 2021, Vol. 41
2021, Vol. 41文章信息
- 陈科屹, 张会儒, 张博, 何友均
- CHEN Keyi, ZHANG Huiru, ZHANG Bo, HE Youjun
- 长白山北坡天然次生林典型建群种的种群结构及动态特征
- Population structure and dynamic characteristics of typical constructive species in natural secondary forest on the northern slope of Changbai Mountain
- 生态学报. 2021, 41(13): 5142-5152
- Acta Ecologica Sinica. 2021, 41(13): 5142-5152
- http://dx.doi.org/10.5846/stxb202101050049
-
文章历史
- 收稿日期: 2021-01-05
- 修订日期: 2021-04-12





2. 中国林业科学研究院资源信息研究所, 北京 100091;
3. 北京林业大学森林资源和环境管理国家林业局重点实验室, 北京 100083
2. Research Institute of Forest Resources Information Techniques, Chinese Academy of Forestry, Beijing 100091, China;
3. State Forestry Administration Key Laboratory of Forest Resources & Environmental Management, Beijing Forestry University, Beijing 100083, China
种群是物种个体存在、发展进化以及自然演替的基本单元, 是生态系统的基本构成。种群结构是种群的基本特征, 反映了种群的配置状态以及未来发展与演替趋势, 体现了种群与生境的交互关系及其在群落中的地位和作用。种群的结构和动态一直是种群生态学研究的核心问题之一[1-3]。开展种群结构及动态特征研究有助于揭示种群的生存现状和发展方向, 对于揭示群落演替规律和指导森林生态系统保护修复工作具有重要意义。
长白山林区是东北林区的重要组成部分, 是全球同纬度地带森林资源最丰富的区域之一, 也是我国北疆重要的生态安全屏障。本区域属典型温带森林生态系统, 地带性顶级群落为阔叶红松林, 在维持区域森林生态系统的平衡及稳定中发挥了重要作用[4]。由于长期以来对天然林资源的过量采伐, 导致林区资源锐减, 生态系统质量逐步下降, 珍贵树种濒危, 原始林早已消耗殆尽[5-6]。自国家实施天然林保护工程以来, 长白山林区的森林资源开始受到有效保护, 植被得以逐渐恢复, 形成了当前以臭冷杉(Abies nephrolepis)、鱼鳞云杉(Picea jezoensis var. microsperma)、蒙古栎(Quercus mongolica)、红松(Pinus koraiensis)等为建群种的天然次生林群落, 已成为当前阶段长白山林区重要的组成部分, 在森林生态系统恢复过程中发挥着极为重要的作用。目前, 围绕长白山林区的研究主要集中在森林生态系统服务功能和价值[7]、主要林木材积和生物量模型[8]、群落生产力及种间关系[9-11]、群落环境响应与演替机理[12-13]等方面。但在主要种群的结构和动态研究方面, 目前仅见为数不多的报道[14], 对长白山林区典型群落的建群种种群结构与数量动态的研究还很不充分, 主要种群的生存现状以及天然群落的恢复程度等关键问题亟待给予量化分析和科学评价, 这些研究不足制约着各界对长白山林区典型群落生命过程的认识和天然林保护修复策略的科学制定。
本研究以长白山林区2种典型天然次生林的建群种(鱼鳞云杉、臭冷杉、红松、蒙古栎)为研究对象, 采用空间代替时间的方法, 编制静态生命表, 绘制存活曲线, 分析长白山林区典型天然次生林建群种的种群结构特征;应用4个生存分析函数、种群动态数量化分析以及时间序列预测, 分析其动态变化趋势, 旨在阐明该地区关键种群的结构特征和动态变化规律, 以期为长白山林区的生态系统保护修复和森林可持续经营提供基础资料和科学依据。
1 研究区概况研究区位于长白山北坡吉林省汪清林业局境内, 地形属山地丘陵地带, 气候为温带大陆性季风气候, 年平均气温3.9℃, 年平均降水量547 mm。研究区的森林植被属长白山植物区系, 以针阔叶混交林为主, 目前的常见林分是由原始地带性顶级群落云冷杉林或阔叶红松林遭受人为强烈干扰破坏后形成的天然次生林。主要树种有臭冷杉、鱼鳞云杉、蒙古栎、红松、白桦(Betul aplatyphylla)、山杨(Populus davidiana)、落叶松(Larix gmelinii)、色木槭(Acer mono)、春榆(Ulmus japonica)、紫椴(Tilia amurensis)、水曲柳(Fraxinus mandshurica)、黑桦(Betula dahurica)、樟子松(Pinus sylvestris var. mongolica)、胡桃楸(Junglus mandshurica)、黄檗(Phellodendron amurense)等。
