 2021, Vol. 41
2021, Vol. 41文章信息
- 潘英杰, 何志瑞, 刘玉林, 董凌勃, 吕文文, 上官周平, 邓蕾
- PAN Yingjie, HE Zhirui, LIU Yulin, DONG Lingbo, Lü Wenwen, SHANGGUAN Zhouping, DENG Lei
- 黄土高原天然次生林植被演替过程中土壤团聚体有机碳动态变化
- Dynamics of soil aggregate-associated organic carbon during secondary forest succession in the Loess Plateau, China
- 生态学报. 2021, 41(13): 5195-5203
- Acta Ecologica Sinica. 2021, 41(13): 5195-5203
- http://dx.doi.org/10.5846/stxb202101180187
-
文章历史
- 收稿日期: 2021-01-18
- 修订日期: 2021-04-19





2. 中国科学院大学研究生院, 北京 100049;
3. 甘肃省子午岭林业管理局合水分局连家砭林场, 合水 745000;
4. 西北农林科技大学水土保持研究所, 杨凌 712100
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Lianjiabian Forest Farm, Ziwuling Forestry Administration, Heshui 745000, China;
4. Institute of Soil and Water Conservation, Northwest A&F University, Yangling 712100, China
土地利用变化是影响土壤碳“源/汇”功能转变的关键因素之一[1], 能够改变土壤团聚体稳定性、土壤有机碳在团聚体中的分布、植物残体的分解能力、土壤微生物群落等[2-3]。土壤总有机碳(TOC)在较短时间内对土地利用变化引起的改变不甚敏感, 并且对总有机碳的测定很难准确、及时地反映土壤碳库的内在变化[4], 而土壤团聚体有机碳作为土壤碳库的组成之一, 能够较好地反映土地利用与管理对土壤有机质数量和质量的影响, 其对土地利用变化的响应更为敏感[2, 5]。基于土壤有机碳在不同团聚体中的差异, 阐明土壤团聚体内有机碳含量和分布特征, 对于揭示因土地利用变化而引起的土壤有机碳变化动态和稳定性机制具有重要的意义。
植被演替是土地利用的主要变化过程, 它是在自然条件下的植被恢复过程, 在这个过程中, 地上植被逐步变化, 地下碳积累也随之发生改变, 从而造成不同演替阶段TOC积累速率明显不同[6]。不同土地利用类型的土壤有机质因凋落物的数量和质量以及环境的不同而发生变化, 影响土壤的有机碳含量以及团聚体中有机碳的稳定性和含量[7]。植被演替通过加速植物群落发展和提高土壤有机质输入, 进而促进土壤团聚体的形成[8-9]和提高团聚体稳定性[10]。团聚体在有机碳的储存、供应及转化能力等方面发挥着重要作用, 其有机碳含量显著影响土壤的养分保持能力和碳固存[11]。不同粒径团聚体的有机碳含量可以用来微观表征有机质与有机碳矿化速率之间的平衡。目前国内外关于植被恢复对土壤团聚体有机碳的影响主要存在两种观点。大多数认为植被恢复首先增加大团聚体的有机碳, 随后微团聚体有机碳也逐渐地增加, 而且植被恢复中大团聚体有机碳累积速度快于微团聚体以及粉粒与黏粒结合态有机碳的累积速度[2, 12]。但是, 也有研究发现植被恢复首先增加土壤微团聚体有机碳, 并且微团聚体有机碳对TOC贡献较大[13]。然而, 现有研究多集中在短期的植被恢复过程[2, 14], 而对长期植被恢复过程(如:植被演替过程)的研究较少。
子午岭是黄土高原目前保存较完好的天然次生植被区, 经过长期的恢复和保护, 在该区形成了时间跨度大(约160a)、保存完整(从弃耕地到成熟次生林)、受干扰较小的天然次生林植被演替系列[15], 为开展长期植被恢复中土壤有机碳动态及稳定机制研究提供了理想的平台。有关该区域植被演替对团聚体有机碳影响的报道较少, 且尚不清楚长期植被演替过程中, 植物、微生物等对土壤团聚体有机碳的影响。因此, 本文以天然次生林植被演替序列为研究对象, 探究了天然次生林植被演替过程中土壤团聚体有机碳的动态变化, 以及分析凋落物、根系、微生物和团聚体稳定性等与团聚体有机碳的关系, 以期明确长期植被恢复中土壤团聚体碳库动态与影响因素, 为深入认识黄土高原植被恢复中土壤有机碳库稳定性机制提供科学依据。
