 2021, Vol. 41
2021, Vol. 41文章信息
- 左雪燕, 崔丽娟, 李伟, 窦志国, 蔡杨, 刘志君, 雷茵茹
- ZUO Xueyan, CUI Lijuan, LI Wei, DOU Zhiguo, CAI Yang, LIU Zhijun, LEI Yinru
- 基于高光谱数据的互花米草叶片功能性状反演
- Inversion of functional traits of Spartina alterniflora leaves based on hyperspectral data
- 生态学报. 2021, 41(15): 6159-6169
- Acta Ecologica Sinica. 2021, 41(15): 6159-6169
- http://dx.doi.org/10.5846/stxb202004020781
-
文章历史
- 收稿日期: 2020-04-02
- 网络出版日期: 2021-05-20





2. 北京汉石桥湿地生态系统国家定位观测研究站, 北京 101399
2. Beijing Hanshiqiao National Wetland Ecosystem Research Station, Beijing 101399, China
滨海湿地生态系统是介于陆地和海洋之间的过渡带, 具有独特的生态过程, 具备保护海岸线、防止海岸侵蚀、调节气候、净化水质、为生物提供栖息地等功能, 是全球生物多样性最丰富、最具价值的生态系统之一[1-6]。但是随着人类活动和气候变化的加剧, 滨海湿地面临严重威胁, 生态系统功能遭到严重破坏[7-11]。互花米草的入侵是我国滨海湿地生态系统面临的严重问题之一, 因为缺少天敌, 且对环境的适应性较强而具有超强的竞争力, 在滨海滩涂湿地上逐渐蔓延, 造成本土植物多样性减少, 对经济、社会、生态均产生严重危害[12-14]。
植物功能性状是指与植物获得、利用和保存资源的能力有关的一系列植物形态生理学性状[15-17]。功能性状在器官水平上可以分为:全株性状、叶性状、根性状、茎性状等;还可以分为形态性状、生理性状等[16, 18-19]。叶片功能性状是植物功能性状的重要组成部分, 能够反映植物的生长策略和适应机制。其中, 比叶面积、叶片养分含量等与植物的相对生长速率存在显著相关关系[17];叶绿素浓度、叶氮、比叶面积、氮磷比等反映的是植物对资源的最大化获取的生存策略[20-21]。传统上的实验室内进行功能性状的测定方法费时费力, 而高光谱数据以其快速、高效且提供的信息量大为解决这些问题提供了有效手段[22-24]。
目前, 对于高光谱数据反演植被功能性状已经取得了很大的进展, 但是对于湿地的研究主要集中某一指标(如氮、磷、生物量)的研究[1, 25-34], 对互花米草的反演研究则主要集中于叶绿素含量、生物量等方面, 对于叶片功能性状的系统研究比较缺乏。因此, 本研究利用ASD FS4便携式地物光谱仪来获取互花米草的光谱数据, 基于高光谱数据建立与叶片功能性状之间的定量模型, 最后通过决定系数(R2)以及均方根误差(RMSE)比较选出最佳反演模型, 为实现无损监测互花米草的生长状况提供理论和技术基础。
1 材料与方法 1.1 研究区概况江苏盐城滨海湿地, 地处32°56′-33°36′N, 120°13′-120°56′E(图 1, 分类图来源于MODIS Land Cover Type Product, MCD12Q1)。位于淮河下游, 东临黄海, 有582 km海岸线。季风盛行, 雨水充沛, 冬季盛行来自高纬度陆地的偏北风, 夏季盛行来自太平洋的偏南风, 全年平均气温受到海洋的调节, 年、日变化小, 该地区属于弱潮区, 潮汐类型为不规则半日潮[35]。盐城滨海湿地是我国滨海湿地生态系统中类型最为齐全的区域, 也是中国最大的动态潮间系统。该研究区为淤长型盐沼湿地, 样地位于盐城大丰麋鹿自然保护区。植被以盐生植被为主, 有互花米草(Spartina alterniflora)、芦苇(Phragmites communis)、白茅(Imperata cylindrica)等[36], 其中互花米草自入侵以来, 在该保护区不断扩张, 对当地的生态和经济造成了严重威胁。

|
| 图 1 研究区位置以及样点分布 Fig. 1 Location of study area and distribution of sample points |
本研究于2019年8月在江苏盐城进行试验数据的采集工作, 使用的光谱仪是美国Analytical Spectral Device(ASD)公司生产的便携式地物波谱仪FS4, 光谱范围为350-2500 nm, 采样间隔为1 nm。平行于海岸线设置3个条带, 每个条带上采集20个采样点, 采样点间隔10 m。采样点植物群落单一, 均为互花米草群落, 高度为100-175 cm, 株数为80-160株/m2, 植被盖度为60%-80%, 土壤质地为砂性土和壤性土。在每个采样点采集上、中、下部三片生长健康、无病害的叶片, 用ASD FS4搭配的叶片夹测量互花米草叶片的中部, 每个叶片采集10条光谱数据, 测定时避开叶片的主叶脉, 将构成样本的三个叶片光谱反射率值进行平均作为该样点的叶片光谱反射率。
1.2.2 叶片功能性状测定在每个采样选择3株生长状况良好的互花米草植株, 采集地上植株上中下3个部位的互花米草叶片, 4小时内测定叶片的鲜重、叶厚、SPAD以及叶面积后, 在105℃下, 杀青15 min, 之后85℃恒温, 烘干至恒重, 得到干重;之后经过粉碎过筛, 测定其他指标。总氮和总碳可由元素分析仪同时获得, 通过高温燃烧使得植物样品分解, 生成混合气体之后由热导检测系统自动测定[37];植物体中的磷以有机态为主, 经过H2SO4-H2O2消煮使得有机磷转化为磷酸盐, 之后利用分光光度计测定, 绘制标准曲线, 测定总磷含量[38](表 1)。
| 叶片功能性状 Leaf founctional traits | 测定方法 Measurement methods |