2 研究方法 2.1 数据获取在充分踏查的基础上, 于研究区内选择典型云冷杉天然次生林和蒙古栎-红松天然次生林, 分别设置12块100 m×100 m的样地(云冷杉林表示为YLK-1—YLK-12;蒙古栎-红松林为ZH-1—ZH-12), 样地面积共计24 hm2(表 1)。采用相邻格子法以10 m×10 m为基本单位, 调查记录样地内胸径(DBH)≥1 cm的全部乔木个体的特征值, 包括胸径、树高、冠幅、枝下高等指标。同时, 记录各样地的地理坐标、土壤性质等基本环境指标。
| 样地 Plot |
面积 Area |
坡度 Gradient/(°) |
坡位 Slope position |
海拔 Altitude/m |
郁闭度 Crown density |
| YLK-1 | 1 hm2(100 m×100 m) | 3 | 下 | 742 | 0.59 |
| YLK-2 | 1 hm2(100 m×100 m) | 5 | 中 | 752 | 0.66 |
| YLK-3 | 1 hm2(100 m×100 m) | 15 | 中 | 760 | 0.76 |
| YLK-4 | 1 hm2(100 m×100 m) | 16 | 上 | 773 | 0.66 |
| YLK-5 | 1 hm2(100 m×100 m) | 5 | 中 | 780 | 0.70 |
| YLK-6 | 1 hm2(100 m×100 m) | 15 | 上 | 792 | 0.82 |
| YLK-7 | 1 hm2(100 m×100 m) | 5 | 上 | 771 | 0.77 |
| YLK-8 | 1 hm2(100 m×100 m) | 5 | 下 | 732 | 0.75 |
| YLK-9 | 1 hm2(100 m×100 m) | 5 | 下 | 749 | 0.71 |
| YLK-10 | 1 hm2(100 m×100 m) | 5 | 下 | 759 | 0.76 |
| YLK-11 | 1 hm2(100 m×100 m) | 5 | 上 | 769 | 0.65 |
| YLK-12 | 1 hm2(100 m×100 m) | 3 | 上 | 773 | 0.73 |
| ZH-1 | 1 hm2(100 m×100 m) | 8 | 中 | 705 | 0.66 |
| ZH-2 | 1 hm2(100 m×100 m) | 8 | 中 | 738 | 0.71 |
| ZH-3 | 1 hm2(100 m×100 m) | 7 | 中 | 721 | 0.72 |
| ZH-4 | 1 hm2(100 m×100 m) | 8 | 中 | 741 | 0.83 |
| ZH-5 | 1 hm2(100 m×100 m) | 10 | 上 | 794 | 0.74 |
| ZH-6 | 1 hm2(100 m×100 m) | 8 | 中 | 639 | 0.46 |
| ZH-7 | 1 hm2(100 m×100 m) | 8 | 中 | 637 | 0.85 |
| ZH-8 | 1 hm2(100 m×100 m) | 8 | 中 | 635 | 0.88 |
| ZH-9 | 1 hm2(100 m×100 m) | 7 | 中 | 677 | 0.90 |
| ZH-10 | 1 hm2(100 m×100 m) | 7 | 中 | 685 | 0.77 |
| ZH-11 | 1 hm2(100 m×100 m) | 6 | 中 | 703 | 0.87 |
| ZH-12 | 1 hm2(100 m×100 m) | 10 | 中 | 705 | 0.67 |
| YLK-1—YLK-12: 云冷杉林Picea jezoensis var. microsperma—Abies nephrolepis forest; ZH-1—ZH-12: 蒙古栎-红松林Quercus mongolica—Pinus koraiensis forest | |||||
在生境条件相似的情况下, 物种的径级与龄级对环境的响应情况具有一致性, 当物种年龄难以准确鉴定时, 用径级替代龄级具有一定的科学性[15]。以臭冷杉、鱼鳞云杉、蒙古栎和红松4个建群种的DBH为基准, 当DBH<5 cm划分为1龄级;当DBH≥5 cm按每5 cm为一个龄级, 即5 cm≤DBH<10 cm划分为2龄级, 10 cm≤DBH<15 cm划分为3龄级, 以此类推。据此标准, 臭冷杉共划分为11个龄级, 鱼鳞云杉共划分为11个龄级, 蒙古栎共划分为13个龄级, 红松共划分为8个龄级, 具体情况见表 2。
| 龄级 Age class |
臭冷杉 Abies nephrolepis (stems) |
鱼鳞云杉 Picea jezoensis var. microsperma(stems) |
蒙古栎 Quercus mongolica (stems) |
红松 Pinus koraiensis (stems) |
| 1 | 1458 | 280 | 107 | 2594 |
| 2 | 524 | 156 | 666 | 1378 |
| 3 | 487 | 237 | 719 | 422 |
| 4 | 427 | 214 | 705 | 122 |
| 5 | 339 | 186 | 566 | 50 |
| 6 | 164 | 131 | 333 | 18 |
| 7 | 54 | 48 | 199 | 10 |
| 8 | 29 | 24 | 108 | 1 |
