 2021, Vol. 41
2021, Vol. 41文章信息
- 陈国鹏, 鲜骏仁, 俞筱押
- CHEN Guopeng, XIAN Junren, YU Xiaoya
- 林窗对岷江冷杉更新幼苗根系生物量积累与分配的影响
- Effect of forest gap on accumulations and allocations of root biomass in Abies faxoniana regeneration seedlings
- 生态学报. 2021, 41(14): 5685-5694
- Acta Ecologica Sinica. 2021, 41(14): 5685-5694
- http://dx.doi.org/10.5846/stxb202007151857
-
文章历史
- 收稿日期: 2020-07-15
- 网络出版日期: 2021-05-13





2. 四川农业大学环境学院, 成都 611130;
3. 黔南民族师范学院旅游与资源环境学院, 都匀 558000
2. College of Environmental Sciences, Sichuan Agricultural University, Chengdu 611130, China;
3. School of Tourism and Resource Environment, Qiannan Normal University for Nationalities, Duyun 558000, China
根系是植物的重要器官, 在生态系统的养分循环、地力维持和有机碳贮存等方面发挥着重要作用[1]。通常情况下, 将直径(d)小于2 mm的根称为细根, 它具有巨大的吸收表面积, 负责水分和养分的吸收, 是根系中最活跃和最敏感的部分[2-4];d ≥2 mm的称为粗根, 在碳和其他营养物质的运输与储存、锚定树木地上部分等方面发挥着重要作用[1, 5]。粗根和细根是地下生物量的主要组成部分, 资源在粗根和细根间的分配权衡是植物生活史策略的一个重要维度[5-7], 体现着植物对环境变化的适应能力。
作为森林自然更新的重要驱动力, 林窗将对林分环境产生重要影响, 如增大光合有效辐射量子通量密度、升高地表温度、增湿地表空气、改变土壤有效养分状况等[8-10], 从而对更新幼苗的形态建成产生重要影响[11-13]。更新幼苗通过调整生物量分配策略来适应冠层环境变化, 已有研究表明, 岷江冷杉(Abies faxoniana)[11]、栲(Castanopsis fargesii)[13]和木荷(Schima superba)[14]等林窗内幼苗会将更多的生物量分配到地上部分, 从而表现出比林下更低的根冠比和更高的叶质量比。林窗幼苗根系分配到有限的生物量资源后, 既要为地上部分吸收更多的水分和养分, 又要运输更多的物质并支撑更多的地上生物量, 这就要求林窗幼苗必须权衡好地下资源的分配, 实现有限资源的最优利用, 才能发挥好细根和粗根的各项功能, 保障更新物种的适应与存活。
岷江冷杉自然分布于我国甘肃南部、四川西北部和青海东南部, 是长江、黄河上游水源涵养林的旗舰物种之一, 其自然更新一直是西南亚高山生态恢复迫切需要解决的问题。相关研究已证实林窗更新是岷江冷杉林分更新的主要方式, 林窗出现改变了岷江冷杉林分的局部环境[10], 增加了岷江冷杉幼苗数量[11], 提高幼苗种群抵御外界环境随机干扰的能力[15], 增强种群优势度增长的环境容纳量和内禀增长率[16], 使幼苗具有更强的增长优势和相对稳定的生长环境。在林窗更新过程中, 岷江冷杉幼苗将优化资源分配, 权衡和协调地下资源在粗根和细根间的分配, 才能使其具有更强的种群增长优势。
本研究以川西亚高山岷江冷杉原始林自然更新的幼苗为对象, 通过测定林窗和林下幼苗粗根、细根生物量, 以期揭示幼苗生长过程中优化地下资源分配、协调粗根和细根的生长机制, 旨在回答:林窗是否影响岷江冷杉更新幼苗根系生物量的积累和分配过程?粗根和细根间的异速生长关系是否会随林分环境或个体大小发生调整?