| 叶片含水率 Moisture content(MC) | (鲜重-干重)/鲜重 |
| 相对叶绿素含量 Soil and plant analyzer develotrnent (SPAD) | 叶绿素计(SPAD-502Plus; Konica Minolta, Japan) |
| 比叶面积 Specific leaf area (SLA) | 扫描仪扫描叶片, 通过ImageJ软件提取叶面积, 比叶面积=单叶面积/单叶干重 |
| 总氮 Total nitrogen(TN) | 元素分析仪(vario PYRO cube; Elemental, Germany) |
| 总磷 Total phosphorus(TP) | 钼锑抗比色法(NY/T 2421-2013) |
| 总碳 Total carbon(TC) | 元素分析仪(vario PYRO cube; Elemental, Germany) |
(1) 光谱数据转换
通过ViewSpecPro软件对互花米草叶片高光谱数据进行处理, 导出互花米草叶片反射率(仪器自动重采样为1 nm分辨率), 350-2500 nm共2151条波段, 每30条波段取平均作为一个采样点的光谱反射率值, 同时对叶片原始光谱反射率进行一阶微分(First derivative, FD)转换, 公式为:
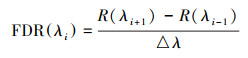
|
式中, λi为每个波段的波长, FDR(λi)为波长λi处的一阶微分光谱值, △λ为波段i到波段i+1的波长值。
(2) 主成分分析
高光谱数据的波段数量多、相关性强, 存在数据冗余, 造成计算复杂。主成分分析是常用的高光谱降维手段, 通过主成分分析, 将互相关联的多个变量压缩为少数不相关的新的成分变量, 少数的几个成分变量包含原始高光谱数据的大量信息。本研究中的主成分分析通过R语言软件中的psych函数来实现。
1.2.4 数据分析处理及模型建立与评价将通过主成分分析获得新的主成分作为自变量, 构建互花米草叶片功能性状的预测模型, 模型选择逐步回归模型(Stepwise regression, SR)、BP神经网络(BP artificial neural network, BPNN)、支持向量机(Support vector machines, SVM)、随机森林(Random forest, RF)模型, 模型的实现依托于软件Weka 3.8。基于R语言实现原始高光谱反射率以及一阶微分转换后的光谱数据与互花米草叶片功能性状的相关性分析, 选择最佳波段, 构建预测模型。模型精度由决定系数(R2)、均方根误差(RMSE)进行评价。最后, 通过Origin 2020软件出图。
2 结果与分析 2.1 互花米草叶片功能性状分析首先对互花米草的功能性状进行分析, 得到表 2的结果。从表中可以看出, 互花米草的叶片含水率均值为0.66, 范围为0.56-0.74, 含水率相对较高;SPAD为41.41, 标准偏差为5.17;TC/TN、TN、TP、SLA的均值分别为27.12、1.32 g/kg、0.18 g/kg、41.68, 其中SLA的标准偏差较大, 达到20.26。功能性状的取值范围以及标准偏差的存在说明了互花米草叶片之间的差异, 为基于高光谱数据进行功能性状的反演提供了条件。
| 叶片功能性状 Leaf founctional traits |
样本量 No. of samples |
最小值 Minimum |
最大值 Maximum |
均值 Mean |
标准偏差 Standard Deviation |
| 叶片含水率 Moisture content(MC) | 60 | 0.56 | 0.74 | 0.66 | 0.03 |
| 相对叶绿素含量 Soil and plant analyzer develotrnent (SPAD) |
60 | 30.67 | 53.23 | 41.41 | 5.17 |
| 碳氮比 TC/TN | 60 | 13.23 | 35.77 | 27.12 | 4.62 |
| 总氮 Total nitrogen(TN) | 60 | 0.92 | 2.37 | 1.32 | 0.28 |
| 总磷 Total phosphorus(TP) | 60 | 0.11 | 0.31 | 0.18 | 0.04 |
| 比叶面积 Specific leaf area(SLA) | 60 | 4.99 | 115.51 | 41.68 | 20.26 |
本文通过主成分分析将原始光谱反射率数据以及一阶微分数据进行降维, 减少自变量个数, 提高运算效率。从表 3可以看出原始光谱反射率的PC1的贡献率为0.53, PC2的贡献率为0.31, 前两个主成分累计贡献率达到0.84, 考虑到PC3的贡献率达到0.1, 因此保留前3个主成分, 累计贡献率达到0.94。从表 4可以看出, 一阶微分光谱主成分分析的第一主成分的贡献率为0.49, 第二主成分的贡献率为0.18, 其余主成分贡献率均低于0.1, 因此选择了累计贡献率达到0.8的前4个主成分作为新的变量。
| PC | 特征根 Eigenvalue |
贡献率 Proportion explained |
累计贡献率 Cumulative proportion |
| PC1 | 1132.48 | 0.53 | 0.53 |
| PC2 | 668.34 | 0.31 | 0.84 |
| PC3 | 216.48 | 0.10 | 0.94 |
| FDPC | 特征根 Eigenvalue |
贡献率 Proportion explained |
累计贡献率 Cumulative proportion |
| FDPC1 | 1053.20 | 0.49 | 0.49 |
| FDPC2 | 386.42 | 0.18 | 0.67 |
| FDPC3 | 176.36 | 0.08 | 0.75 |
| FDPC4 | 115.68 | 0.05 | 0.81 |