| 9 | 12 | 14 | 63 | — |
| 10 | 2 | 5 | 30 | — |
| 11 | 1 | 2 | 16 | — |
| 12 | — | — | 4 | — |
| 13 | — | — | 3 | — |
| DBH: 胸径Diameter at breast height; DBH<5 cm划分为1龄级;DBH≥5 cm按每5 cm为一个龄级, 即5 cm≤DBH<10 cm划分为2龄级, 10 cm≤DBH<15 cm划分为3龄级, 以此类推 | ||||
由于林木生长周期漫长, 难以实现全周期跟踪调查, 且目前常规的树龄实测方法皆会对林木造成损伤, 因此本研究采用空间代替时间的方法, 即用径级对应龄级, 统计各龄级株数, 编制种群生命表, 进而分析其动态变化。根据4个种群的龄级结构编制静态生命表, 绘制存活曲线。静态生命表主要参数的计算参照江洪的方法[16]。由于静态生命表是一种采用特定时间时代重叠的年龄动态历程替代种群全部生活史的绘制方式, 以及样地调查中系统误差的存在, 均可能造成死亡率为负值的情况, 因此采用匀滑技术(smooth out)对实际种群个体数进行匀滑修正得到ax*, 以避免上述情况[16-18]。
存活曲线采用指数函数Nx=N0e-bx和幂函数Nx=N0x-b对存活数和龄级关系进行拟合[19], 依据决定系数R2和F检验值确定最优模型。如果指数函数拟合效果好, 存活曲线即为Deevey-Ⅱ;幂函数拟合效果好, 存活曲线则为Deevey-Ⅲ。
2.2.3 生存分析生存分析可以辅助种群生命表更好地揭示种群的生存规律。运用生存分析中的4个函数, 分析4个建群种的种群动态变化情况, 其表达式分别如下[20]:
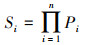
|
(1) |

|
(2) |
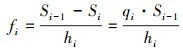
|
(3) |
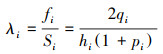
|
(4) |
式中, Pi为存活率, Si为生存率函数;Fi为累计死亡率函数;hi为区间长度, fi死亡密度函数;λi为危险率函数。
2.2.4 种群动态分析采用种群结构分析方法以量化种群或相邻龄级间个体数量的动态关系, 其表达式如下[21]:
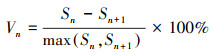
|
(5) |
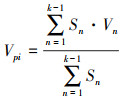
|
(6) |
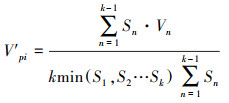
|
(7) |
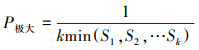
|
(8) |
式中, Vn为种群从n到n+1龄级的种群个体数量变化动态指数, Vpi和V′pi分别为不考虑和考虑外部干扰因素下整个种群结构的数量变化动态指数, Sn和Sn+1分别为第n、第n+1龄级的个体数量, k为种群最大龄级, max和min为取括号中数列极大值和极小值, P极大为种群随机干扰风险极大值。当Vn或Vpi值为正、零、负数时, 态时间序列预测
采用时间序列分析中的一次移动平均法对4个种群的年龄结构进行预测, 其表达式如下[22]:
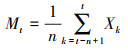
|
(9) |
式中, n为预测时间, t为龄级, Xk为k龄级内的种群个体数量, Mt为n个龄级后t龄级的种群个体数量。
3 结果与分析 3.1 种群生命表和存活曲线由4个建群种的种群静态生命表可知(表 3), 随着龄级的增加, 各个种群的存活个体数量均呈现逐渐减少的趋势, 符合物种的生物学特性。然而, 同一种群在不同龄级的存活量以及不同种群之间存活量的变化却存在较大差异。鱼鳞云杉和蒙古栎种群的死亡率随龄级的增加整体呈上升趋势, 分别在第7龄级和第9龄级出现第一个峰值。2个种群随着龄级的增加, 生命期望值逐渐减小, 这表明二者在幼龄时的生理活动相对旺盛。臭冷杉和红松种群的存活个体数量波动较大, 其中臭冷杉种群在第1龄级就出现了一次死亡高峰, 死亡率高达64.10%, 在第2—4龄级时种群数量相对稳定, 在第6龄级时再次出现死亡高峰, 且持续时间较长, 最高死亡率达到87.50%, 生命期望值在此时降到最低。由于受多轮死亡高峰的影响, 臭冷杉种群的生命期望值也有所波动。红松种群的死亡率一直处在较高的水平, 各龄级死亡率均高于42%, 生命期望值高峰出现在第6龄级。综合来看, 臭冷杉种群的波动幅度最大, 表明其受环境压力的影响更大。
| 种群 Population |
龄级 Age class |
ax* | lx | lnlx | dx | qx | Lx | Tx | ex | Kx |
| 臭冷杉 | 1 | 1458 | 1000 | 6.9078 | 641 | 0.6410 | 680 | 1902 | 1.9020 | 1.0244 |
| Abies nephrolepis | 2 | 524 | 359 | 5.8833 | 25 | 0.0696 | 347 | 1222 | 3.4039 | 0.0722 |
| 3 | 487 | 334 | 5.8111 | 41 | 0.1228 | 314 | 875 | 2.6198 | 0.1310 | |
