylie B, Gilmanov T, Howard D, Ji L, Xiao J F, Li J, Yuan W P, Zhao T B, Chen S P, Zhou G S, Kato T. Net ecosystem productivity of temperate grasslands in northern China: an upscaling study. Agricultural and Forest Meteorology, 2014, 184: 71-81. DOI:10.1016/j.agrformet.2013.09.004 |
| [18] |
Terrer C, Vicca S, Hungate B A, Phillips R P, Prentice I C. Mycorrhizal association as a primary control of the CO2 fertilization effect. Science, 2016, 353(6294): 72-74. DOI:10.1126/science.aaf4610 |
| [19] |
Girardin M P, Bernier P Y, Raulier F, Tardif J C, Conciatori F, Guo X J. Testing for a CO2 fertilization effect on growth of Canadian boreal forests. Journal of Geophysical Research: Biogeosciences, 2011, 116(G1): G01012. |
| [20] |
张方敏. 美国森林碳源汇分布的模拟研究[D]. 南京: 南京信息工程大学, 2012.
|
| [21] |
张文海, 吕锡芝, 余新晓, 范敏锐. 气候和CO2变化对北京山区油松林NPP的影响. 广东农业科学, 2012, 39(6): 4-7. DOI:10.3969/j.issn.1004-874X.2012.06.002 |
| [22] |
Shao Q Q, Sun C Y, Liu J Y, He J F, Kuang W H, Tao F L. Impact of urban expansion on meteorological observation data and overestimation to regional air temperature in China. Journal of Geographical Sciences, 2011, 21(6): 994-1006. DOI:10.1007/s11442-011-0895-9 |
| [23] |
Veroustraete F, Sabbe H, Eerens H. Estimation of carbon mass fluxes over Europe using the C-Fix model and Euroflux data. Remote Sensing of Environment, 2002, 83(3): 376-399. DOI:10.1016/S0034-4257(02)00043-3 |
| [24] |
Luo X, Jia B H, Lai X. Contributions of climate change, land use change and CO2 to changes in the gross primary productivity of the Tibetan Plateau. Atmospheric and Oceanic Science Letters, 2020, 13(1): 8-15. DOI:10.1080/16742834.2020.1695515 |
| [25] |
孙静, 范文义, 于颖, 王斌, 陈晨. 基于InTEC模型的塔河森林净初级生产力影响因子定量分析. 应用生态学报, 2019, 30(3): 793-804. |
| [26] |
魏书精, 罗碧珍, 孙龙, 魏书威, 刘芳芳, 胡海清. 森林生态系统土壤呼吸时空异质性及影响因子研究进展. 生态环境学报, 2013, 22(4): 689-704. DOI:10.3969/j.issn.1674-5906.2013.04.024 |
| [27] |
辛丽花, 韩士杰, 郑俊强, 周玉梅, 李莉. CO2浓度升高对土壤微生物及土壤酶影响的研究进展. 土壤通报, 2006, 36(6): 1231-1235. DOI:10.3321/j.issn:0564-3945.2006.06.042 |
| [28] |
周玉梅, 韩士杰, 郑俊强, 辛丽花, 张海森. CO2浓度升高对森林土壤微生物呼吸与根(际)呼吸的影响. 植物生态学报, 2007, 31(3): 386-393. DOI:10.3321/j.issn:1005-264X.2007.03.007 |
| [29] |
Runion G B, Curl E A, Rogers H H, Backman P A, Rodríguez-Kábana R, Helms B E. Effects of free-air CO2 enrichment on microbial populations in the rhizosphere and phyllosphere of cotton. Agricultural and Forest Meteorology, 1994, 70(1/4): 117-130. |
| [30] |
宋清洁, 崔霞, 张瑶瑶, 孟宝平, 高金龙, 向宇轩. 基于小型无人机与MODIS数据的草地植被覆盖度研究——以甘南州为例. 草业科学, 2017, 34(1): 40-50. |
| [31] |
马丽, 徐满厚, 翟大彤, 贾燕燕. 高寒草甸植被-土壤系统对气候变暖响应的研究进展. 生态学杂志, 2017, 36(6): 1708-1717. |
| [32] |
马文瑛, 赵传燕, 彭守璋, 高云飞, 袁亚鹏, 李文娟. 黑河天涝池五种植被类型土壤呼吸速率动态特征及其影响因子. 生态学报, 2015, 35(17): 5654-5665. |
| [33] |
Haverd V, Smith B, Canadell J G, Cuntz M, Mikaloff-Fletcher S, Farquhar G, Woodgate W, Briggs P R, Trudinger C M. Higher than expected CO2 fertilization inferred from leaf to global observations. Global Change Biology, 2020, 26(4): 2390-2402. DOI:10.1111/gcb.14950 |
| [34] |
Ainsworth E A, Rogers A. The response of photosynthesis and stomatal conductance to rising[CO2]: mechanisms and environmental interactions. Plant, Cell & Environment, 2007, 30(3): 258-270. |
| [35] |
Smith K W, Reed S C, Cleveland C C, Ballantyne A P, Anderegg W R L, Wieder W R, Liu Y Y, Running S W. Large divergence of satellite and earth system model estimates of global terrestrial CO2 fertilization. Nature Climate Change, 2016, 6(3): 306-310. DOI:10.1038/nclimate2879 |
| [36] |
卢玲, 李新, Veroustraete F. 中国西部地区植被净初级生产力的时空格局. 生态学报, 2005, 25(5): 1026-1032. DOI:10.3321/j.issn:1000-0933.2005.05.013 |