1 材料与方法 1.1 研究区概况研究区位于四川省平武县内的王朗国家级自然保护区(32°49′—33°02′N, 103°55′—104°10′E, 海拔2300—4980 m), 保护区年平均气温2.9 ℃、年积温(≥10 ℃)1056.5 ℃、最高和最低气温分别为26.2 ℃和-17.8 ℃, 多年平均降水量859.9 mm;主要森林植被包括原始云杉(Picea spp.)冷杉(A. spp.)林、针阔混交林、桦木(Betula spp.)林和人工针叶林。主要土壤类型为山地棕壤、山地暗棕壤、亚高山草甸土、高山草甸土和高山流石滩荒漠土。原始岷江冷杉林主要分布区介于海拔2600—3500 m[10]。
1.2 样地调查调查样地设置在王朗国家级自然保护区大窝凼的岷江冷杉-缺苞箭竹(Fargesia denudate)-苔藓林(33°00′11.17″N, 104°01′34.43″E, 海拔2910 m), 郁闭度0.5、平均胸径45 cm, 岷江冷杉密度1533株/hm2。伴生树种有紫果云杉(P. purpurea)、大翅色木槭(Acer mono var. macropterum)和方枝圆柏(Sabina saltuaria)等。林下植物有缺苞箭竹、苦糖果(Lonicera stanishii)、悬钩子(Rubus corchorifolius)、紫花碎米荠(Cardamine tangutorum)、鹿药(Smilacina japonica)、铁线蕨(Adiantum capillus-veneris)、山羽藓(Abietinella abietina)和塔藓(Hylocomium splendens)等[11, 15]。
沿垂直于等高线的方向设置3条长300 m、宽2 m的样带, 间隔50 m以上。分林窗和林下分别测量冷杉幼苗(规定高度小于100 cm)的基径、苗高, 将每株幼苗全部挖出并做好标记后带回相距约8 km处的保护区管理局实验室, 将幼苗从基部切下, 取出根系后轻轻抖去粘在根系上的土壤, 并用流动水冲去黏附在根表面的土壤, 用干净的滤纸吸去多余水分后分粗根(d≥2 mm)和细根(d < 2 mm)在万分之一电子天平上分别称量鲜质量, 然后部分取样后装入信封袋在通风干燥箱中105 ℃杀青15 min后于80 ℃烘至恒量, 再将鲜质量换算成干质量(文中的生物量均为干质量)。共采集林下幼苗288株、林窗幼苗480株[11]。
1.3 数据分析根据全部幼苗的基径范围(1.14—13.69 mm), 以2.0 mm的间隔将幼苗划分为6个径级(I—Ⅵ), 大于13.0 mm的个别样本并入Ⅵ径级。鉴于部分径级根系生物量分布的非正态性(图 1), 用各径级中位数反映其平均状况, 并用中位数计算根系的绝对生长速率(absolute growth rate, AGR)和相对生长速率(relative growth rate, RGR)[17]。
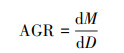
|
(1) |

|
(2) |

|
| 图 1 根系生物量随径级增粗的变化 Fig. 1 Changes of root biomass with increase of diameter classes 图中n为样本量, 穿过各箱体的“——”为生物量变化的拟合线, 箱体外的“*”、箱体中“□”和“—”分别为最大和最小值、平均值和中位数 |
式中, dM表示两径级间幼苗根系生物量的变化, dD表示幼苗径级的变化, M表示低径级幼苗根系生物量。
在SPSS 20.0中用独立样本t检验比较不同生境间各径级根系生物量和分配比例的差异, t检验过程中要先进行方差齐次性检验。在OriginPro 2018中用幂函数y=axb分别拟合林窗、林下根系生物量与分配比例随径级增大的变化规律, 不同生境间拟合方程差异的比较在FitCmpData下进行F检验。
粗根和细根生物量间异速生长关系采用logFRB=β+αlogCRB函数拟合, 式中: FRB为细根生物量, CRB为粗根生物量, α为异速生长指数, β为异速生长常数。在SMART 2.0中用标准化主轴回归(SMA)的方法拟合, 若α与1差异显著, 则表明两变量间为异速生长关系;若差异不显著, 则为等速生长关系;分别检验不同生境和不同径级间α是否存在异质性, 当α异质时, 进行多重比较;当α同质时, 进一步采用Wald显著性检验分析组间是否存在截距上的变化, 并进行多重比较[18]。所有检验的差异显著均设为P < 0.05。
2 结果分析 2.1 生物量的积累林窗幼苗细根生物量、粗根生物量和总根生物量明显小于林下幼苗(表 1), 林窗显著影响Ⅱ—Ⅴ径级幼苗粗根生物量、Ⅱ和Ⅴ径级幼苗总根生物量(表 2)。林窗、林下幼苗的粗根、细根和总根生物量随基径增粗的变化显著(图 1)。幂函数可以较好地拟合根系生物量随径级的变化(表 3), 拟合结果的F检验表明:生境极显著影响岷江冷杉幼苗细根、粗根生物量与径级的关系(P < 0.01), 显著影响总根生物量与径级间的关系(P < 0.05)。
| 项目Items | 林窗Gap | 林下Canopy | 总体All |
| 细根生物量Fine root biomass/g | 0.54(0.25, 1.20) | 0.75(0.31, 2.00) | 0.60(0.27, 1.47) |
| 粗根生物量Coarse root biomass/g | 0.50(0.24, 1.07) | 0.89(0.37, 2.24) | 0.62(0.28, 1.44) |