| 4 | 427 | 293 | 5.6802 | 60 | 0.2048 | 263 | 561 | 1.9147 | 0.2291 | |
| 5 | 339 | 233 | 5.4510 | 121 | 0.5193 | 173 | 298 | 1.2790 | 0.7325 | |
| 6 | 164 | 112 | 4.7185 | 75 | 0.6696 | 75 | 125 | 1.1161 | 1.1076 | |
| 7 | 54 | 37 | 3.6109 | 17 | 0.4595 | 29 | 50 | 1.3514 | 0.6152 | |
| 8 | 29 | 20 | 2.9957 | 12 | 0.6000 | 14 | 21 | 1.0500 | 0.9163 | |
| 9 | 12 | 8 | 2.0794 | 7 | 0.8750 | 5 | 7 | 0.8750 | 2.0794 | |
| 10 | 2 | 1 | 0.0000 | 0 | 0.0000 | 1 | 2 | 2.0000 | 0.0000 | |
| 11 | 1 | 1 | 0.0000 | — | — | 1 | 1 | 1.0000 | — | |
| 鱼鳞云杉 | 1 | 296 | 1000 | 6.9078 | 132 | 0.1320 | 934 | 3885 | 3.8850 | 0.1416 |
| Picea jezoensis var. | 2 | 257 | 868 | 6.7662 | 132 | 0.1521 | 802 | 2951 | 3.3998 | 0.1650 |
| microsperma | 3 | 218 | 736 | 6.6012 | 131 | 0.1780 | 671 | 2149 | 2.9198 | 0.1960 |
| 4 | 179 | 605 | 6.4052 | 132 | 0.2182 | 539 | 1478 | 2.4430 | 0.2461 | |
| 5 | 140 | 473 | 6.1591 | 132 | 0.2791 | 407 | 939 | 1.9852 | 0.3272 | |
| 6 | 101 | 341 | 5.8319 | 132 | 0.3871 | 275 | 532 | 1.5601 | 0.4895 | |
| 7 | 62 | 209 | 5.3423 | 128 | 0.6124 | 145 | 257 | 1.2297 | 0.9479 | |
| 8 | 24 | 81 | 4.3944 | 34 | 0.4198 | 64 | 112 | 1.3827 | 0.5443 | |
| 9 | 14 | 47 | 3.8501 | 30 | 0.6383 | 32 | 48 | 1.0213 | 1.0169 | |
| 10 | 5 | 17 | 2.8332 | 10 | 0.5882 | 12 | 16 | 0.9412 | 0.8873 | |
| 11 | 2 | 7 | 1.9459 | — | — | 4 | 4 | 0.5714 | — | |
| 蒙古栎 | 1 | 685 | 1000 | 6.9078 | 109 | 0.1090 | 946 | 4640 | 4.6400 | 0.1154 |
| Quercus mongolica | 2 | 610 | 891 | 6.7923 | 110 | 0.1235 | 836 | 3694 | 4.1459 | 0.1318 |
| 3 | 535 | 781 | 6.6606 | 109 | 0.1396 | 727 | 2858 | 3.6594 | 0.1503 | |
| 4 | 460 | 672 | 6.5103 | 110 | 0.1637 | 617 | 2131 | 3.1711 | 0.1788 | |
| 5 | 385 | 562 | 6.3315 | 109 | 0.1940 | 508 | 1514 | 2.6940 | 0.2156 | |
| 6 | 310 | 453 | 6.1159 | 110 | 0.2428 | 398 | 1006 | 2.2208 | 0.2782 | |
| 7 | 235 | 343 | 5.8377 | 109 | 0.3178 | 289 | 608 | 1.7726 | 0.3824 | |
| 8 | 160 | 234 | 5.4553 | 110 | 0.4701 | 179 | 319 | 1.3632 | 0.6350 | |
| 9 | 85 | 124 | 4.8203 | 80 | 0.6452 | 84 | 140 | 1.1290 | 1.0361 | |
| 10 | 30 | 44 | 3.7842 | 21 | 0.4773 | 34 | 56 | 1.2727 | 0.6487 | |
| 11 | 16 | 23 | 3.1355 | 17 | 0.7391 | 15 | 22 | 0.9565 | 1.3437 | |
| 12 | 4 | 6 | 1.7918 | 2 | 0.3333 | 5 | 7 | 1.1667 | 0.4055 | |
| 13 | 3 | 4 | 1.3863 | — | — | 2 | 2 | 0.5000 | — | |