1 材料与方法 1.1 研究区概况研究区位于甘肃省合水县连家砭林场的子午岭林区, 属于北温带半湿润半干旱地区, 年平均气温10 ℃, 年均降雨量为587 mm, 主要集中在7—9月。研究区的海拔在1211—1453 m之间。土壤大部分是石灰性褐土, 由原生或次生的黄土形成, 厚度为50—130 m。该区为温凉半干旱区黄土覆盖的森林草原地带, 适于山杨(Populus davidiana)、辽东栎(Quercus liaotungensis)等落叶林和温性针叶林发育, 灌木以沙棘(Hippophae rhamnoides)为主, 草本植物以白羊草(Bothriochloa ischaemum)、茭蒿(Artemisia giraldii)、达乌里胡枝子(Lespedeza davurica)等为主(表 1)。根据历史资料记载和咨询当地土地使用历史情况, 农田弃耕后按照自然演替序列, 演替到草地(白羊草), 灌木(沙棘)、先锋林(山杨)和顶级林(辽东栎)时, 大约需要的演替时间分别为30、60、90、160a[3, 6, 15]。
| 地点 Location |
农田 Cropland |
草地 Grassland |
灌木 Shrub |
先锋林 Pioneer forest |
顶级林 Climax forest |
| 经纬度/(°)Latitude and longitude | 108.464—108.473 E36.068—36.078 N | 108.521—108.539 E36.072—36.090 N | 108.521—108.529 E36.083—36.089 N | 108.527—108.534 E36.044—36.055 N | 108.535—108.540 E36.047—36.051 N |
| 海拔Altitude/m | 1474—1476 | 1319—1323 | 1352—1354 | 1450—1455 | 1440—1450 |
| 盖度Coverage/% | — | 73.3±7.07 | 79.7±2.5 | 88.7±4.04 | 82.2±3.79 |
| pH | 8.70±0.18 | 8.57±0.08 | 8.31±0.08 | 8.35±0.14 | 8.34±0.12 |
| 含水量Water content/% | 11.473±0.75 | 18.547±1.94 | 15.857±1.97 | 13.222±1.04 | 14.422±1.40 |
| 凋落物生物量/(g/kg)Litter biomass | 175.06±8.24 | 39.71±13.00 | 124.54±25.30 | 144.44±27.83 | 231.55±33.09 |
| 微生物生物量碳/(g/kg)Microbial biomass carbon | 1.51±0.58 | 2.47±0.35 | 4.94±0.71 | 5.49±0.97 | 4.56±0.68 |
| 根系生物量/(g/kg)Root biomass | 92.57±16.68 | 126.50±25.04 | 133.58±26.82 | 296.22±36.07 | 409.77±47.01 |
选择研究区天然次生林植被演替序列为对象, 按照农田、草地、灌木、先锋林、顶级林演替序列, 分别选择各演替阶段代表性的植被类型:玉米地、白羊草、沙棘、山杨、辽东栎。2020年9月在每个植被演替阶段, 分别选择3个20 m×20 m样地, 每个样地随机选取3个1 m×1 m的样方, 选取海拔、坡度、坡向和环境基本相同的样地, 以减小实验样地带来的误差。收集样方内的未分解的凋落物, 每个样方随机钻取3个土壤样品(0—20 cm, 直径9 cm), 过2 mm筛, 去除杂物混合成一份样品, 用于测土壤理化性质, 同时分拣土壤样品中的根系, 水洗烘干后用于计算根系生物量。从每个样方周围1 m范围内的0—20 cm土层中取原状土样, 自然风干后进行团聚体稳定性分析以及土壤团聚体有机碳的测定。
1.3 测定方法土壤团聚体采用分级湿筛法[16], 收集4个团聚体粒级>2 mm、2—0.25 mm、0.25—0.053 mm和 < 0.053 mm。将土壤团聚体稳定性指标采用平均重量直径(MWD)、几何平均直径(GMD)和水稳性团聚体百分比(WAS)。分别按照以下公式计算[17]:
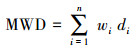
|
(1) |
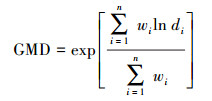
|
(2) |
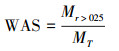
|
(3) |
