 2021, Vol. 41
2021, Vol. 41文章信息
- 莫冉, 宋卫信, 李群, 张锋
- MO Ran, SONG Weixin, LI Qun, ZHANG Feng
- 代谢互养关系在维持微生物物种多样性中的作用
- The role of metabolic cross-feeding in maintaining microbial species diversity
- 生态学报. 2021, 41(16): 6506-6512
- Acta Ecologica Sinica. 2021, 41(16): 6506-6512
- http://dx.doi.org/10.5846/stxb202010222691
-
文章历史
- 收稿日期: 2020-10-22
- 网络出版日期: 2021-05-21

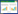




物种多样性的产生和维持机制一直是生态学领域的热点问题。传统的竞争排斥原理认为, 多物种竞争单一有限的资源时, 只有竞争力最强的可以存活, 其他参与者都会消亡[1-2]。后来, 现代竞争共存理论提出, 竞争物种也可以在同一环境中共存[3]。那么怎么从物种间相互作用的角度来解释竞争共存的机制?近年, 微生物生态学的发展揭示出, 微生物物种之间普遍存在的代谢互养关系(cross-feeding)可能是维持微生物多样性的一种机制[4-8]。实验室研究表明, 在连续培养中的长期进化研究表明, 受单一资源限制的微生物群体(如肠道微生物、土壤微生物)会进化出稳定的多态性, 对种群结构进行分析:降解优势生长的菌株排泄出的产物可以用作另一种菌株的底物资源促使其生长[9-13]。这种物种1产生的代谢资源可以被物种2利用, 促进物种2生长的关系称为单向互养关系。如果将多个竞争同一有限碳源的微生物物种混合在一起培养, 发现它们能够稳定共存, 此时外界碳源耗尽, 培养基中的碳源都来自这些微生物细胞的代谢副产物[14-16]。这种物种间利用彼此代谢资源的关系称为双向互养关系。
互养关系的广泛存在已得到大量实验验证, 但是相关的理论研究还比较少[7, 15, 17-19], 尤其实验中很难量化观察到代谢产物的产生率[14], 所以数学模型是解开互养关系复杂性的重要工具。本文考虑两物种间单向、双向及多物种间双向互养的三种关系结构, 整合资源及物种动态, 建立微生物互养关系的动力学模型, 系统地研究了互养关系中代谢资源的利用能否促进共存以及如何促进共存。
1 模型与方法考虑不同的互养关系结构:两物种间单向互养、双向互养以及多物种间的互养, 基于资源竞争模型构建了包含互养关系的物种及资源变化的动力学模型, 利用R*法则判断资源与种群生长之间的关系。
1.1 经典资源竞争模型当多物种竞争同一外界供应的有限的资源时, 经典资源竞争模型[20-22]表示如下:
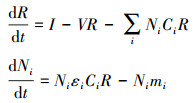
|
(1) |
其中, R表示外界供给的有限可利用资源, Ni表示物种i的种群密度, I表示外界资源R的供应率, V表示单位资源R的自然消耗率, Ci表示物种i对资源R的摄取率, εi表示物种i对资源R的利用率, mi表示物种i的死亡率。竞争排斥原理主张共存的物种必须占据独特的生态位[23-24], 即存活物种的数量不能超过资源的数量。由模型(1)中物种动态方程得, 平衡态时资源临界值Ri*=mi/εiCi, 在竞争中获胜的物种是临界资源需求R*最低的物种[20-21, 25-26], 其他竞争者都被淘汰, 此时物种量由资源量决定, 表达式为Ni=(I-VR)/CiR。当Ri≥Ri*时, dNi/dt≥0, 物种i的种群密度增大;当Ri < Ri*时, dNi/dt < 0, 物种i的种群密度开始减小直至该物种消亡。所以物种及资源数量达到平衡态时, 若在竞争中占据优势的为物种1, 则资源数量为R1*(R2*> R1*), 此时物种1生存, 物种2被排斥。
| 符号 Symbol |
表示的含义 Meaning of symbol |
符号 Symbol |
表示的含义 Meaning of symbol |
|
| I | 外界资源供应率 | C | 物种对资源的摄取率 | |
| V | 单位资源自然消耗率 | ε | 物种对资源的利用率 | |
| R | 资源数量 | μ | 物种对代谢资源的利用率 | |
| N | 物种的种群密度 | P | 代谢资源产生率 | |