| 红松 | 1 | 2594 | 1000 | 6.9078 | 469 | 0.4690 | 766 | 1274 | 1.2740 | 0.6330 |
| Pinus koraiensis | 2 | 1378 | 531 | 6.2748 | 368 | 0.6930 | 347 | 508 | 0.9567 | 1.1810 |
| 3 | 422 | 163 | 5.0938 | 116 | 0.7117 | 105 | 161 | 0.9877 | 1.2436 | |
| 4 | 122 | 47 | 3.8501 | 28 | 0.5957 | 33 | 56 | 1.1915 | 0.9057 | |
| 5 | 50 | 19 | 2.9444 | 12 | 0.6316 | 13 | 23 | 1.2105 | 0.9985 | |
| 6 | 18 | 7 | 1.9459 | 3 | 0.4286 | 6 | 10 | 1.4286 | 0.5596 | |
| 7 | 10 | 4 | 1.3863 | 3 | 0.7500 | 3 | 4 | 1.0000 | 1.3863 | |
| 8 | 1 | 1 | 0.0000 | — | — | 1 | 1 | 1.0000 | — | |
| ax*: 存活个体数修正值Modified survivors number; lx: 标准化存活个体数Standardized survivors number; lnlx: 标准化存活个体数的自然对数The natural logarithm of standardized survivors number; dx: 标准化死亡个体数Standardized number of mortality; qx: 死亡率Mortality; Lx: 标准化平均存活数Standardized mean survivors number; Tx: 标准化存活个体的总数Standardized total individual number; ex: 个体生命期望值Life expectation of individual; Kx: 个体消失率Elimination rate of individual | ||||||||||
由表 4可知, 经模型检验结果显示, 4个建群种种群在两种模型下的拟合结果均达到了极显著水平, 而指数模型的R2值均大于幂函数模型, 且P值更小, 表明4个建群种种群的存活曲线均更趋近于Deevey-Ⅱ型。不同种群间的存活曲线反映了种群的数量变化趋势及结构特征, 结合各个种群的存活曲线(图 1)进一步细化种群所属的亚型, 其中鱼鳞云杉和蒙古栎种群属于Ⅱ1亚型, 即各龄级的存活个体数量相差较大;臭冷杉种群属于属于Ⅱ3亚型, 即幼年期的死亡率较高, 成年后会有所降低, 但是死亡率的波动依然较大;红松种群属于Ⅱ2亚型, 即各龄级的死亡率较为接近, 表明外力干扰一直存在。
| 种群Population | 模型Model | R2 | F | P |
| 臭冷杉 | y=5908.848x-2.758 | 0.726 | 23.907 | 0.001 |
| Abies nephrolepis | y=4899.463e-0.700x | 0.928 | 115.452 | 0.000 |
| 鱼鳞云杉 | y=1042.924x-1.876 | 0.673 | 18.525 | 0.002 |
| Picea jezoensis var. microsperma | y=992.579e-0.489x | 0.907 | 87.680 | 0.000 |
| 蒙古栎 | y=3272.018x-1.974 | 0.629 | 18.620 | 0.001 |
| Quercus mongolica | y=2655.758e-0.460x | 0.881 | 81.728 | 0.000 |
| 红松 | y=8471.224x-3.518 | 0.873 | 41.192 | 0.001 |
| Pinus koraiensis | y=9921.599e-1.072x | 0.982 | 323.767 | 0.000 |

|
| 图 1 4个建群种的存活曲线 Fig. 1 Survival curve of four constructive species lnlx: 标准化存活数的自然对数The natural logarithm of standardized survivors number; DBH<5 cm划分为1龄级;DBH≥5 cm按每5 cm为一个龄级, 即5 cm≤DBH<10 cm划分为2龄级, 10 cm≤DBH<15 cm划分为3龄级, 以此类推 |
由4个种群的生存函数曲线可知(图 2), 生存率和累计死亡率均随龄级的增大分别呈单调递减和单调递增的趋势, 且均表现为幼龄时变幅大, 随后逐渐趋于平稳。但不同种群两类曲线函数的拐点出现的龄级有所差异, 其中红松种群最早, 出现在第4龄级;其次是臭冷杉种群, 出现在第6龄级;鱼鳞云杉和蒙古栎种群相对较晚, 分别出现在第8和第9龄级。鱼鳞云杉和蒙古栎种群的死亡密度曲线走势较为平缓, 死亡密度值分别低于0.0264和0.0220。在早期阶段, 臭冷杉和红松均具有较高的危险率和较大的死亡密度, 表明2个种群幼龄个体的生长过程竞争较为激烈, 随后死亡密度逐渐降低并趋于平缓。4个种群的危险率随龄级增加均呈现波动式上升, 这与死亡率的动态变化一致, 这是由于到龄级后期, 种群个体开始进入生理衰退, 种群数量逐渐减少。相比之下, 臭冷杉种群的危险率起伏较大, 受环境影响的变化更为明显。