| 总根生物量Total root biomass/g | 5.63(2.61, 13.79) | 8.58(3.47, 20.53) | 6.38(2.73, 15.57) |
| 细根分配比例The fractional allocation to fine roots/% | 52.17(41.85, 61.78) | 45.79(37.03, 55.38) | 49.65(40.06, 59.87) |
| 粗根分配比例The fractional allocation to coarse roots/% | 47.85(38.45, 58.15) | 54.26(44.93, 63.09) | 50.56(40.28, 60.14) |
| 项目Items | 径级Classes of diameter | |||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | Ⅵ | |
| 细根生物量Fine root biomass/g | ns | ns | ns | ns | ns | ns |
| 粗根生物量Coarse root biomass/g | ns | ** | ** | * | * | ns |
| 总根生物量Total root biomass/g | ns | * | ns | ns | * | ns |
| 细根分配比例The fractional allocation to fine roots/% | ns | * | ** | ns | ns | ns |
| 粗根分配比例The fractional allocation to coarse roots/% | ns | * | ** | ns | ns | ns |
| **, P < 0.01; *, P < 0.05; ns, P>0.05 | ||||||
| 项目 Items |
生境 Habitats |
参数Parameters | R2 | F | P | |
| a | b | |||||
| 细根生物量 | 林窗 | 0.101±0.015 | 2.057±0.090 | 0.996 | 16.648 | < 0.01 |
| Fine root biomass/g | 林下 | 0.071±0.022 | 2.393±0.185 | 0.990 | ||
| 粗根生物量 | 林窗 | 0.035±0.004 | 2.869±0.064 | 0.999 | 55.487 | < 0.01 |
| Coarse root biomass/g | 林下 | 0.027±0.007 | 3.157±0.149 | 0.996 | ||
| 总根生物量 | 林窗 | 0.123±0.007 | 2.467±0.034 | 0.999 | 4.652 | < 0.05 |
| Total root biomass/g | 林下 | 0.175±0.065 | 2.330±0.219 | 0.985 | ||
全部幼苗根系生物量分配到细根和粗根的比例分别为49.65%和50.56%;林窗幼苗分配到细根和粗根的比例分别为52.17%和47.85%, 而林下幼苗分配到的比例分别为45.79%和54.26%(表 1), 林窗对Ⅱ、Ⅲ径级幼苗根系生物量的分配比例影响显著(表 2, P < 0.05)。随着岷江冷杉幼苗基径增粗, 林窗幼苗分配到细根中生物量比例从Ⅰ径级的59.20%下降到Ⅵ径级的39.87%、而粗根生物量则从41.29%增加到60.13%;林下幼苗细根生物量分配比例从Ⅰ径级的47.53%下降到Ⅵ径级的39.09%, 粗根生物量分配比例从51.92%增长到61.69%;其变化趋势均可以用幂函数较好地拟合(图 2、表 4), F检验表明:生境极显著影响岷江冷杉幼苗根系生物量分配比例随径级变化的关系(P < 0.01)。

|
| 图 2 根系生物量分配比例随径级增加的变化 Fig. 2 Changes of root biomass allocation with increase of diameter classes |
| 项目Items | 生境Habitats | 参数Parameters | R2 | F | P | |
| a | b | |||||
| 细根分配比例 | 林窗 | 60.329±2.103 | -0.189±0.030 | 0.879 | 11.06 | < 0.01 |
| The fractional allocation to fine roots/% | 林下 | 49.145±1.844 | -0.100±0.031 | 0.647 | ||
| 细根分配比例 | 林窗 | 40.291±1.484 | 0.204±0.027 | 0.922 | 13.387 | < 0.01 |
| The fractional allocation to fine roots/% | 林下 | 50.280±1.699 | 0.096±0.026 | 0.720 | ||
各径级幼苗根系的生长速率如图 3所示。随基径的增粗, 绝对生长速率多呈增加趋势, 相对生长速率呈下降趋势, 细根生长速率小于粗根生长速率。细根绝对生长速率增加至Ⅳ径级后下降, Ⅲ—Ⅴ径级林下幼苗细根绝对生长速率明显大于林窗幼苗。Ⅱ径级林窗幼苗细根相对生长速率略有上升;Ⅴ径级时林下幼苗粗根绝对生长速率明显大于林窗幼苗。

|