| m | 物种的死亡率 |
允许竞争者共存的条件是打破竞争排除, 比如通过资源动态的改变影响临界资源R*对物种生长的限制[27]。当物种1产生的代谢资源被物种2消耗的这种互养关系可以抵消它相对于物种2的竞争优势时, 两个物种就可以共存(图 2A)。为了量化给定环境条件下物种间相互作用的强度和迹象, 给定参数P、μ, 当外界资源α持续供给时, 图 2A所示关系的动力系统模型可表示为:
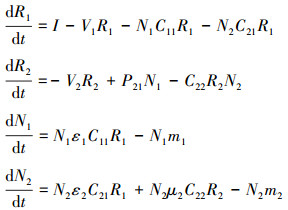
|
(2) |
其中, Vj表示单位资源j的自然消耗率, Cij表示物种i对资源j的摄取率, Pji表示物种i产生资源j的产生率, μi表示物种i对代谢资源的利用率。当R1≥R2*时, 两物种同时利用外界资源R1生长;当R1 < R2*时, 物种2不再利用资源R1, 开始衰落, 但是代谢资源R2的存在使得物种2的种群密度继续增加, 直至达到新的平衡。在竞争外界资源α时, 物种1相对于物种2有竞争优势, 但是物种2有代谢资源β的加持, 所以物种1、2很可能共存, 并且在资源β和α的共同支撑下, 物种2很可能“弱者逆袭”, 占据竞争优势。
在模型(2)中, 只存在一种代谢资源, 由物种1产生并供养物种2, 接下来考虑环境中存在两种代谢资源的情况。在营养资源α有限的环境中, 物种1产生的代谢资源可以作为物种2生长所需的营养物质, 同样, 物种2产生的代谢资源也可以作为物种1生长用的营养物质, 形成双向互养关系(图 2B), 具体表示为:
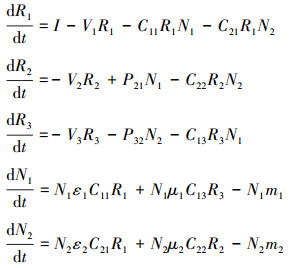
|
(3) |
该模型中符号含义同上。只有一种代谢资源R2时, 当R1=R1*, 物种1的种群密度不再变化, 但是加入代谢资源R3后, R1 < R1*时, 物种1的种群密度还可以继续增加, 所以该模型中资源R的利用率提高, 平衡态时两物种的种群密度也相应变大。且双向互养关系中优势物种是1或2, 互养关系都可以促进共存。
1.3 多物种互养关系的动力学建模前面的模型只考虑了两物种之间的互养关系, 可是自然界中物种之间的关系通常以复杂的生态网形式存在, 这里将互养关系推广至多物种之间(图 2C), 模型表示为:
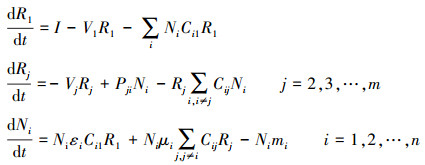
|
(4) |
该模型中我们假设每个物种都会产生一种代谢资源, 且该代谢资源可被竞争环境中除自身以外的其他所有参与者利用。类似地, 该模型中资源的利用率会进一步提高, 但是由于每种代谢资源的利用者较多, 所以各物种的种群密度可能较模型(3)中的种群密度有所减小。
2 结果与分析实际分析中模型的解析解难求, 所以通过数值模拟来验证上述的理论分析, 采用的软件是Matlab2017b, 算法是龙格-库塔法(ode45)。模拟中物种的种群密度的初值设定为10, 外界供给资源数量的初值设定为100, 代谢资源数量的初值为0。图 1给出了没有代谢资源时种群密度及资源数量的动态变化情况, A中R1>R2*时, 物种1、2种群密度增加;R1* < R1 < R2*时, 物种2的种群密度减小, 物种1的种群密度增加;R1=R1*时, 种群密度及资源量达到平衡态, 此时物种2的种群密度为0;同理, B中四物种竞争同一种有限资源时, 最终只有一个物种可以存活。

|
| 图 1 没有互养关系的资源竞争结果 Fig. 1 Results of competition for resources without cross-feeding A: 单向互养关系; B: 双向互养关系 |
图 2中A可以促进两物种共存;R1* < R1 < R2*时, 物种2利用代谢资源R2, 随着资源摄取总量的增加, 物种2的种群密度变大, 若代谢资源的产生率及利用率足够大, 物种2的种群密度超过物种1, “弱者逆袭”。B可以促进两物种共存;R1 < R2*时, 物种2可以利用代谢资源R2继续生长, 同样R1 < R1*时, 物种1可以利用代谢资源R3继续生长, 所以双向互养关系中物种的种群密度明显高于单向互养关系的。图 2C表示四物种竞争同一种有限资源时, 物种可以利用彼此的代谢资源, 产生共存。故不同的互养关系结构中竞争物种都可以共存, 且通过资源量与种群密度关系分析得劣势物种通过利用其他竞争成员的代谢产物, 打破外界资源量对其生长的限制, 改变原本消亡的命运, 优势物种通过利用其他竞争成员的代谢产物, 增大种群密度。