|
| 图 2 4个建群种生存率、累计死亡率、死亡密度和危险率函数曲线 Fig. 2 Survival rate, accumulated mortality rate, mortality rate, and hazard rate functional curves of four constructive species |
由表 5可知, 鱼鳞云杉和蒙古栎种群分别在V2和V1、V2出现了负值, 表明2个种群在该时段呈现衰退的结构动态关系;臭冷杉和红松种群在所有指数级的种群动态指数均大于0, 表明2个种群在所有龄级均呈现增长的结构动态关系。根据4个种群的忽略外部干扰和考虑外部干扰的总体数量变化动态指数来看, 各种群均表现为Vpi>V′pi>0, 表明4个种群均属于增长型种群, 增长潜力排序为红松>臭冷杉>鱼鳞云杉>蒙古栎。从随机干扰风险极大值指数来看(P极大), 4个种群受外界干扰的敏感程度均较高, 其中红松种群受干扰的概率最大, 臭冷杉种群次之, 鱼鳞云杉和蒙古栎种群受随机干扰的概率相对较小。
| 指数级 Index |
种群动态指数Dynamic index value/% | |||
| 臭冷杉 Abies nephrolepis |
鱼鳞云杉 Picea jezoensis var. microsperma |
蒙古栎 Quercus mongolica |
红松 Pinus koraiensis |
|
| V1 | 64.06 | 44.29 | -83.93 | 46.88 |
| V2 | 7.06 | -34.18 | -7.37 | 69.38 |
| V3 | 12.32 | 9.70 | 1.95 | 71.09 |
| V4 | 20.61 | 13.08 | 19.72 | 59.02 |
| V5 | 51.62 | 29.57 | 41.17 | 64.00 |
| V6 | 67.07 | 63.36 | 40.24 | 44.44 |
| V7 | 46.30 | 50.00 | 45.73 | 90.00 |
| V8 | 58.62 | 41.67 | 41.67 | — |
| V9 | 83.33 | 64.29 | 52.38 | — |
| V10 | 50.00 | 60.00 | 46.67 | — |
| V11 | — | — | 75.00 | — |
| V12 | — | — | 25.00 | — |
| Vpi | 41.68 | 23.60 | 16.41 | 56.44 |
| V′pi | 3.79 | 1.07 | 0.42 | 7.06 |
| P极大 | 9.09 | 4.55 | 2.56 | 12.50 |
| V1—V12: 相邻龄级数量动态变化指数Quantitative dynamics index of adjacent age class; Vpi: 种群数量动态变化指数Quantitative dynamics index of population; V′pi: 受外界干扰时种群数量动态变化指数Quantitative dynamics index of population by external interference; P极大: 随机干扰风险概率极大值The maximum of probability in random disturbance | ||||
时间序列预测表明(图 3), 在未来2、4、6、8个龄级后, 臭冷杉和红松种群各龄级的个体数量均呈现增长趋势。其中, 在未来2、4个龄级后, 2个种群个体数量增幅随龄级增加的变化趋势也趋于一致, 即随龄级增加个体数量增幅呈波动式上升;但是, 在6、8个龄级后两者有所区别, 臭冷杉种群的个体数量增幅随龄级的增加呈单调递增。对于同一龄级在2、4、6、8个龄级后的变动, 2个种群均表现为时间跨度越大个体数量增幅越多。鱼鳞云杉和蒙古栎种群在幼、中龄级出现了衰退迹象。其中, 鱼鳞云杉种群在未来2个龄级后, 第3龄级的个体数量出现减少;在未来4个龄级后, 第4龄级和第5龄级的个体数量相比于未来2个龄级后的个体数量均有所减少。蒙古栎种群在未来2个龄级后, 第2龄级和第3龄级的个体数量出现减少;在未来4个龄级后, 第4龄级的个体数量出现减少;在未来6个龄级后的第6龄级和未来8个龄级后的第8龄级的个体数量均比其上一个龄级的个体数量有所减少。但是, 随着时间的推移和种群龄级的增长, 依然呈现出增长的势态。

|
| 图 3 4个建群种数量动态时间序列预测 Fig. 3 Time sequence prediction of number dvnamics of four constructive species |