其中di是筛分出来的任意粒径范围内微团聚体的平均直径(mm);wi是任意粒径范围内微团聚体的质量占土样干重的百分比(%)。Mr> 0.25为>0.25 mm水稳定性团聚体质量(g);MT为团聚体总量(g)。
采用pH-3C型酸度计测定了土壤pH(土水比1∶5)。TOC含量、团聚体有机碳含量(SAOC)通过K2Cr2O7-H2SO4氧化法测定[18]。土壤微生物生物量碳(MBC)采用改进的氯仿熏蒸K2SO4浸提法测定[19]。计算公式:
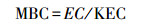
|
其中MBC表示微生物生物量碳质量分数(mg/kg);EC表示熏蒸土样有机碳量与未熏蒸土样有机碳之差(mg/kg);KEC表示微生物体中碳被浸提出来的比例, 取0.45。
1.4 数据分析利用SPSS 25.0对不同植被演替阶段土壤团聚体有机碳含量、MWD、GMD、WAS、TOC含量进行方差分析(ANOVA)。采用皮尔逊相关系数评价不同因子间的相关关系, 采用逐步回归分析影响不同粒径团聚体有机碳的主要因素。采用Origin 2019进行绘图。
2 结果分析 2.1 植被演替中土壤团聚体有机碳动态变化不同植被演替阶段各粒径土壤团聚体有机碳含量差异极显著(P < 0.001)。2—0.25 mm粒径SAOC最高, < 0.053 mm粒径SAOC最低(图 1)。除草地0.25—0.053 mm粒径SAOC之外, 随着植被演替各粒径SAOC均呈现上升趋势。在顶级林中, > 2 mm、2—0.25 mm、0.25—0.053 mm和 < 0.053 mm的各粒径土壤有机碳含量比农田分别高51.2%、53.7%、56.3%和45.6%。

|
| 图 1 不同植被演替阶段土壤团聚体有机碳含量变化 Fig. 1 Change of aggregate-associated organic carbon in different vegetation succession stages 相同小写字母表示差异不显著(P > 0.05) |
植被演替中TOC含量以顶级林最高, 比农田、草地、灌木、先锋林样地土壤分别高51.2%、56.5%、35.9%和10.3%。除草地以外, 各植被演替阶段TOC含量显著高于农田(P < 0.05), TOC呈现随植被演替不断增加的趋势(图 2)。

|
| 图 2 不同植被演替阶段土壤有机碳含量变化 Fig. 2 Changes of soil organic carbon content in different vegetation succession stages 相同小写字母表示差异不显著(P > 0.05) |
植被演替中, 除先锋林外, > 2 mm粒径的团聚体占比均随植被演替不断增加, 顶级林达到最高值。2—0.5 mm、0.25—0.053 mm、< 0.053 mm均表现为减小(图 3)。顶级林土壤团聚体的MWD、GMD和WAS值最高, 分别比农田高51.0%、15.7%和14.6%, 这表明随着植被演替, 团聚体稳定性得到显著提高。

|
| 图 3 不同植被演替阶段团聚体粒径组成和稳定性动态变化 Fig. 3 Dynamic changes of aggregate size composition and stability in different vegetation succession stages 相同小写字母表示差异不显著(P > 0.05) |
皮尔逊相关分析表明(表 2), 各粒径SAOC均与团聚体稳定性、TOC、MBC、根系生物量、凋落物生物量呈显著性正相关(P < 0.05)。各粒径SAOC与TOC的相关系数表现为(> 2 mm)>(2—0.25 mm)>(< 0.053 mm)>(0.25—0.053 mm)。除0.25—0.053 mm粒径SAOC以外, 与MBC的相关系数范围为(0.314—0.344);根系生物量与>2 mm粒径SAOC的相关系数最大(0.686), 凋落物生物量与0.25—0.053 mm粒径SAOC的相关系数最大(0.669)。
| 团聚体有机碳 Aggregate-associated organic carbon |
演替阶段 Succession stage |
MWD | GMD | WAS | TOC | MBC | LBC | RBC |
| > 2 mm | 0.726** | 0.398** | 0.253* | 0.347** | 0.673** | 0.341** | 0.686** | 0.415** |
| 2—0.25 mm | 0.747** | 0.431** | 0.334** | 0.371** | 0.589** | 0.344 | 0.670** | 0.449** |