| 图 3 根系生物量的绝对生长速率和相对生长速率 Fig. 3 Absolute and relative growth rate of root biomass |
林窗、林下幼苗粗根和细根生物量间存在极显著的异速生长关系(图 4), 异速生长指数分别为0.969和0.985, 均与1.0无显著差异(P=0.186、0.564)。这表明岷江冷杉幼苗粗根和细根生物量间为近等速生长关系;林窗环境对异速生长指数无显著影响(P=0.172), 在共有斜率0.975下, 林窗的异速生长常数0.028显著大于林下的-0.078, 说明在同样大小的粗根生物量情况下林窗幼苗具有更高的细根生物量分配比例。

|
| 图 4 生境对幼苗粗根和细根生物量间异速生长关系的影响 Fig. 4 Allometric biomass relationships between fine root biomass and coarse root biomass in different habitats |
不同径级岷江冷杉幼苗粗根和细根生物量异速生长关系如图 5和表 5所示:除Ⅵ径级的异速生长关系不显著(P=0.073)、Ⅰ径级的异速生长指数与1.0无显著差异外(P=0.137), 其余4个径级的异速生长指数均显著大于1.0(P < 0.01)。随着径级的增粗, 异速生长指数呈增大趋势, 多重比较发现, Ⅴ径级的异速生长指数显著大于Ⅰ—Ⅲ径级。从各径级异速生长关系拟合线与1:1线的位置看(图 4):小个体岷江冷杉幼苗细根的生物量大于粗根生物量;此后随着幼苗的生长, 分配给粗根的生物量相对多于分配给细根的生物量, 这种趋势在大径级幼苗上更为突出。

|
| 图 5 不同径级幼苗粗根细根生物量异速生长关系 Fig. 5 Allometric relationships between fine root biomass and coarse root biomass in six diameter classes |
| 径级 Diameter classes |
N | R2 | 指数 Exponent |
斜率多重检验 Multiple comparison of slopes |
截距 Intercepts |
||||
| Ⅰ | Ⅱ | Ⅲ | Ⅳ | Ⅴ | |||||
| Ⅰ | 92 | 0.098** | 1.161ns(0.953, 1.415) | 1 | 0.254 | ||||
| Ⅱ | 265 | 0.327** | 1.232**(1.116, 1.361) | ns | 1 | 0.131 | |||
| Ⅲ | 194 | 0.236** | 1.404**(1.240, 1.590) | ns | ns | 1 | 0.010 | ||
| Ⅳ | 99 | 0.124** | 1.349**(1.119, 1.627) | ns | ns | ns | 1 | -0.182 | |
| Ⅴ | 52 | 0.164** | 1.864**(1.441, 2.410) | ** | ** | ** | ns | 1 | -0.607 |
| Ⅵ | 18 | 0.187ns | -0.953 | ||||||
| R2列表示回归关系的显著性, 指数列表示指数与理论值的差异显著性, 截距漂移检验列表示截距漂移量的差异显著性 | |||||||||
粗根和细根是地下净初级生产力的重要贡献者, 在森林生态系统的生物地球化学循环中发挥着重要作用[1]。但根系取样的困难一直制约着根系生态学的研究, 偏远山区森林的根系生态学研究尤其滞后。本研究采用许多学者倾向的全根系挖掘法[19], 测定了川西亚高山岷江冷杉原始林768株更新幼苗的粗根和细根生物量, 并根据基径大小划分成不同径级以反映幼苗的生长过程。结果发现:岷江冷杉幼苗粗根、细根和总根生物量随基径增粗积累明显(图 1), 根系生物量中分配到细根的比例逐渐减少、分配到粗根的比例逐渐增加(图 2), 其现象与黄荆(Vitex negundo)[20]、红松(Pinus koraiensis)[21]、思茅松(Pinus. kesiya var. langbianensis)[22]及东北天然次生林下多个树种[23]的一致, 但不同于南川木波罗(Artocarpus nanchuanensis)幼苗[24]。最优分配理论认为, 植物倾向于将资源优先分配到能获得限制性资源的器官, 以帮助植物获取更多的受限资源, 从而维持最大生长速率[25-26], 在树木个体增大的过程中, 粗根生物量分配比例的增加是为了支撑地上部分高生长的需要, 当地上部分的纵向生长基本稳定后, 粗根所占的比例下降、细根比例增加[22, 27]。本研究中岷江冷杉尚处于幼苗阶段, 纵向生长正在进行中, 细根生物量分配比例只是单调减少。而功能均衡理论认为细根和粗根的结构差异是影响其生物量分配策略调整的关键[23, 28], 细根是连接根尖的组织, 具有分生能力强、生产速度快、周转周期短的特点[1-3];而粗根比细根生产成本高、是次生分生组织的不断积累[1, 5, 29], 小径级的岷江冷杉幼苗根系发育不完全, 尤其粗根还没有发育完成, 故细根生物量所占比重较高;而随着幼苗的生长, 尽管细根和粗根的生物量均在不断增加, 但细根是不断的死亡和更新, 而粗根却在持续积累, 就呈现出细根分配比例逐渐下降、粗根的比例逐渐升高的趋势。
