 2021, Vol. 41
2021, Vol. 41文章信息
- 赵巧华, 徐嘉, 刘玲, 王健键
- ZHAO Qiaohua, XU Jia, LIU Ling, WANG Jianjian
- 垂向湍流、温、光对浅水湖泊下沉藻类垂向分布的影响
- Coupled effects of vertical turbulence, temperature and light on vertical distribution of sinking algae
- 生态学报. 2021, 41(13): 5465-5475
- Acta Ecologica Sinica. 2021, 41(13): 5465-5475
- http://dx.doi.org/10.5846/stxb202002110230
-
文章历史
- 收稿日期: 2020-02-11
- 修订日期: 2021-04-01

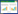




藻类的垂向分布是影响其生长及水生态系统演变的关键要素, 其中SCMax是其分布的主要特征之一[1]。SCMax的形成和维持是物理与生物过程耦合的结果[2], 其深度、强度等特征的变化与水柱的营养状态、垂向湍流的强度、初级生产力及水华暴发等密切相关[3-5]:水柱中, SCMax对应处的初级生产力占比较大, 通常达到30%—70%[6], 可见, 藻类的垂向分布是表征水生态系统演变的重要参数。但由于野外观测的不足(主要包括观测时间频率较低、采样深度间隔难以确定)、遥感又无法获取藻类廓线信息等原因使得有关藻类垂向分布的研究相对滞后。
近年来, 诸多大型浅水湖泊富营养化严重、水华频发, 其中藻类生长是对营养盐、光合有效辐射、温度、动力作用等过程的综合反应[7-9]。气候变化降低了水华暴发所需要的营养盐浓度的阈值, 凸显了物理过程(如气候变化、温、光等)对藻类生长和分布的影响[10-13]。物理过程中湍流主要通过改变藻类的垂向位置、水中光能分布两个方面影响藻类对资源的获取, 进而影响其生长[14-15]。可见, 物理过程(温、光及湍流)不仅能影响藻类生长, 而且还能影响藻类的垂向分布[6, 16], 因而湍流、温、光对藻类生长、分布的影响受到诸多学者的关注:Huisman[17-19]在忽略了温度效应的基础上分析了垂向湍流、光对藻类生长的耦合影响机制。由于温度不仅影响藻类的生长率[20], 同时也会影响藻类生长的最适宜光强, 使得其研究忽略了温度对藻类生长和垂向分布的影响[21]。与海洋相比, 浅水湖泊中营养盐的垂向分布相对均匀, 因而其中SCMax的形成和维持有别于海洋。另一方面, 由于浅水湖泊的几何深度浅, 采样船只扰动等原因导致叶绿素垂向分布的数据匮乏, 限制了人们对大型浅水湖泊中SCMmax生态效应的认识。迄今为止, 关于物理过程(湍流、温、光)影响藻类垂向分布的研究多处于割裂状态或停留在定性阶段[22-23], 从而限制了物理过程在水生态环境修复中的应用、降低了水生态演变的模拟精度。
近三十年来, 长江中下游湖泊水华频发问题久治不愈[24];太湖流域及湖区在经历了长达十年以上的高强度综合治理后仍在2017年出现了前所未有的大面积水华(1403 km2)[25], 说明此类湖泊(如太湖)营养盐浓度偏高、春季水温偏低导致的藻类生长对营养盐需求偏弱, 进而凸显了物理过程(湍流、温、光)的生态作用。另一方面, 春季的优势种群多为硅藻或绿藻[26], 属于下沉藻, 因而下沉藻(硅藻、绿藻)的生长、分布特征直接关系到每年蓝藻水华首次暴发的时间;浅水湖泊中最大初级生产力常出现在次表层, 其强度和深度的变化显著[27]。可见, 湍流、温、光是影响春季浅水湖泊中下沉藻生长、分布的关键物理过程;研究下沉藻的垂向分布特征及其形成机制是提高估算初级生产力精度和探究蓝藻水华暴发机制的关键。因而, 本文以大型浅水湖泊(太湖)的物理场为背景, 采用藻类的生长—扩散模型, 探究春季浅水湖泊中垂向湍流、温及光对下沉藻的垂向分布的影响机制, 为厘清藻类群落演替机制奠定基础。