| 0.25—0.053 mm | 0.645** | 0.358** | 0.287** | 0.333** | 0.512** | 0.174** | 0.635** | 0.669** |
| < 0.053 mm | 0.649** | 0.535** | 0.357** | 0.455** | 0.568** | 0.314** | 0.587** | 0.349** |
| **表示在0.01水平差异显著;*表示在0.05水平差异显著(双尾), n=45;MWD: Mean weight diameter平均重量直径;GMD: geometric mean diameter几何平均直径;WAS: Water stable aggregates水稳性团聚体百分比;TOC: total organic carbon总有机碳;MBC: microbial biomass carbon微生物生物量碳;LB: litter biomass凋落物生物量;RB: root biomass根系生物量 | ||||||||
由于MBC与TOC存在共线性, 所以剔除MBC后, 进行逐步回归, 得出最优的多元线性回归模型(表 3)。结果表明, 植被演替阶段(恢复时间)是各粒径SAOC的主要影响之一, 且相对贡献最大。除此之外, 影响>2 mm粒径SAOC的主要因素还包括总有机碳和根系生物量, 总有机碳的贡献更大;影响2—0.25 mm粒径SAOC的主要因素还包括总有机碳;影响0.25—0.053 mm粒径SAOC的主要因素还包括凋落物生物量;影响 < 0.053 mm粒径SAOC的主要因素还包括TOC和MWD。
| 团聚体有机碳 Aggregate-associated organic carbon |
方程 Equation |
R2 | F | P | n |
| > 2 mm | y=0.024 x1+0.316 x5 +0.007 x8+8.253 | 0.658 | 58.181 | 0.000 | 45 |
| 2—0.25 mm | y=0.049 x1+0.172 x5+12.349 | 0.652 | 84.282 | 0.000 | 45 |
| 0.25—0.053 mm | y=0.05 x1+0.023 x7+11.364 | 0.686 | 98.115 | 0.000 | 45 |
| < 0.053 mm | y=0.017 x1+0.194 x5+1.688 x2+6.008 | 0.536 | 35.269 | 0.000 | 45 |
| x1为演替阶段;x2为MWD (Mean weight diameter) 平均重量直径;x3GMD (geometric mean diameter) 几何平均直径;x4WAS (Water stable aggregates) 水稳性团聚体百分比;x5TOC (total organic carbon) 总有机碳;x6为MBC (microbial biomass carbon) 微生物生物量碳;x7为LB (litter biomass) 凋落物生物量;x8为RB (root biomass) 根系生物量 | |||||
土壤团聚化是土壤固碳的重要途径[20]。本研究发现, 与农田相比, 植被演替中各粒级团聚体有机碳显著提高(P < 0.05);其中不同植被演替阶段各粒级SAOC之间差异均达显著水平。不同植被演替阶段由于有机物料的输入和输出差异, 导致了其土壤物理性状和微生物活性不同, 从而对土壤有机碳在各粒径团聚体分布产生影响[21-23]。
本研究发现植被演替后土壤TOC含量远高于农田, 表明弃耕后土壤质量有所改善。TOC含量的变化可能是由于林地凋落物等的积累, 增加了土壤中的胶结物质, 进而促进团聚体的形成与稳定, 并增加了有机碳的固存[24-25]。与农田相比, 草地TOC较低, 这可能是因为弃耕后减少了对土地的管理, 土壤孔隙变少且不进行施肥, 草地自身的凋落物等残体的分解并不能满足其养分循环的需求, 再加上土壤微生物的数量增加对养分的需求量也会增加, 这些原因都有可能降低TOC的含量[26]。
本研究中, 各粒径SAOC与TOC显著正相关(P < 0.05), > 2 mm粒径SAOC与土壤总有机碳相关系数最大。> 2 mm粒径SAOC的变化最大, 这可能是随着植被演替增新鲜植物残体有机碳含量增加, 更多的有机碳被大团聚体保护起来, 进而促进了有机碳在土壤中的固定[27]。顶级林0.25—0.053 mm粒径SAOC显著高于其他粒径原因可能在于:一是植被演替到先锋林后团聚体粒径组成无显著区别(P > 0.05), 团聚体达到较稳定状态, > 2 mm、2—0.25 mm团聚体透气性较好, 有机碳周转速度快, 碳固存较少[28];二是较小团聚体内的碳组分可能更难分解, 但由于 < 0.053 mm团聚体含量太少, 表现不明显。