异速生长分配理论根据异速生长方程来检验生物量分配与个体大小的关系, 能够较好地解释物种和个体大小对生物量分配的影响, 对生物量分配理论的发展有重要意义[25, 30-31]。本研究中Ⅰ—Ⅴ径级岷江冷杉幼苗粗根-细根生物量标准化主轴估计的R2均较低。鉴于生态学研究中, 尤其是在野外实验条件下, 回归分析中R2均很低的实际[32], 可明确岷江冷杉不同径级更新幼苗的粗根生物量和细根生物量间极显著的回归关系反映着粗根和细根间权衡协变的异速生长关系。这些异速生长指数或等于1或显著大于1(图 4、5), 与东北天然次生林下树木细根生物量生长速率显著小于粗根生物量生长速率的异速生长关系显著不同(乔木型灌木、典型灌木以及混合树种粗根生物量和细根生物量间的异速生长指数分别为0.683、0.588和0.704)[23]。本研究中细根生物量与粗根生物量的异速生长指数显著大于1(表 5), 似乎与生物量分配比例(图 2)和根系绝对生长速率(图 3) 的结果不一致, 但拟合线与1∶1线的交点位置反映着异速生长关系随个体大小的调整[33], 同径级小个体岷江冷杉幼苗细根的生物量大于粗根, 随着幼苗的生长, 分配到粗根的生物量相对多于分配给细根的生物量, 且这种趋势随着幼苗的增大更为明显(图 5), 这也表明异速生长法与百分比分配法均可用于解释岷江冷杉幼苗根系生物量的分配[25]。
植物器官生物量间异速生长关系受个体大小影响明显[34-35], 异速生长指数随个体发育发生漂变反映了有机体在整个发育过程中各种功能的权衡[34], 本研究发现岷江冷杉幼苗粗根生物量和细根生物量间的异速生长关系会随径级增粗由等速生长调整为显著大于1的异速生长(图 5、表 5), 这种变化说明随着幼苗个体的增长, 单位粗根的生长需要增加更多的细根[1, 5, 33]。
3.2 林窗的影响冠层结构是影响根系生物量的重要因素[1, 36-38], 许多研究已证实林窗的出现将导致林分尺度上的“根窗效应”[36, 38], 即林窗内的根系生物量小于林下根系生物量。本研究以更新幼苗为对象, 试图揭示林窗对幼苗个体根系生物量的影响, 结果发现林窗干扰不仅影响着岷江冷杉更新幼苗粗根和细根的平均生物量, 如林窗内幼苗的平均细根生物量、粗根生物量和总根生物量均小于林下幼苗, 细根生物量分配比例大于林下(表 1), 还显著影响着细根、粗根生物量随基径的积累与分配过程(表 3、4), 林窗幼苗细根生物量分配比例的减幅大于林下幼苗(图 2)。这是由于形成林窗后光的有效性明显增加[10], 草本和灌木等下层植物带来的竞争可能会加剧[11], 这将削弱更新幼苗的生长[9, 39], 而高密度的岷江冷杉更新幼苗导致的种内竞争[16]也可能会制约到幼苗根系的生长[11], 从而导致林窗幼苗根系生物量分配比例的变幅大于林下幼苗。同时, 由于林冠层的截留作用减少了林下的土壤水分输入[40], 林窗内相对湿润的土壤可能与低的根系生物量有关[36, 38], 林窗内外生物量积累的差异反映着植物的最优分配策略[25, 31]。另外, 温度是根分生组织活动的驱动因素[36, 38], 本样地前期的研究表明, 林窗内5 cm和15 cm层土壤的日平均温度比林冠下相应深度分别高2.1℃和2.7℃[10], 进而促进了根系生物量资源向最活跃的细根部分分配;而亚高山地区林冠下的融雪引起的霜冻和较薄的雪被可能导致土壤的温度更低[38], 这将制约根系生物量向细根的分配。
林窗对不同个体大小岷江冷杉幼苗根系生物量及分配比例的影响具有阶段差异性, 这些差异表现在Ⅱ—Ⅴ径级幼苗粗根生物量、Ⅱ与Ⅴ径级的细根生物量、Ⅱ与Ⅲ径级的根系生物量分配比例(表 2)。造成这种差异的主要原因可能是不同径级幼苗个体密度的变化[21, 41-42], 本研究中林窗幼苗密度约为林下幼苗的6.45倍[15], 高密度个体导致的竞争压力和资源约束使林窗内更新幼苗相对弱小[15-16], 粗根生物量显著小于林下幼苗。Ⅱ、Ⅲ径级也是6个径级中密度最高和次高的两个径级, 个体间的竞争作用不仅影响着粗根生物量的积累也影响着根系生物量的分配比例。
从异速生长分析来看, 林窗对岷江冷杉幼苗粗根和细根生物量间的异速生长指数无显著影响(图 4), 异速生长指数与1.0无显著差异则表明林窗和林下岷江冷杉幼苗粗根和细根的生物量呈近等速生长关系。当两器官生物量间为等速生长关系时, 异速生长常数β可反映两器官生物量间的比例关系[31, 43], 在林窗和林下岷江冷杉幼苗粗根和细根生物量共有斜率与1.0无显著差异的情况下, 林窗内幼苗异速生长常数显著大于林下幼苗也预示着林窗幼苗具有更高的细根生物量分配比例。
在研究一些树木的生长过程时, 很难追踪到从苗木出土到植株死亡的真实过程, 不可能获得全生命周期的资料, 所以依靠特定时间断面上树木的各个径级组来反映其生长过程, 这种获取资料的方法称为空间替代时间的方法[44]。本研究考虑到冷杉生长缓慢、寿命长的特点, 同时鉴于林窗环境对岷江冷杉幼苗(3—20 a, 1 a定株监测)基径生长没有显著影响[10], 因此用基径增粗来表示幼苗的生长过程。这种替代方法或许不能反映出岷江冷杉幼苗最真实的完整生长过程, 如在个体生长过程中经历的环境筛较为复杂, 大小和基径关系可能一般为曲线关系, 使得个体生长过程的研究更为复杂[44]。故在未来的研究中, 探讨大小与径级间的关系及影响因素也应得到考虑。
植物生物量分配随环境可利用性资源、植株个体大小等变化而变化, 生物量分配格局的差异是植物生理和形态因素共同作用的结果[23, 25, 31]。因此, 本研究可得出:随着岷江冷杉幼苗的生长, 粗根、细根生物量不断积累, 但根系生物量中分配到粗根的比例逐渐增加、分配到细根的比例逐渐减少, 粗根-细根生物量间的异速生长指数由等速生长调整为异速生长;林窗显著影响细根、粗根生物量随基径增粗的积累与分配过程, 细根生物量分配比例随基径增粗的减幅大于林下幼苗。