原始光谱反射率与叶片功能性状之间的相关性如图 2所示。分析结果表明, 叶片含水率在731-828、1365-1608、1742-1901、1977-2426 nm波段内与原始反射率之间显著相关, 与绿色植物的光谱反射率的水分敏感波段相似;SPAD在601-720 nm波段内极显著负相关(P < 0.01), 符合植物吸收绿光进行光合作用的原理;TC/TN与原始光谱数据的相关性较弱, 在426-448 nm,654-687 nm波段内呈显著正相关(P < 0.05), TN在1378-1572 nm, 1762-1891 nm波段内呈显著正相关, TC/TN与TN与原始光谱数据总体相关性较弱;总磷在1305-1390 nm1509-1836 nm范围内呈极显著正相关;SLA与原始光谱数据在720-1142 nm波段内呈极显著正相关(P < 0.01)。根据相关性分析的结果, 选择与互花米草功能性状显著相关的敏感波段作为自变量, 建立基于原始反射率敏感互花米草功能性状预测模型。

|
| 图 2 原始光谱反射率与叶片功能性状相关性 Fig. 2 Correlation between original spectral reflectance and leaf functional traits MC:含水率Moisture content;SPAD:相对叶绿素含量Soil and plant analyzer develotrnent;TC:总碳Total carbon;TN:总氮Total nitrogen;TP:总磷Total phosphorus;SLA:比叶面积Specific leaf area |
一阶微分转换后的高光谱数据与互花米草叶片功能性状的相关性分析结果如图 3所示。从结果中可以看出, 总体上一阶微分后的高光谱数据与功能性状的相关性明显高于原始光谱反射率, 并且相关性更为复杂。首先对于叶片含水率, 在541-560、804-967、977-998、1264-1387、1547-1593、1628-1687 nm等波段叶片与一阶微分高光谱数据显著相关(P < 0.01);SPAD与一阶微分数据在641-675、679-702、711-760 nm波段内极显著相关(P < 0.01);TC/TN在765-775、917-961、1004-1049、1722-1743、2213-2244 nm等波段内显著相关(P < 0.05);TN在1076-1087、1275-1362、1725-1743 nm等波段内与一阶微分数据显著相关;TP与一阶微分数据在1461-1542、1678-1712、2055-2127 nm等波段内极显著相关(P < 0.01);SLA与一阶微分数据在900-971、1084-1166、1266-1387、1722-1771 nm等波段内呈极显著相关。根据相关性分析的结果, 选择与互花米草功能性状显著相关的敏感波段作为自变量, 建立基于一阶微分敏感波段的互花米草功能性状预测模型。

|
| 图 3 一阶微分数据与叶片功能性状相关性分析 Fig. 3 Correlation analysis between first derivative spectral data and leaf functional traits |
根据上述表 3、表 4的结果, 分别基于原始光谱、一阶微分提取的主成分构建不同功能性状的SR、BPNN、SVM、RF预测模型。随机选择40个样本用于建模, 其余用于验证, 最优模型的确定由决定系数(R2)和均方根误差(RMSE)来确定。
2.5.1 基于主成分分析的叶片功能性状建模与评价分别基于原始和一阶微分变换的高光谱数据的主成分变量构建SR、BPNN、SVM、RF4种预测模型, 建模结果如表 5所示。对于两种数据形式的建模结果, 除了TC/TN、TN、SLA三个性状的极个别模型一阶微分的建模效果略低于原始高光谱数据外, 其余均明显优于原始高光谱数据建模, 尤其从SR来看, 原始光谱数据对TC/TN以及TN的建模精度极低, 而一阶微分则得到了显著性的关系结果, 虽然精度较低, 但也说明一阶微分有助于减少建模过程中的不确定性, 从而提高建模精度。比较4种模型的建模结果, SR、BPNN、SVM、RF的建模R2依次为0-0.75、0.34-0.89、0.58-1.00、0.98-0.99, 虽然SVM的部分建模结果优于RF模型, 但是较之于SVM模型, RF模型的稳定性更高, 综合比较可以发现, 4种模型的排序为RF>SVM>BPNN>SR, SR模型整体上较差, 可能是由于高光谱数据与叶片功能性状之间的关系是非线性的, 因此其建模效果较差。
| 叶片功能性状 Leaf founctional traits |
模型 Model |
原始 Original | 一阶微分 First derivative | |||
| R2 | RMSE | R2 | RMSE | |||
| 叶片含水率 Moisture content(MC) |
SR | 0.37 | 0.03 | 0.75 | 0.02 | |
| BPNN | 0.70 | 0.03 | 0.89 | 0.02 | ||
| SVM | 0.99 | 0.00 | 1.00 | 0.00 | ||
| RF | 0.98 | 0.01 | 0.99 | 0.01 | ||
| 相对叶绿素含量 Soil and plant analyzer develotrnent(SPAD) |
SR | 0.36 | 4.38 | 0.42 | 4.18 | |
| BPNN | 0.56 | 4.48 | 0.67 | 3.98 | ||
| SVM | 0.78 | 4.55 | 0.84 | 4.56 | ||
| RF | 0.99 | 1.73 | 0.99 | 1.70 | ||
| TC/TN | SR | 0.08 | 4.47 | 0.11 | 4.66 | |
| BPNN | 0.34 | 4.60 | 0.39 | 4.45 | ||
| SVM | 0.58 | 4.46 | 0.86 | 4.24 | ||