|
| 图 2 不同互养关系的资源竞争结果 Fig. 2 Results of resource competition in different cross-feeding relationships |
但是图 2C中有出现一个奇怪的现象, 在外界资源以及代谢资源的共同支撑下, 还是有物种灭绝。数值模拟中, 不同物种对外界资源的利用率取值是不同的, 有大有小, 而最后灭绝的确是其中处于竞争劣势的物种(物种2)。对比图 1A和图 2A, 在其他参数都不变的情况下物种1产生代谢资源给物种2利用, 这个过程中, 物种1的种群密度是有所减小的, 所以物种产生代谢资源对其本身是“不利”的。而且如果在模拟中物种利用代谢资源的能力相同, 那么物种竞争外界资源的劣势就很可能无法被抵消。所以在多物种竞争及互养关系中, 本身处于竞争劣势的物种当其产生代谢资源过多且各竞争者利用代谢资源能力相当时, 超出自身能力时, 它便会逐渐被淘汰, 从而灭绝[15]。图 3的A和B分别提高物种2对外界资源R1的利用率以及对代谢资源R2、R4、R5的利用率, 模拟发现物种2可以因此改变灭绝的命运, 与其它3个物种共存。类似地, 当物种数更多时, 只有互养关系中代谢资源的利用可以弥补劣势种在竞争外界资源时的不足, 多物种才可以全部共存。

|
| 图 3 多物种互养关系中改变参数对资源竞争结果的影响 Fig. 3 Effects of changing parameters on resource competition outcomes in cross-feeding of multiple species |
图 4分别给出了单向互养关系中物种1、2在平衡态时的种群密度, 代谢资源产生率与利用率均大于0是两竞争物种共存的必要条件, 参数值的变化不影响共存的结果。其他互养关系结果类似, 表明互养关系促进的共存对代谢资源相关参数不敏感, 参数的改变只影响平衡态时的物种的种群密度。

|
| 图 4 代谢资源参数对平衡态物种的种群密度的影响 Fig. 4 Effects of metabolic resource parameters on population density of equilibrium species |
包含互养关系的竞争系统中具有丰富的动力学行为, 标准的Lotka-Volterra方程只解释了一种相互作用(互惠或竞争), 而广义Lotka-Volterra方程能够准确地模拟竞争和互惠共存的同时发生, 但却忽视了相互作用间的机制[28]。Goldford等人[15]通过经典的MacArthur消费者-资源模型模拟验证了包括非特异性互养关系的群落构建过程, 所有共存的物种都能够在单一供应资源和其他物种的分泌物中生长, 但这些模型的细节缺乏普遍性, 无法解释其他实验结果;本文的不同之处则在于分析建立不同结构互养关系的动力学模型, 使得互养关系的概念及结构更加清晰。Sun等人[18]建立两物种间互养关系的模型, 二者产生的代谢资源可以被对方利用, 并对模型进行定性分析, 当营养资源的外部供应有限且代谢资源的合成率较高时, 共存是有可能的。本文进一步考虑多物种间的互养关系, 通过数值模拟结果对模型进行验证分析。
代谢资源的存在是自然生态系统的重要现象。资源竞争理论说明生态系统中随着外界供给资源的消耗只保留一种优势物种的现象。本文研究表明物种通过利用其他竞争成员的代谢产物, 打破外界资源量对其生长的限制, 使得原本被排斥的物种得以继续生长, 之后资源的竞争使得各物种及资源达到动态平衡, 从而实现竞争物种稳定共存。另外, 通过对模型中代谢资源相关参数变化的分析, 证明互养关系促进的共存对这些参数变化不敏感, 参数的改变只影响物种的种群密度的大小。所以得出结论是微生物之间的互养关系可能是维持生物多样性的一种机制。
将本文的模型扩展到空间生态模型可以更好地讨论物种间的交流, 进一步研究物种间互养关系产生的进化是非常有趣的[29]。另外, 本文是通过建立确定性模型展开讨论, 但是由于自然环境的不确定性与随机性, 微生物物种的种群密度随时间变化是一个随机过程, 那么考虑种群统计随机性[30-31], 在模型中加入随机干扰共存结果是否会受影响, 这将是下一步要做的工作。
| [1] |
Tilman D. Resource Competition and Community Structure. Princeton: Princeton University Press, 1982.