| [1] |
Pransiska Y, Triadiati T, Tjitrosoedirjo S, Hertel D, Kotowska M M. Forest conversion impacts on the fine and coarse root system, and soil organic matter in tropical lowlands of Sumatera (Indonesia). Forest Ecology and Management, 2016, 379: 288-298. DOI:10.1016/j.foreco.2016.07.038 |
| [2] |
Finér L, Ohashi M, Noguchi K, Hirano Y. Factors causing variation in fine root biomass in forest ecosystems. Forest Ecology and Management, 2011, 261(2): 265-277. DOI:10.1016/j.foreco.2010.10.016 |
| [3] |
Chen G S, Hobbie S E, Reich P B, Yang Y S, Robinson D. Allometry of fine roots in forest ecosystems. Ecology Letters, 2019, 22(2): 322-331. DOI:10.1111/ele.13193 |
| [4] |
赵佳宁, 梁韵, 柳莹, 王玉珏, 杨倩茹, 肖春旺. 森林生态系统细根周转规律及影响因素. 植物学报, 2020, 55(3): 308-317. |
| [5] |
Kaushal R, Jayaparkash J, Mandal D, Kumar A, Alam N M, Tomar J M S, Mehta H, Chaturvedi O P. Canopy management practices in mulberry: impact on fine and coarse roots. Agroforestry Systems, 2019, 93(2): 545-556. DOI:10.1007/s10457-017-0148-8 |
| [6] |
燕辉, 刘广全, 李红生. 青杨人工林根系生物量、表面积和根长密度变化. 应用生态学报, 2010, 21(11): 2763-2768. |
| [7] |
程瑞梅, 王瑞丽, 肖文发, 封晓辉, 刘泽彬, 葛晓改, 王晓荣, 张炜银. 三峡库区马尾松根系生物量的空间分布. 生态学报, 2012, 32(3): 823-832. |
| [8] |
管云云, 费菲, 关庆伟, 陈斌. 林窗生态学研究进展. 林业科学, 2016, 52(4): 91-99. |
| [9] |
McGuire J P, Mitchell R J, Moser E B, Pecot S D, Gjerstad D H, Hedman C W. Gaps in a gappy forest: plant resources, longleaf pine regeneration, and understory response to tree removal in longleaf pine savannas. Canadian Journal of Forest Research, 2001, 31(5): 765-778. DOI:10.1139/x01-003 |
| [10] |
张远彬, 王开运, 鲜骏仁. 岷江冷杉林林窗小气候及其对不同龄级岷江冷杉幼苗生长的影响. 植物生态学报, 2006, 30(6): 941-946. DOI:10.3321/j.issn:1005-264X.2006.06.008 |
| [11] |
鲜骏仁, 胡庭兴, 张远彬, 王开运. 林窗对川西亚高山岷江冷杉幼苗生物量及其分配格局的影响. 应用生态学报, 2007, 18(4): 721-727. DOI:10.3321/j.issn:1001-9332.2007.04.003 |
| [12] |
马莉薇, 张文辉, 周建云, 薛瑶芹, 吴敏, 马闯. 秦岭北坡林窗大小对栓皮栎实生幼苗生长发育的影响. 林业科学, 2013, 49(12): 43-50. |
| [13] |
吴小琪, 杨圣贺, 黄力, 李笑寒, 杨超, 钱深华, 杨永川. 常绿阔叶林林冠环境对栲幼苗建成的影响. 植物生态学报, 2019, 43(1): 55-64. |
| [14] |
汤景明, 翟明普. 木荷幼苗在林窗不同生境中的形态响应与生物量分配. 华中农业大学学报, 2006, 25(5): 559-563. DOI:10.3321/j.issn:1000-2421.2006.05.023 |
| [15] |
陈国鹏, 鲜骏仁, 曹秀文, 刘锦乾, 杨永红, 雷炜. 林窗对岷江冷杉幼苗生存过程的影响. 生态学报, 2016, 36(20): 6475-6486. |
| [16] |
陈国鹏, 鲜骏仁, 李广. 林窗对岷江冷杉幼苗种群优势度增长与周期性波动的影响. 自然资源学报, 2018, 33(9): 1542-1551. |
| [17] |