| RF | 0.99 | 1.74 | 0.98 | 1.98 | ||
| 总氮 Total nitrogen(TN) |
SR | - | - | 0.10 | 0.28 | |
| BPNN | 0.41 | 0.27 | 0.32 | 0.27 | ||
| SVM | 0.72 | 0.20 | 0.99 | 0.04 | ||
| RF | 0.98 | 0.11 | 0.98 | 0.12 | ||
| 总磷 Total phosphorus(TP) |
SR | 0.22 | 0.04 | 0.34 | 0.04 | |
| BPNN | 0.63 | 0.04 | 0.70 | 0.03 | ||
| SVM | 0.99 | 0.00 | 1.00 | 0.00 | ||
| RF | 0.98 | 0.02 | 0.98 | 0.02 | ||
| 比叶面积 Specific leaf area(SLA) |
SR | 0.42 | 17.04 | 0.32 | 16.98 | |
| BPNN | 0.66 | 16.67 | 0.78 | 16.66 | ||
| SVM | 0.78 | 20.85 | 0.77 | 20.95 | ||
| RF | 0.99 | 7.34 | 0.98 | 7.11 | ||
| SR:逐步回归模型Stepwise regression;BPNN:BP神经网络BP artificial neural network;SVM:支持向量机Support vector machines;RF:随机森林Random forest;RMSE:Root mean square error | ||||||
综合上述基于主成分建模的结果发现, 虽然SVM构建的模型效果有两次优于RF模型, 但是其建模效果不稳定, 建模R2介于0.58-1.00, 而RF模型的建模R2均大于0.90。综合来看, RF模型更加准确且稳定。鉴于以上, 基于相关性分析得到的敏感波段建立RF回归模型, 验证RF模型对互花米草叶片功能性状的预测精度以及稳定性。从互花米草叶片功能性状的RF模型(表 6、图 4)的结果来看, 建模R2均大于0.90, 验证R2介于0.73-0.95之间, 所有模型均得到了较好的结果。比较原始光谱敏感波段和一阶微分光谱敏感波段建模的结果发现, 一阶微分数据下的TN含量以及SLA的建模精度(RTN2=0.97, RMSETN=0.11;RSLA2=0.94, RMSESLA=8.10)低于原始光谱建模(RTN2=0.97, RMSETN=0.10;RSLA2=0.94, RMSESLA=8.15), 其余均优于原始光谱;对于验证模型的验证, 除TP的一阶微分验证R2低于原始外, 其他均优于原始。总体上说明了对原始光谱数据进行一阶微分转换有助于提高预测的准确性, 也表明随机森林模型对叶片功能性状预测的可行性。
| 叶片功能性状 Leaf founctional traits |
原始光谱 Original | 一阶微分 First derivative | |||
| R2 | RMSE | R2 | RMSE | ||
| 叶片含水率 Moisture content(MC) | 0.94 | 0.01 | 0.96 | 0.01 | |
| 相对叶绿素含量 Soil and plant analyzer develotrnent(SPAD) | 0.96 | 1.69 | 0.97 | 1.48 | |
| TC/TN | 0.96 | 1.97 | 0.97 | 0.01 | |
| 总氮 Total nitrogen(TN) | 0.97 | 0.11 | 0.97 | 0.10 | |
| 总磷 Total phosphorus(TP) | 0.96 | 0.02 | 0.96 | 0.02 | |
| 比叶面积 Specific leaf area(SLA) | 0.94 | 8.10 | 0.94 | 8.15 | |

|
| 图 4 互花米草叶片功能性状随机森林验证集预测值和真实值 Fig. 4 Predicted and true values of S. alterniflora leaf functional traits in a random forest validation set |
通过对互花米草不同叶片功能性状与原始高光谱以及一阶变换后的高光谱数据的相关性分析, 针对不同指标得到了不同的相关性分析结果, 其中MC主要在红外区域与高光谱数据存在显著相关关系, 主要是由于在红外光范围内, 植物的光谱反射率会受到水分的影响, 并且该结果也得到了浦瑞良等[22]和哈布热等[39]研究的证实;与SPAD显著相关的波段主要集中于可见光的绿光波段, 这主要是因为植物主要通过吸收绿光来进行光合作用, 该结果在前人的研究中得到了肯定[40-42];TC/TN、TN、TP、SLA与高光谱数据显著相关的波段主要集中于近红外和红外波段, 其中TC/TN、TN与原始光谱反射率的相关性较低, 与易秋香[43]等以及施润和等[44]的相关性绝对值达到0.85和0.81相差较大, 但是TN与一阶微分光谱在675、770、850、1076-1850 nm范围内与易秋香等[43]的研究结果存在交叉;TP与原始光谱的结果与郭澎涛等[45]的TP在529-578 nm以及699-746 nm与原始反射率显著相关的结果不同, 而TP与一阶微分光谱的相关性与其研究结果存在交叉波段;对于以上结果, 推断原因有二, 一是由于植物种类的差异, 使得光谱以及指标之间都存在差异;二是由于样本数量的不足, 导致相关性水平较低[43]。因此, 本研究得到的结果既与前人的研究存在相似之处, 又存在一定独特性, 说明不同的植物类型、样本量以及不同的数据处理方法, 会使得功能性状与高光谱数据之间的相关性分析存在差异。