3.2 植被演替中团聚体稳定性、凋落物和根系对土壤团聚体有机碳动态的影响土壤团聚体MWD、GMD和WAS的值越大, 土壤结构越稳定, 土壤抗侵蚀能力越强[29]。通过对团聚体有机碳和团聚体稳定性的相关分析(表 2), 发现团聚体稳定性与团聚体有机碳含量显著正相关, 团聚体稳定性指数与 < 0.053 mm粒径SAOC的相关系数最大, 与0.25—0.053 mm粒径SAOC的相关系数最小。团聚体形成过程中, 有机碳是重要的胶结物质, 大团聚体分解使得有机碳进入小团聚体, 随植被演替团聚体趋于稳定, < 0.053 mm粒径SAOC变化趋势与其相似, 而0.25—0.053 mm粒径SAOC在先锋林、顶级林阶段出现显著上升。原因可能在于到了演替后期, 团聚体的形成与分解趋于平衡, 碳倾向于由大团聚体向小团聚体的流动[30]。
凋落物和根系生物量在植被演替过程中显著增加(P < 0.001)。凋落物生物量与团聚体有机碳显著正相关(P < 0.001), 与0.25—0.053 mm粒径SAOC的相关系数最大(0.669), 与 < 0.053 mm粒径SOAC最小(0.587);根系生物量与团聚体有机碳显著正相关(P < 0.001), 与>2 mm粒径SAOC的相关系数最大(0.686), 与 < 0.053 mm粒径SAOC最小(0.349), 这与前人的研究结果一致[31]。顶级林的枯枝落叶的质量和数量大于先锋林、灌木、草本和农田。大量枯枝落叶进入土壤, 为土壤微生物提供了充足的碳源, 微生物活动促进了土壤中有机碳的转化和分解[3, 32]。植物类型不同对微生物活性和群落结构的刺激作用不同, 对SAOC的作用也不尽相同。随着植被演替, 各粒径SAOC含量不断增加, 这可能与凋落物、根系残体等有机质的投入量增加有关[3]。大团聚体中的有机碳比微团聚体中的有机碳更容易矿化[21], 而微团聚体中的有机碳则大多是高度腐殖化的惰性组分。关于团聚体形成的经典模型, 认为新鲜输入的有机碳进入土壤后, 首先与土壤中粒径较小的微团聚体胶结形成大团聚体, 微团聚体再形成于其内部, 而存在于微团聚体中的有机碳由于受到物理保护, 所以其稳定性比大团聚体中的新鲜有机质更高[33]。另外, 土壤团聚体的形成可阻止空气和水的扩散, 从而阻止团聚体内有机碳的分解[34], 而具有较小孔隙的微团聚体对有机碳稳定性的贡献更大[5]。
3.3 植被演替中土壤微生物对土壤团聚体有机碳动态的影响土壤MBC随着植被演替不断升高的趋势, 与团聚体有机碳的变化趋势相同。植物的生长对MBC有着直接和间接的影响:一方面根系分泌物和凋落物的输入为微生物活动提供碳源[6];另一方面可以通过改变土壤结构和营养状况间接的改变微生物的生存环境[3]。植被演替过程中凋落物生物量、根系生物量以及根系分泌物等会发生改变, 进而会影响微生物活性和数量, 导致MBC发生显著变化[35]。本研究中, 除0.25—0.053 mm团聚体有机碳之外, 其余粒径团聚体有机碳含量均与MBC呈显著正相关, 其中MBC与0.25—0.053 mm团聚体的相关系数最大(0.344)。土壤微生物及其代谢产物促进土壤内不同大小团聚体结构的形成[36]。植被演替有助于改变植物和土壤性质, 并导致微生物多样性产生变化[37], 这体现在植被演替后期土壤微生物多样性的增加。土壤微生物多样性越大, 土壤有机碳组分的周转速度越快[39]。大团聚体由微团聚体与有机物等胶结剂胶结形成, 含有许多不稳定的有机物如轻组碳及微生物量碳等, 而易于被微生物利用发生矿化分解[3]。在植被演替过程中, SAOC与MBC相互促进, 使土壤团聚体肥力与健康状况逐渐改善[3, 5]。
4 结论从农田到顶级林(辽东栎)的长期植被演替过程中, 土壤团聚体稳定性、TOC和各粒径团聚体有机碳含量均随植被演替时间的增加而增加, 表明随着植被恢复土壤结构趋于稳定, 含碳量增加, 土壤质量得以改善。团聚体稳定性、根系、凋落物、MBC与团聚体有机碳含量显著正相关, 且不同粒径团聚体有机碳的主要影响因子存在差异, 其中, 影响>0.053 mm粒径团聚体有机碳的主要因素为植物生物量(包括:凋落物生物量、根系生物量), 而 < 0.053 mm粒径团聚体有机碳主要受团聚体本身特性的影响。因此, 植被恢复有利于团聚体的稳定性和土壤有机碳的保护, 通过长期植被恢复可以有效地改善黄土高原地区的土壤质量, 提高土壤碳储量。
| [1] |
Mahowald N M, Randerson J T, Lindsay K, Munoz E, Doney S C, Lawrence P, Schlunegger S, Ward D S, Lawrence D, Hoffman F M. Interactions between land use change and carbon cycle feedbacks. Global Biogeochemical Cycles, 2017, 31(1): 96-113. DOI:10.1002/2016GB005374 |