种群结构受到生物和非生物因素的共同影响[23]。研究区4个建群种的种群结构及动态特征是物种生物学特性与生态学过程交互作用的结果。研究表明, 研究区的臭冷杉、鱼鳞云杉、蒙古栎和红松种群的存活曲线均趋近于Deevey-Ⅱ型, 表明当前各种群结构总体处于稳定状态, 但不同种群存在一定差异性。种群个体数量的变化直接受种群更新情况和个体存活情况的影响, 蒙古栎和鱼鳞云杉种群幼龄个体相对不足, 随龄级增大种群危险率也呈上升趋势, 目前依靠数量较为充足的中龄个体维持种群的相对稳定, 但是长期来看2个种群存在偏离稳定状态的风险。在野外调查时发展, 大量蒙古栎存在萌发生长, 更新个体主要源于母树根部或伐桩, 个体数量的扩繁和种群范围的扩大均受到较大限制[24]。且萌生蒙古栎的寿命期远低于实生个体, 因此可能较早出现衰亡。臭冷杉和红松种群个体数量在幼龄时均出现了较大的减少, 表明生境对2个种群的结构和动态有较大影响, 这可能与种群密度大、生存空间较小, 养分竞争强烈等因素有关。尤其是在云冷杉中, 由于部分中、成龄耐阴树种的枝下高较低, 使的幼龄个体除相互竞争以外, 还需要同中、成龄个体的下枝竞争, 导致其死亡率较高。幼龄阶段的高死亡率在其他群落和种群中也普遍存在[25-26]。在中龄阶段, 臭冷杉种群表现出相对稳定, 此阶段臭冷杉在群落中占据了一定的空间和环境资源, 所以表现出了更强的适应能力, 这与滕毅等[27]的研究结果较为一致。臭冷杉种群在高死亡率之后会出现高生命期望值也表明, 存活下来的臭冷杉个体具有较强的生命力。红松的最高生命期望值出现在第6龄级, 表明到此龄级时红松受到的环境筛选和竞争压力有所减小。另外, 由于研究区红松种群缺乏大龄级个体, 这可能影响着红松种群的结构特征。
4个生存分析函数结果显示, 臭冷杉和红松种群在幼龄阶段个体数量大幅度减少, 中期较为平稳, 后期出现衰退。鱼鳞云杉和蒙古栎种群在前期、中期均保持相对平稳, 后期出现衰退。种群数量动态分析表明, 从整体上看各种群均属于增长型, 但对外界的干扰响应较为敏感, 如果外界环境产生波动, 就会影响种群的正常生长。4个种群中红松种群最为敏感, 徐玮泽等[28]经过长期动态监测发现, 长白山原始阔叶红松林中红松、紫椴等优势树种的生长受到了阻碍。种群数量动态时间序列预测分析表明, 臭冷杉和红松种群在未来2、4、6、8个龄级后种群个体数量均呈现不同幅度的增加, 且增长势态较为稳定。鱼鳞云杉和蒙古栎种群在幼、中龄级出了衰退迹象, 但在后期也呈现增长势态。这是由于种群的发展须以数量丰富的幼龄个体为前提, 幼龄个体数量占比越大则增长趋势越强劲[29-30]。由于鱼鳞云杉和蒙古栎种群的幼龄个体相对不足, 种群更新将会受到明显影响。尤其是蒙古栎种群, 幼龄个体数量严重不足, 影响持续的时间更长。目前, 鱼鳞云杉和蒙古栎群种群在整体上仍然呈现增长趋势是因为有相对充足的中龄个体数量。
针对4个建群种种群的生存状况, 有必要进一步加强森林经营管理, 科学开展种群保护和恢复, 真正做到分类施策。当前, 研究区较为适宜臭冷杉和红松种群的生存, 种群总体处于良好的进展状态, 在严格保护的情况下2个种群能够实现自然更新, 应该适度开展上层疏伐, 为幼龄个体顺利进入中、上林层营造空间。针对鱼鳞云杉和蒙古栎种群天然更新不足的情况, 有必要基于近自然经营的思想, 科学开展森林抚育, 及时伐除“霸王树”和病残木, 适度营造小面积林窗, 优化林内生存环境, 促进种群天然更新以及下层林木顺利进入林冠层;对于缺乏实生种的蒙古栎, 有必要加强母树保护与种源保存, 开展种子采集, 在林窗地带开展人工补植补造, 增加实生蒙古栎种苗的数量。此外, 长白山林区是实施天然林保护工程的重点区域, 自天保工程实施以来森林植被开始受到较好保护, 2017年研究区又被划入东北虎豹国家公园, 应该积极把握东北虎豹国家公园建设契机, 不断健全完善国家公园管理制度, 严格保护措施, 强化监督管控, 扎实开展综合执法行动, 加快保护管理能力建设, 切实保护好天然林资源。同时, 还应该充分利用长白山北坡天然林经营国家长期科研基地平台优势, 加强天然林保护修复的理论基础研究和应用基础研究, 重点破解关键种高效繁育、天然林适应性经营、退化天然林生态功能恢复、天然林资源生态综合监测等关键问题, 加快天然林保护修复科技成果的转移转化, 强化科技支撑能力。
| [1] |
Gurevitch J, Scheiner S M, Fox G A. The Ecology of Plants. Sunderland: Sinauer Associates, 2002.
|
| [2] |
董鸣. 生态学透视——种群生态学. 北京: 科学出版社, 2016.
|
| [3] |
Li W, Zhang G F. Population structure and spatial pattern of the endemic and endangered subtropical tree Parrotia subaequalis (Hamamelidaceae). Flora- Morphology, Distribution, Functional Ecology of Plants, 2015, 212: 10-18. DOI:10.1016/j.flora.2015.02.002 |
| [4] |
于大炮, 周旺明, 周莉, 代力民. 长白山区阔叶红松林经营历史与研究历程. 应用生态学报, 2019, 30(5): 1426-1434. |
| [5] |
杨清培, 郝占庆. 长白山林区森林资源变化及其社会效应. 东北林业大学学报, 2006, 34(6): 92-96. DOI:10.3969/j.issn.1000-5382.2006.06.035 |
| [6] |
陈科屹. 云冷杉过伐林经营诊断及目标树抚育效果研究[D]. 北京: 中国林业科学研究院, 2018.