3.2 基于不同数据类型的不同模型估算结果比较分析两种数据形式与功能性状的相关性分析结果表明, 在相关性水平上以及敏感波段数目上均证明了一阶微分光谱数据与叶片功能性状的相关性水平更高。根据表 5、6及图 4中不同模型建模以及验证精度的结果进一步验证了一阶微分变换的高光谱数据的预测效果优于原始数据。以上结果, 与郭超凡等[40]、艾金泉等[42]、余哲修等[46]的结果一致, 说明一阶微分光谱能够提高高光谱与功能性状的相关性, 提高功能性状的反演精度。在本研究中, 选择SR、BPNN、SVM、RF4种回归模型进行功能性状的预测, SR模型的建模R2介于0.10-0.75, BPNN建模R2介于0.32-0.89, SVM建模R2介于0.58-1, RF模型的建模R2介于0.98-0.99, 与前人的结果相比较, 本研究取得的结果是相对准确的(表 7)。
以主成分分析结果为变量, 比较4种模型在不同指标上的估算结果发现, 逐步回归模型在MC、SPAD、TP以及SLA的估算精度明显优于TC/TN和TN(表 5);而其他3种模型在不同指标上的结果无较大变化, 因此, 其他3种模型较逐步回归模型更加稳定, 其中RF模型的结果更加稳定。综上所述, RF优于其他3种模型, 可能是因为RF的建模过程随机性强, 决策树的节点分裂以及每个节点分割变量的产生都是随机的, 且不需要进行参数调整, 也不易出现过拟合现象, 因而更加稳定和准确[50, 51];SVM和BPNN预测效果不稳定的原因可能是其模型的构建需要较大的样本量来支撑, 而本研究中的样本量不足以使其稳定, 并且在Cui等[52]的研究中发现, RF、SVM、BPNN在不同季节不同指标的反演中均达到了很好的效果, 因此排除了采样时间这一影响因素;而SR模型效果较差的原因可能是由于功能性状与高光谱之间的关系是非线性的, 因而得到的模型准确性差。由于植物种类和性状指标的差异, 模型的适用性会存在差异, 本研究中RF模型在现有指标以及现有样本量情况下都取得了很好结果, 而其他模型的效果还有待进一步的验证。
4 结论本研究以江苏盐城滨海湿地的互花米草为研究对象, 采用室内光谱仪获得互花米草的高光谱反射率, 通过实验室测定获得互花米草叶片的相对叶绿素含量、叶片含水率、比叶面积、叶氮、叶磷、碳氮比指标。阐明了不同叶片功能性状与高光谱数据的相关性, 选出了敏感波段;比较了逐步回归、BP神经网络、支持向量机、随机森林4种模型的预测效果, 并进一步验证了随机森林模型的适用性和准确性, 主要结论如下:
(1) 不同性状与高光谱数据对应的敏感波段存在差异, 其中, 与叶片含水率显著相关的波段主要位于近红外和红外光波段;相对叶绿素含量的敏感波段则位于可见光波段;碳氮比和比叶面积两个性状与原始光谱数据的可见光波段高度相关, 总氮、总磷则与原始光谱数据的红外波段高度相关, 而对于一阶微分光谱数据, 相关性比较复杂, 敏感波段既存在于可见光波段, 又存在于红外波段, 显著相关的波段远多于原始高光谱反射率。总体上来说, 一阶微分变换的光谱数据与功能性状之间的相关性更强, 有助于提高建模精度。
(2) 通过对原始光谱数据以及一阶微分变换后的高光谱数据进行主成分分析提取新的主成分参数, 建立不同功能性状的预测模型, 综合比较不同性状不同模型的结果, 发现随机森林模型>支持向量机>BP神经网络>逐步回归, 其中随机森林的建模以及验证精度均大于0.80, 逐步回归模型的建模精度明显低于其他模型, 原因可能是功能性状与高光谱数据之间的关系是非线性的, 因而导致模型结果存在较大差异。
本研究进一步证实了原始光谱数据以及一阶微分数据对互花米草不同功能性状的预测能力, 可为大范围监测互花米草的入侵提供先验知识以及基础数据。同时本研究还进一步证实了随机森林模型的适应性和稳定性, 但该结果还需要大量的实验来进行检验和修正。
| [1] |
Wang L W, Wei Y X. Revised normalized difference nitrogen index (NDNI) for estimating canopy nitrogen concentration in wetlands. Optik, 2016, 127(19): 7676-7688. DOI:10.1016/j.ijleo.2016.05.115 |
| [2] |
Sheaves M, Brookes J, Coles R, Freckelton M, Groves P, Johnston R, Winberg P. Repair and revitalisation of Australias tropical estuaries and coastal wetlands: opportunities and constraints for the reinstatement of lost function and productivity. Marine Policy, 2014, 47: 23-38. DOI:10.1016/j.marpol.2014.01.024 |
| [3] |
Barbier E B, Hacker S D, Kennedy C, Koch E W, Stier A C, Silliman B R. The value of estuarine and coastal ecosystem services. Ecological monographs, 2011, 81(2): 169-193. DOI:10.1890/10-1510.1 |
| [4] |
Cai F, Su X Z, Liu J H, Li B, Lei G. Coastal erosion in China under the condition of global climate change and measures for its prevention. Progress in Natural Science, 2009, 19(4): 415-426. DOI:10.1016/j.pnsc.2008.05.034 |
| [5] |
崔丽娟, 张曼胤, 王义飞. 湿地功能研究进展. 世界林业研究, 2006, 19(3): 18-21. DOI:10.3969/j.issn.1001-4241.2006.03.004 |
| [6] |