| [2] |
Panettieri M, Rumpel C, Dignac M F, Chabbi A. Does grassland introduction into cropping cycles affect carbon dynamics through changes of allocation of soil organic matter within aggregate fractions. Science of the Total Environment, 2017, 576: 251-263. DOI:10.1016/j.scitotenv.2016.10.073 |
| [3] |
Deng L, Kim D G, Peng C H, Shangguan Z P. Controls of soil and aggregate-associated organic carbon variations following natural vegetation restoration on the Loess Plateau in China. Land Degradation & Development, 2018, 29(11): 3974-3984. |
| [4] |
Debnath S, Attri B L, Kumar A, Kishor A, Narayan R, Sinha K, Bhowmik A, Sharma A, Singh D B. Influence of peach (Prunus persica Batsch) phenological stage on the short-term changes in oxidizable and labile pools of soil organic carbon and activities of carbon-cycle enzymes in the North-Western Himalayas. Pedosphere, 2020, 30(5): 638-650. DOI:10.1016/S1002-0160(20)60026-1 |
| [5] |
Zhu G Y, Shangguan Z P, Deng L. Dynamics of water-stable aggregates associated organic carbon assessed from delta C-13 changes following temperate natural forest development in China. Soil & Tillage Research, 2021, 205: 104782. |
| [6] |
Deng L, Wang K B, Tang Z S, Shangguan Z P. Soil organic carbon dynamics following natural vegetation restoration: evidence from stable carbon isotopes (δ13C). Agriculture, Ecosystems & Environment, 2016, 221: 235-244. |
| [7] |
Luan H A, Gao W, Tang J W, Li R N, Li M Y, Zhang H Z, Chen X P, Masiliunas D, Huang S W. Aggregate-associated changes in nutrient properties, microbial community and functions in a greenhouse vegetable field based on an eight-year fertilization experiment of China. Journal of Integrative Agriculture, 2020, 19(10): 2530-2548. DOI:10.1016/S2095-3119(20)63269-5 |
| [8] |