|
| [2] |
Gause G F. Experimental analysis of Vito Volterra's mathematical theory of the struggle for existence. Science, 1934, 79(2036): 16-17. |
| [3] |
梁仁君, 林振山, 韩洪凌, 陈成忠. 物种对资源竞争的动力机制及数值模拟. 生态学报, 2007, 27(12): 5390-5397. DOI:10.3321/j.issn:1000-0933.2007.12.052 |
| [4] |
Morris B E L, Henneberger R, Huber H, Moissl-Eichinger C. Microbial syntrophy: interaction for the common good. FEMS Microbiology Reviews, 2013, 37(3): 384-406. DOI:10.1111/1574-6976.12019 |
| [5] |
Bronstein J L. The exploitation of mutualisms. Ecology Letters, 2001, 4(3): 277-287. DOI:10.1046/j.1461-0248.2001.00218.x |
| [6] |
Holert J, Yücel O, Suvekbala V, Kulić Ž, Möller H, Philipp B. Evidence of distinct pathways for bacterial degradation of the steroid compound cholate suggests the potential for metabolic interactions by interspecies cross-feeding. Environmental Microbiology, 2014, 16(5): 1424-1440. DOI:10.1111/1462-2920.12407 |
| [7] |
Henson M A, Phalak P. Byproduct cross feeding and community stability in an in silico biofilm model of the gut microbiome. Processes, 2017, 5(1): 13. |
| [8] |
Van Hoek M J A, Merks R M H. Emergence of microbial diversity due to cross-feeding interactions in a spatial model of gut microbial metabolism. BMC Systems Biology, 2017, 11(1): 56. DOI:10.1186/s12918-017-0430-4 |
| [9] |
Pfeiffer T, Bonhoeffer S. Evolution of cross-feeding in microbial populations. The American Naturalist, 2004, 163(6): E126-E135. DOI:10.1086/383593 |
| [10] |
Hoek T A, Axelrod K, Biancalani T, Yurtsev E A, Liu J H, Gore J. Resource availability modulates the cooperative and competitive nature of a microbial cross-feeding mutualism. PLoS Biology, 2016, 14(8): e1002540. DOI:10.1371/journal.pbio.1002540 |
| [11] |
Turroni F, Milani C, Duranti S, Mahony J, van Sinderen D, Ventura M. Glycan utilization and cross-feeding activities by bifidobacteria. Trends in Microbiology, 2018, 26(4): 339-350. DOI:10.1016/j.tim.2017.10.001 |
| [12] |
Krause S M B, Johnson T, Karunaratne Y S, Fu Y F, Beck D A C, Chistoserdova L, Lidstrom M E. Lanthanide-dependent cross-feeding of methane-derived carbon is linked by microbial community interactions. Proceedings of the National Academy of Sciences of the United States of America, 2017, 114(2): 358-363. DOI:10.1073/pnas.1619871114 |
| [13] |
Seth E C, Taga M E. Nutrient cross-feeding in the microbial world. Frontiers in Microbiology, 2014, 5: 350. |
| [14] |
Pacheco A R, Moel M, Segrè D. Costless metabolic secretions as drivers of interspecies interactions in microbial ecosystems. Nature Communications, 2019, 10(1): 103. DOI:10.1038/s41467-018-07946-9 |
| [15] |
Goldford J E, Lu N X, Bajić D, Estrela S, Tikhonov M, Sanchez-Gorostiaga A, Segrè D, Mehta P, Sanchez A. Emergent simplicity in microbial community assembly. Science, 2018, 361(6401): 469-474. DOI:10.1126/science.aat1168 |