李伟, 崔丽娟, 赵欣胜, 张曼胤, 马牧源, 康晓明, 王义飞. 中国滨海湿地及其生态系统服务功能研究概述. 林业调查规划, 2014, 39(4): 24-30. |
| [7] |
李晶, 雷茵茹, 崔丽娟, 潘旭, 张骁栋, 张曼胤, 李伟. 我国滨海滩涂湿地现状及研究进展. 林业资源管理, 2018(2): 24-28, 137-137. |
| [8] |
Tian B, Wu W T, Yang Z Q, Zhou Y X. Drivers, trends, and potential impacts of long-term coastal reclamation in China from 1985 to 2010. Estuarine, Coastal and Shelf Science, 2016, 170: 83-90. DOI:10.1016/j.ecss.2016.01.006 |
| [9] |
崔利芳, 王宁, 葛振鸣, 张利权. 海平面上升影响下长江口滨海湿地脆弱性评价. 应用生态学报, 2014, 25(2): 553-561. |
| [10] |
Blankespoor B, Dasgupta S, Laplante B. Sea-level rise and coastal wetlands. AMBIO, 2014, 43(8): 996-1005. DOI:10.1007/s13280-014-0500-4 |
| [11] |
Kirwan M L, Megonigal J P. Tidal wetland stability in the face of human impacts and sea-level rise. Nature, 2013, 504(7478): 53-60. DOI:10.1038/nature12856 |
| [12] |
Roques A. Biological invasion. Integrative Zoology, 2012, 7(3): 227-227. DOI:10.1111/j.1749-4877.2012.00311.x |
| [13] |
Ehrenfeld J G. Ecosystem consequences of biological invasions. Annual Review of Ecology, Evolution, and Systematics, 2010, 41: 59-80. DOI:10.1146/annurev-ecolsys-102209-144650 |
| [14] |
李加林, 杨晓平, 童亿勤, 张殿发, 沈永明, 张忍顺. 互花米草入侵对潮滩生态系统服务功能的影响及其管理. 海洋通报, 2005, 24(5): 33-38. DOI:10.3969/j.issn.1001-6392.2005.05.006 |
| [15] |
Díaz S, Kattge J, Cornelissen J H C, Wright I J, Lavorel S, Dray S, Reu B, Kleyer M, Wirth C, Prentice I C, Garnier E, Bönisch G, Westoby M, Poorter H, Reich P B, Moles A T, Dickie J, Gillison A N, Zanne A E, Chave J, Joseph W S, Sheremet'ev S N, Jactel H, Baraloto C, Cerabolini B, Pierce S, Shipley B, Kirkup D, Casanoves F, Joswig J S, Günther A, Falczuk V, Rüger N, Mahecha M D, Gorné L D. The global spectrum of plant form and function. Nature, 2016, 529(7585): 167-171. DOI:10.1038/nature16489 |
| [16] |
Violle C, Navas M L, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E. Let the concept of trait be functional!. Oikos, 2007, 116(5): 882-892. DOI:10.1111/j.0030-1299.2007.15559.x |
| [17] |
Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(30): 11001-11006. DOI:10.1073/pnas.0403588101 |
| [18] |
Cornelissen J H C, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich D E, Reich P B, Ter Steege H, Morgan H D, Van Der Heijden M G A, Pausas J G, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal Of Botany, 2003, 51(4): 335-380. DOI:10.1071/BT02124 |
| [19] |
Garnier E, Navas M L. A trait-based approach to comparative functional plant ecology: concepts, methods and applications for agroecology. A review. Agronomy for Sustainable Development, 2012, 32(2): 365-399. |
| [20] |