Gabarrón-Galeote M A, Martínez-Murillo J F, Quesada M A, Ruiz-Sinoga J D. Seasonal changes in the soil hydrological and erosive response depending on aspect, vegetation type and soil water repellency in different Mediterranean microenvironments. Solid Earth, 2013, 4(2): 497-509. DOI:10.5194/se-4-497-2013 |
| [9] |
Mekonnen M, Keesstra S D, Stroosnijder L, Baartman J E M, Maroulis J. Soil conservation through sediment trapping: a review. Land Degradation & Development, 2015, 26(6): 544-556. |
| [10] |
Burri K, Graf F, Böll A. Revegetation measures improve soil aggregate stability: a case study of a landslide area in Central Switzerland. Forest Snow and Landscape Research, 2009, 82(1): 45-60. |
| [11] |
张顺涛, 任涛, 周橡棋, 方娅婷, 廖世鹏, 丛日环, 鲁剑巍. 油/麦-稻轮作和施肥对土壤养分及团聚体碳氮分布的影响. 土壤学报, 2021. DOI:10.11766/trxb202004090091 |
| [12] |
Cheng M, Xiang Y, Xue Z J, An S S, Darboux F. Soil aggregation and intra-aggregate carbon fractions in relation to vegetation succession on the Loess Plateau, China. Catena, 2015, 124: 77-84. DOI:10.1016/j.catena.2014.09.006 |
| [13] |
Tang F K, Cui M, Lu Q, Liu Y G, Guo H Y, Zhou J X. Effects of vegetation restoration on the aggregate stability and distribution of aggregate-associated organic carbon in a typical karst gorge region. Solid Earth, 2016, 7(1): 141-151. DOI:10.5194/se-7-141-2016 |
| [14] |
Wu J J, Zhang Q, Yang F, Lei Y, Zhang Q F, Cheng X L. Afforestation impacts microbial biomass and its natural 13C and 15N abundance in soil aggregates in central China. Science of the Total Environment, 2016, 568: 52-56. DOI:10.1016/j.scitotenv.2016.05.224 |
| [15] |
王凯博, 陈美玲, 秦娟, 刘勇, 安慧, 上官周平. 子午岭植被自然演替中植物多样性变化及其与土壤理化性质的关系. 西北植物学报, 2007, 27(10): 2089-2096. DOI:10.3321/j.issn:1000-4025.2007.10.025 |
| [16] |
Elliott E T. Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Science Society of America Journal, 1986, 50(3): 627-633. DOI:10.2136/sssaj1986.03615995005000030017x |
| [17] |
Kemper W D, Rosenau R C. Aggregate stability and size distributions. Klute A, ed. Methods of Soil Analysis. Part Ⅰ. Agronomy Monograph. Madison: American Society of Agronomy, 1986: 424-442.
|
| [18] |
鲍士旦. 土壤农化分析 (第三版). 北京: 中国农业出版社, 2000: 30-34.