| [16] |
Smith N W, Shorten P R, Altermann E, Roy N C, McNabb W C. The classification and evolution of bacterial cross-feeding. Frontiers in Ecology and Evolution, 2019, 7: 153. DOI:10.3389/fevo.2019.00153 |
| [17] |
Muscarella M E, O'Dwyer J P. Species dynamics and interactions via metabolically informed consumer-resource models. Theoretical Ecology, 2020, 13(4): 503-518. DOI:10.1007/s12080-020-00466-7 |
| [18] |
Sun Z P, Koffel T, Stump S M, Grimaud G M, Klausmeier C A. Microbial cross-feeding promotes multiple stable states and species coexistence, but also susceptibility to cheaters. Journal of Theoretical Biology, 2019, 465: 63-77. DOI:10.1016/j.jtbi.2019.01.009 |
| [19] |
Niehaus L, Boland I, Liu M B, Chen K, Fu D, Henckel C, Chaung K, Miranda S E, Dyckman S, Crum M, Dedrick S, Shou W Y, Momeni B. Microbial coexistence through chemical-mediated interactions. Nature Communications, 2019, 10(1): 2052. DOI:10.1038/s41467-019-10062-x |
| [20] |
MacArthur R. Species packing and competitive equilibrium for many species. Theoretical Population Biology, 1970, 1(1): 1-11. DOI:10.1016/0040-5809(70)90039-0 |
| [21] |
Tilman D. Resource competition between plankton algae: an experimental and theoretical approach. Ecology, 1977, 58(2): 338-348. DOI:10.2307/1935608 |
| [22] |
Tilman D. Plant Strategies and the Dynamics and Structure of Plant Communities. Princeton: Princeton University Press, 1988.
|
| [23] |
储诚进, 王酉石, 刘宇, 蒋林, 何芳良. 物种共存理论研究进展. 生物多样性, 2017, 25(4): 345-354. |
| [24] |
Loreau M, Hector A. Partitioning selection and complementarity in biodiversity experiments. Nature, 2001, 412(6842): 72-76. DOI:10.1038/35083573 |
| [25] |
南春容, 董双林. 资源竞争理论及其研究进展. 生态学杂志, 2003, 22(2): 36-42. DOI:10.3321/j.issn:1000-4890.2003.02.010 |
| [26] |
王锋刚, 曾晓东. 植物种群资源竞争与共存的理论模型研究. 气候与环境研究, 2015, 20(2): 229-234. |
| [27] |
Cui W P, Marsland Ⅲ R, Mehta P. Effect of resource dynamics on species packing in diverse ecosystems. Physical Review Letters, 2020, 125(4): 048101. DOI:10.1103/PhysRevLett.125.048101 |
| [28] |
Vet S, de Buyl S, Faust K, Danckaert J, Gonze D, Gelens L. Bistability in a system of two species interacting through mutualism as well as competition: Chemostat vs. Lotka-Volterra equations. PLoS One, 2018, 13(6): e0197462. DOI:10.1371/journal.pone.0197462 |
| [29] |
Stump S M, Johnson E C, Sun Z P, Klausmeier C A. How spatial structure and neighbor uncertainty promote mutualists and weaken black queen effects. Journal of Theoretical Biology, 2018, 446: 33-60. DOI:10.1016/j.jtbi.2018.02.031 |
| [30] |
马祖飞, 李典谟. 种群统计随机性和环境随机性对种群绝灭的影响. 生态学报, 2003, 23(12): 2702-2710. DOI:10.3321/j.issn:1000-0933.2003.12.025 |
| [31] |
袁怀民, 张启敏. 一类受分数Brown运动影响的资源竞争种群系统的渐近行为. 河南师范大学学报: 自然科学版, 2018, 46(4): 8-13. |