Carlucci M B, Debastiani V J, Pillar V D, Duarte L D S. Between-and within-species trait variability and the assembly of sapling communities in forest patches. Journal of Vegetation Science, 2015, 26(1): 21-31. DOI:10.1111/jvs.12223 |
| [21] |
Liu M C, Kong D L, Lu X R, Huang K, Wang S, Wang W B, Qu B, Feng Y L. Higher photosynthesis, nutrient-and energy-use efficiencies contribute to invasiveness of exotic plants in a nutrient poor habitat in northeast China. Physiologia Plantarum, 2017, 160(4): 373-382. DOI:10.1111/ppl.12566 |
| [22] |
浦瑞良, 宫鹏. 高光谱遥感及其应用. 北京: 高等教育出版社, 2000.
|
| [23] |
Aragón P, Sánchez-Fernández D, Hernando C. Use of satellite images to characterize the spatio-temporal dynamics of primary productivity in hotspots of endemic Iberian butterflies. Ecological Indicators, 2019, 106: 105449. DOI:10.1016/j.ecolind.2019.105449 |
| [24] |
童庆禧, 张兵, 郑兰芬. 高光谱遥感: 原理、技术与应用. 北京: 高等教育出版社, 2006.
|
| [25] |
王佳鹏, 施润和, 张超, 刘浦东, 曾毓燕. 基于光谱分析的长江口湿地互花米草叶片叶绿素含量反演研究. 遥感技术与应用, 2017, 32(6): 1056-1063. |
| [26] |
王林凤. 泥炭藓湿地典型植物高光谱特征研究[D]. 杭州: 杭州师范大学, 2017.
|
| [27] |
Rebelo A J, Somers B, Esler K J, Meire P. Can wetland plant functional groups be spectrally discriminated?. Remote Sensing of Environment, 2018, 210: 25-34. DOI:10.1016/j.rse.2018.02.031 |
| [28] |
Yendrek C R, Tomaz T, Montes C M, Cao Y Y, Morse A M, Brown P J, McIntyre L M, Leakey A D B, Ainsworth E A. High-throughput phenotyping of maize leaf physiological and biochemical traits using hyperspectral reflectance. Plant Physiology, 2017, 173(1): 614-626. DOI:10.1104/pp.16.01447 |
| [29] |
Gitelson A A, Merzlyak M N, Chivkunova O B. Optical properties and nondestructive estimation of anthocyanin content in plant leaves. Photochemistry and Photobiology, 2001, 74(1): 38-45. DOI:10.1562/0031-8655(2001)074<0038:OPANEO>2.0.CO;2 |
| [30] |
窦志国, 崔丽娟, 武高洁, 李晶, 潘旭, 蔡张杰, 雷茵茹, 李伟. 芦苇粉大尾蚜虫害下芦苇叶绿素高光谱反演估算. 生态学杂志, 2018, 37(10): 3163-3170. |
| [31] |
Li W, Dou Z G, Wang Y, Wu G J, Zhang M Y, Lei Y R, Ping Y M, Wang J C, Cui L J, Ma W. Estimation of above-ground biomass of reed (Phragmites communis) based on in situ hyperspectral data in Beijing Hanshiqiao Wetland, China. Wetlands Ecology and Management, 2019, 27: 87-102. DOI:10.1007/s11273-018-9644-5 |
| [32] |
Li W, Dou Z G, Cui L J, Wang R M, Zhao Z J, Cui S F, Lei Y R, Li J, Zhao X S, Zhai X J. Suitability of hyperspectral data for monitoring nitrogen and phosphorus content in constructed wetlands. Remote Sensing Letters, 2020, 11(5): 495-504. DOI:10.1080/2150704X.2020.1734247 |
| [33] |