|
| [19] |
Vance E D, Brookes P C, Jenkinson D S. An extraction method for measuring soil microbial biomass C. Soil Biology and Biochemistry, 1987, 19(6): 703-707. DOI:10.1016/0038-0717(87)90052-6 |
| [20] |
Lal R, Kimble J M. Conservation tillage for carbon sequestration. Nutrient Cycling in Agroecosystems, 1997, 49(1-3): 243-253. |
| [21] |
罗友进, 魏朝富, 李渝, 任镇江, 廖和平. 土地利用对石漠化地区土壤团聚体有机碳分布及保护的影响. 生态学报, 2011, 31(1): 257-266. |
| [22] |
马瑞萍, 安韶山, 党廷辉, 戴相林. 黄土高原不同植物群落土壤团聚体中有机碳和酶活性研究. 土壤学报, 2014, 51(1): 104-113. |
| [23] |
郝翔翔, 窦森, 韩晓增, 李明敏, 安丰华. 典型黑土区不同生态系统下土壤团聚体中胡敏酸的结构特征. 土壤学报, 2014, 51(4): 824-833. |
| [24] |
Li H, Shen H H, Chen L Y, Liu T Y, Hu H F, Zhao X, Zhou L H, Zhang P J, Fang J Y. Effects of shrub encroachment on soil organic carbon in global grasslands. Scientific Reports, 2016, 6(1): 28974. DOI:10.1038/srep28974 |
| [25] |
Novara A, Rühl J, La Mantia T, et al. Litter contribution to soil organic carbon in the agriculture abandons processes. Solid Earth Discussions, 2015, 7(1): 595-616. |
| [26] |
任荣秀, 杜章留, 孙义亨, 宋学姝, 陆森. 华北低丘山地不同土地利用方式下土壤团聚体及其有机碳分布特征. 生态学报, 2020, 40(19): 6991-6999. |
| [27] |
贺同鑫, 胡宝清, 张建兵, 张诗萌, 庞榆, 裴广廷, 胡刚, 张伟, 孙建飞. 植被恢复十年喀斯特坡地细根对土壤碳氮存留与可利用性的影响. 生态学报, 2020, 40(23): 8638-8648. |
| [28] |
Six J, Elliott E T, Paustian K. Soil macroaggregate turnover and microaggregate formation: a mechanism for C sequestration under no-tillage agriculture. Soil Biology & Biochemistry, 2000, 32(14): 2099-2103. |
| [29] |
丁文峰, 丁登山. 黄土高原植被破坏前后土壤团粒结构分形特征. 地理研究, 2002, 21(6): 700-706. DOI:10.3321/j.issn:1000-0585.2002.06.005 |
| [30] |
Li J Y, Yuan X L, Ge L, Li Q, Li Z G, Wang L, Liu Y. Rhizosphere effects promote soil aggregate stability and associated organic carbon sequestration in rocky areas of desertification. Agriculture, Ecosystems & Environment, 2020, 304: 107126. |
| [31] |
Barthès B, Roose E. Aggregate stability as an indicator of soil susceptibility to runoff and erosion; validation at several levels. Catena, 2002, 47(2): 133-149. DOI:10.1016/S0341-8162(01)00180-1 |
| [32] |
De Gryze S, Six J, Brits C, Merckx R. A quantification of short-term macroaggregate dynamics: influences of wheat residue input and texture. Soil Biology & Biochemistry, 2005, 37(1): 55-66. |
| [33] |
Six J, Elliott E T, Paustian K. Aggregate and soil organic matter dynamics under conventional and no-tillage systems. Soil Science Society of America Journal, 1999, 63(5): 1350-1358. DOI:10.2136/sssaj1999.6351350x |
| [34] |
Ullah R, Lone M I, Ullah K S, Mehdi S M, Qazi M A. Effect of cropping systems and seasonal variations on soil microbial biomass and enzymatic activities in arid soils. The Journal of Animal & Plant Sciences, 2013, 23(2): 493-499. |
| [35] |
董建新, 丛萍, 刘娜, 李玉义, 王婧, 逄焕成. 秸秆深还对黑土亚耕层土壤物理性状及团聚体分布特征的影响. 土壤学报, 2021, 58(4): 1-15. |
| [36] |
Hoogmoed M, Cunningham S C, Baker P J, Beringer J, Cavagnaro T R. Effects of wetting frequency and afforestation on carbon, nitrogen and the microbial community in soil. Agriculture, Ecosystems & Environment, 2016, 231: 34-43. |
| [37] |
Liu Y L, Zhu G Y, Hai X Y, Li J W, Shangguan Z P, Peng C H, Deng L. Long-term forest succession improves plant diversity and soil quality but not significantly increase soil microbial diversity: evidence from the Loess Plateau. Ecological Engineering, 2020, 142: 105631. DOI:10.1016/j.ecoleng.2019.105631 |
| [38] |
吴浩浩, 徐星凯, 段存涛, 李团胜, Cheng W G. 森林土壤融化期异养呼吸和微生物碳变化特征. 生态学报, 2017, 37(8): 2729-2742. |