Paine C E T, Marthews T R, Vogt D R, Purves D, Rees M, Hector A, Turnbull L A. How to fit nonlinear plant growth models and calculate growth rates: an update for ecologists. Methods in Ecology and Evolution, 2012, 3(2): 245-256. DOI:10.1111/j.2041-210X.2011.00155.x |
| [18] |
Warton D I, Wright I J, Falster D S, Westoby M. Bivariate line-fitting methods for allometry. Biological Reviews, 2006, 81(2): 259-291. DOI:10.1017/S1464793106007007 |
| [19] |
Addo-Danso S D, Prescott C E, Smith A R. Methods for estimating root biomass and production in forest and woodland ecosystem carbon studies: A review. Forest Ecology and Management, 2016, 359: 332-351. DOI:10.1016/j.foreco.2015.08.015 |
| [20] |
黎燕琼, 郑绍伟, 龚固堂, 陈俊华, 唐波, 朱自芳, 吴雪仙, 慕长龙. 不同年龄柏木混交林下主要灌木黄荆生物量及分配格局. 生态学报, 2010, 30(11): 2809-2818. |
| [21] |
汪金松, 范秀华, 范娟, 张春雨, 夏富才. 地上竞争对林下红松生物量分配的影响. 生态学报, 2012, 32(8): 2447-2457. |
| [22] |
贾呈鑫卓, 李帅锋, 苏建荣, 童清. 思茅松人工林根系特征与生物量分配. 生态学杂志, 2017, 36(1): 21-28. |
| [23] |
程远峰, 国庆喜, 李晓娜. 东北天然次生林下木树种的生物量器官分配规律. 生态学杂志, 2010, 29(11): 2146-2154. |
| [24] |
贺丽, 齐代华, 闵鹏, 周旭, 蒋宣斌. 不同龄级南川木波罗幼苗生物量及分配格局. 长江流域资源与环境, 2013, 22(10): 1313-1318. |
| [25] |
Poorter H. Sack L Pitfalls and possibilities in the analysis of biomass allocation patterns in plants. Frontiers in Plant Science, 2012, 3: 259. |
| [26] |
陈国鹏, 赵文智, 何世雄, 付晓. 沙柳(Salix psammophila)丛生枝生物量最优分配与异速生长. 中国沙漠, 2016, 36(2): 357-363. |
| [27] |
黄春华, 周光益, 赵厚本, 周志平, 邱治军. 广东天井山成熟杉木人工林根系生物量的测定. 中南林业科技大学学报, 2013, 33(9): 80-86. DOI:10.3969/j.issn.1673-923X.2013.09.018 |
| [28] |
Litton C M, Raich J W, Ryan M G. Carbon allocation in forest ecosystems. Global Change Biology, 2007, 13(10): 2089-2109. DOI:10.1111/j.1365-2486.2007.01420.x |
| [29] |
Loiola P P, Carvalho G H, Batalha M A. Disentangling the roles of resource availability and disturbance in fine and coarse root biomass in savanna. Austral Ecology, 2016, 41(3): 255-262. DOI:10.1111/aec.12306 |
| [30] |
Enquist B J, Niklas K J. Global allocation rules for patterns of biomass partitioning in seed plants. Science, 2002, 295: 1517-1520. DOI:10.1126/science.1066360 |
| [31] |
McCarthy M C, Enquist B J. Consistency between an allometric approach and optimal partitioning theory in global patterns of plant biomass allocation. Functional Ecology, 2007, 21(4): 713-720. DOI:10.1111/j.1365-2435.2007.01276.x |
| [32] |