Dou Z G, Cui L J, Li J, Zhu Y N, Gao C J, Pan X, Lei Y R, Zhang M Y, Zhao X S, Li W. Hyperspectral estimation of the chlorophyll content in short-term and long-term restorations of mangrove in Quanzhou Bay Estuary, China. Sustainability, 2018, 10(4): 1127. DOI:10.3390/su10041127 |
| [34] |
Dou Z G, Li Y Z, Cui L J, Pan X, Ma Q F, Huang Y L, Lei Y R, Li J, Zhao X S, Li W. Hyperspectral inversion of Suaeda salsa biomass under different types of human activity in Liaohe estuary wetland in north-eastern China. Marine and Freshwater Research, 2019, 71(4): 482-492. |
| [35] |
王靖雯, 牛振国. 基于潮位校正的盐城滨海潮间带遥感监测及变化分析. 海洋学报, 2017, 39(5): 149-160. DOI:10.3969/j.issn.0253-4193.2017.05.014 |
| [36] |
左雪燕, 窦志国, 蔡杨, 崔丽娟, 胡宇坤, 严睿, 李伟. 江苏盐城滨海湿地优势植物功能性状分析. 湿地科学与管理, 2019, 15(3): 59-62. |
| [37] |
Smith K A, Tabatabai M A. Automated instruments for the determination of total carbon, hydrogen, nitrogen, sulfur, and oxygen//Smith K A, Cresser M S, eds. Soil and Environmental Analysis: Modern Instrumental Techniques. Boca Raton, FL: CRC Press, 2003: 202-246.
|
| [38] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000.
|
| [39] |
哈布热, 张宝忠, 李思恩, 彭致功, 韩娜娜, 刘露. 基于冠层光谱特征的冬小麦植株含水率诊断研究. 灌溉排水学报, 2018, 37(10): 9-15. |
| [40] |
郭超凡, 郭逍宇. 基于可见光波段包络线去除的湿地植物叶片叶绿素估算. 生态学报, 2016, 36(20): 6538-6546. |
| [41] |
宫兆宁, 赵雅莉, 赵文吉, 林川, 崔天翔. 基于光谱指数的植物叶片叶绿素含量的估算模型. 生态学报, 2014, 34(20): 5736-5745. |
| [42] |
艾金泉, 陈文惠, 陈丽娟, 张永贺, 周毅军, 郭啸川, 褚武道. 冠层水平互花米草叶片光合色素含量的高光谱遥感估算模型. 生态学报, 2015, 35(4): 1175-1186. |
| [43] |
易秋香, 黄敬峰, 王秀珍, 钱翌. 玉米全氮含量高光谱遥感估算模型研究. 农业工程学报, 2006, 22(9): 138-143. DOI:10.3321/j.issn:1002-6819.2006.09.028 |
| [44] |
施润和, 牛铮, 庄大方. 利用高光谱数据估测植物叶片碳氮比的可行性研究. 遥感技术与应用, 2003, 18(2): 76-80. DOI:10.3969/j.issn.1004-0323.2003.02.004 |
| [45] |
郭澎涛, 苏艺, 茶正早, 林清火, 罗微, 林钊沐. 基于BP神经网络的橡胶苗叶片磷含量高光谱预测. 农业工程学报, 2016, 32(S1): 177-183. |
| [46] |
余哲修, 张超, 黄田, 张一, 罗恒春, 牛晓花. 剑湖湿地中菰的叶绿素含量高光谱估算模型研究. 湿地科学, 2018, 16(6): 742-748. |
| [47] |
郑雯, 明金, 杨孟克, 周四维, 汪善勤. 基于波段深度分析和BP神经网络的水稻色素含量高光谱估算. 中国生态农业学报, 2017, 25(8): 1224-1235. |
| [48] |
Karimi Y, Prasher S O, Madani A, Kim S. Application of support vector machine technology for the estimation of crop biophysical parameters using aerial hyperspectral observations. Canadian Biosystems Engineering, 2008, 50(7): 13-20. |
| [49] |
王晓星. 夏玉米冠层光谱特征及其生理生态参量的高光谱估算模型[D]. 杨凌: 西北农林科技大学, 2015.
|
| [50] |
张春兰, 杨贵军, 李贺丽, 汤伏全, 刘畅, 张丽妍. 基于随机森林算法的冬小麦叶面积指数遥感反演研究. 中国农业科学, 2018, 51(5): 855-867. |
| [51] |
姚雄, 余坤勇, 杨玉洁, 曾琪, 陈樟昊, 刘健. 基于随机森林模型的林地叶面积指数遥感估算. 农业机械学报, 2017, 48(5): 159-166. |
| [52] |
Cui L J, Dou Z G, Liu Z J, Zuo X Y, Lei Y R, Li J, Zhao X S, Zhai X J, Pan X, Li W. Hyperspectral inversion of Phragmites communis carbon, nitrogen, and phosphorus stoichiometry using three models. Remote Sensing, 2020, 12(12): 1998. |