|
| [7] |
李国伟, 赵伟, 魏亚伟, 方向民, 高波, 代力民. 天然林资源保护工程对长白山林区森林生态系统服务功能的影响. 生态学报, 2015, 35(4): 984-992. |
| [8] |
范春楠, 庞圣江, 郑金萍, 李兵, 郭忠玲. 长白山林区14种幼树生物量估测模型. 北京林业大学学报, 2013, 35(2): 1-9. |
| [9] |
李旭华, 于大炮, 代力民, 孙建新. 长白山阔叶红松林生产力随林分发育的变化. 应用生态学报, 2020, 31(3): 706-716. |
| [10] |
殷正, 范秀华. 长白山不同演替阶段温带森林林下草本植物对乔木幼苗的影响. 生态学报, 2020, 40(7): 2194-2204. |
| [11] |
李建, 李晓宇, 曹静, 谭凌照, 赵秀海. 长白山次生针阔混交林群落结构特征及群落动态. 生态学报, 2020, 40(4): 1195-1206. |
| [12] |
樊莹, 乔雪涛, 赵秀海. 长白山自然保护区蒙古栎幼树生理生长特性随海拔梯度的变化. 北京林业大学学报, 2019, 41(11): 1-10. |
| [13] |
洪梓明, 邢亚娟, 闫国永, 张军辉, 王庆贵. 长白山白桦山杨次生林细根形态特征和解剖结构对氮沉降的响应. 生态学报, 2020, 40(2): 608-620. |
| [14] |
李艳丽, 杨华, 亢新刚, 邱实. 长白山云冷杉种群结构和动态分析. 北京林业大学学报, 2014, 36(3): 18-25. |
| [15] |
胡刚, 黎洁, 覃盈盈, 胡宝清, 刘熊, 张忠华. 广西北仑河口红树植物种群结构与动态特征. 生态学报, 2018, 38(9): 3022-3034. |
| [16] |
江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992.
|
| [17] |
吴承祯, 洪伟, 谢金寿, 吴继林. 珍稀濒危植物长苞铁杉种群生命表分析. 应用生态学报, 2000, 11(3): 333-336. DOI:10.3321/j.issn:1001-9332.2000.03.003 |
| [18] |
苏建荣, 张志钧, 邓疆, 陈智勇. 云南红豆杉种群结构与生命表分析. 林业科学研究, 2005, 18(6): 651-656. DOI:10.3321/j.issn:1001-1498.2005.06.002 |
| [19] |
Hett J M, Loucks O I. Age structure models of balsam fir and eastern hemlock. Journal of Ecology, 1976, 64(3): 1029-1044. DOI:10.2307/2258822 |
| [20] |
杨凤翔, 王顺庆, 徐海根, 李邦庆. 生存分析理论及其在研究生命表中的应用. 生态学报, 1991, 11(2): 153-158. DOI:10.3321/j.issn:1001-9332.1991.02.013 |
| [21] |
陈晓德. 植物种群与群落结构动态量化分析方法研究. 生态学报, 1998, 18(2): 214-217. DOI:10.3321/j.issn:1000-0933.1998.02.017 |
| [22] |
肖宜安, 何平, 李晓红, 邓洪平. 濒危植物长柄双花木自然种群数量动态. 植物生态学报, 2004, 28(2): 252-257. DOI:10.3321/j.issn:1005-264X.2004.02.017 |
| [23] |
Velázquez E, Kazmierczak M, Wiegand T. Spatial patterns of sapling mortality in a moist tropical forest: consistency with total density-dependent effects. Oikos, 2016, 125(6): 872-882. DOI:10.1111/oik.02520 |
| [24] |
陈科屹, 张会儒, 雷相东. 天然次生林蒙古栎种群空间格局. 生态学报, 2018, 38(10): 3462-3470. |
| [25] |
陈国鹏, 鲜骏仁, 曹秀文, 刘锦乾, 杨永红, 雷炜. 林窗对岷江冷杉幼苗生存过程的影响. 生态学报, 2016, 36(20): 6475-6486. |
| [26] |
赵阳, 齐瑞, 焦健, 陈学龙, 曹家豪, 冯宜明, 杨萌萌. 尕海-则岔地区紫果云杉种群结构与动态特征. 生态学报, 2018, 38(20): 7447-7457. |
| [27] |
滕毅, 张青, 亢新刚, 杨英军, 王全军. 长白山臭冷杉种群结构与动态. 中南林业科技大学学报, 2017, 37(3): 49-56. |
| [28] |
徐玮泽, 刘琪璟, 孟盛旺, 周光. 长白山阔叶红松林树木种群动态的长期监测. 应用生态学报, 2018, 29(10): 3159-3166. |
| [29] |
赵阳, 曹秀文, 李波, 齐瑞, 曹家豪, 陈学龙, 杨萌萌, 陈林生. 甘肃南部林区4种天然林种群结构特征. 林业科学, 2020, 56(9): 21-29. |
| [30] |
陈科屹, 张会儒, 雷相东, 娄明华, 卢军. 云冷杉过伐林垂直结构特征分析. 林业科学研究, 2017, 30(3): 450-459. |