Møller A P, Jennions M D. How much variance can be explained by ecologists and evolutionary biologists?, , 132(4): 492-500. Oecologia, 2002, 132(4): 492-500. DOI:10.1007/s00442-002-0952-2 |
| [33] |
Shipley B, Meziane D. The balanced-growth hypothesis and the allometry of leaf and root biomass allocation. Functional Ecology, 2002, 16(3): 326-331. DOI:10.1046/j.1365-2435.2002.00626.x |
| [34] |
Poorter H, Jagodzinski A M, Ruiz-Peinado R, Kuyah S, Luo Y, Oleksyn J, Usoltsev V A, Buckley T N, Reich P B, Sack L. How does biomass distribution change with size and differ among species? An analysis for 1200 plant species from five continents. New Phytologist, 2015, 208(3): 736-749. DOI:10.1111/nph.13571 |
| [35] |
王杨, 徐文婷, 熊高明, 李家湘, 赵常明, 卢志军, 李跃林, 谢宗强. 檵木生物量分配特征. 植物生态学报, 2017, 41(1): 105-114. |
| [36] |
Bauhus J, Bartsch N. Fine-root growth in beech (Fagus sylvatica) forest gaps. Canadian Journal of Forest Research, 1996, 26(12): 2153-2159. DOI:10.1139/x26-244 |
| [37] |
Finér L, Ohashi M, Noguchi K, Hirano Y. Fine root production and turnover in forest ecosystems in relation to stand and environmental characteristics. Forest Ecology and Management, 2011, 262(11): 2008-2023. DOI:10.1016/j.foreco.2011.08.042 |
| [38] |
Wang Y, Kim J H, Mao Z, Ramel M, Pailler F, Perez J, Rey H, Tron S, Jourdan C, Stokes A. Tree root dynamics in montane and sub-alpine mixed forest patches. Annals of Botany, 2018, 122(5): 861-872. |
| [39] |
Jokela E J, Martin T A. Effects of ontogeny and soil nutrient supply on production, allocation, and leaf area efficiency in loblolly and slash pine stands. Canadian Journal of Forest Research, 2000, 30(10): 1511-1524. DOI:10.1139/x00-082 |
| [40] |
巩合德, 王开运, 杨万勤, 王乾, 张远彬. 川西亚高山原始云杉林内降雨分配研究. 林业科学, 2005, 41(1): 198-201. DOI:10.3321/j.issn:1001-7488.2005.01.033 |
| [41] |
高祥, 丁贵杰, 翟帅帅, 陈模芳, 杜华东. 不同林分密度马尾松人工林根系生物量及空间分布研究. 中南林业科技大学学报, 2014, 34(6): 71-75. DOI:10.3969/j.issn.1673-923X.2014.06.014 |
| [42] |
李帅锋, 贾呈鑫卓, 杨利华, 黄小波, 郎学东, 刘万德, 苏建荣. 林分密度对思茅松人工林根系生物量空间分布的影响. 西北植物学报, 2017, 37(11): 2265-2272. DOI:10.7606/j.issn.1000-4025.2017.11.2265 |
| [43] |
Cheng D L, Ma Y Z, Zhong Q L, Xu W F. Allometric scaling relationship between above- and below-ground biomass within and across five woody seedlings. Ecology and Evolution, 2014, 4(20): 3968-3977. DOI:10.1002/ece3.1184 |
| [44] |
何亚平, 费世民, 蒋俊明, 陈秀明, 张旭东, 何飞. 不同龄级划分方法对种群存活分析的影响-以水灾迹地油松和华山松种群生存分析为例. 植物生态学报, 2008, 32(2): 448-455. DOI:10.3773/j.issn.1005-264x.2008.02.024 |