1 数据与方法 1.1 藻类生长—扩散模型春季的低温和偏高的营养盐浓度使得浅水湖泊中营养盐对春季藻类生长的影响基本可以忽略。本文以春季太湖的物理场为背景场, 假定水温(T)、入射光强(Iin)及水体垂向湍流(D)恒定, 采用生长-扩散模型描述藻细胞密度的时空演变。其一维的控制方程如下:
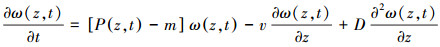
|
(1) |
式中, ω(z, t)在深度为z和时间为t时刻的藻细胞密度;P(z, t)为藻类的生长率;藻类死亡率为m(0.01h-1)[28];藻类在静止水体中的垂向运动速度为v(0.04mh-1)[28]。
水体光能的分布主要受入射光强和水体消光作用两个因素的影响。漫射消光系数(K(z, t))表征水体的消光作用, 是背景漫射消光系数(Kbg)和藻类漫射消光系数之和。
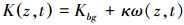
|
(2) |
式中, κ(15×10-12m2cell-1)[29]为单个藻细胞的漫射衰减系数。(3)式描述了水体中光能的垂向廓线随时间的变化。
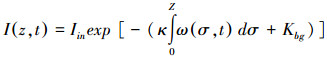
|
(3) |
式中, σ为积分变量, Z为最大水深。光合有效辐射是藻类生长的能量来源, 过强的光能抑制藻体内某些酶的活性, 进而降低藻类光合作用速率(简称:光抑制), 而过弱的光能满足不了藻类生长所需要的的能量而限制藻类生长速率(简称:光限制), 因而存在是最适宜藻类生长的最优光强Iopt(420 μmol m-2s-1)[20]。温度是通过影响藻体中酶的活性而影响藻类的内禀生长率μmax(0.04h-1)[29]。其影响的定量关系如下:
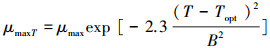
|
(4) |
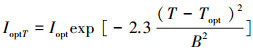
|
(5) |
式中, Topt(16.5℃)[30]为藻类生长的最适宜温度;当水温大于最适宜温度时, B=Tmax-Topt; 当水温小于最适宜温度时, B=Tmin-Topt。注:Tmax、Tmin分别为最高水温和最低水温。μmaxT、IoptT是受水温影响的藻类生长率和最优光强。综上所述, 藻类生长率可表示如下:
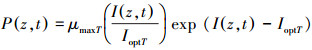
|
(6) |
太湖系典型的大型浅水湖泊, 春季(3月—5月)T通常介于5—30℃;Kbg随时空变化剧烈[31-32], 其值介于0.7—12 m-1;Iin介于0—1900 μmol m-2 s-1, 其中小于9500 μmol m-2 s-1的概率占比85%左右。本文依据太湖的物理场特征, 设置水深(2 m)、Iin(950 μmol m-2 s-1)、Kbg(0.1—7.1m-1, 步长1.5m-1)、T(8—36℃, 步长4℃)、D(0.005—100m2/h, 步长为100倍), 并适度放宽其变化范围。模拟时长为720 h, 即30 d。
1.3 模糊聚类及藻类廓线谱型拟合为了弱化藻类数量对其垂向分布的影响, 实现不同垂向分布的比较, 本文先对藻细胞数密度的廓线(从第25小时—720小时的逐时数据)进行归一化处理, 证实了归一化藻细胞廓线基本不随时间变化。随后利用(4)式对藻类归一化廓线谱型(简称:垂向谱型)进行拟合。
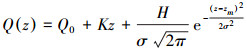
|
(7) |
式中, Q(z)是深度z处的归一化藻细胞密度;Q0是归一化藻细胞密度的背景值;K是归一化藻细胞密度随深度变化的斜率;H是归一化藻细胞密度的垂直积分值;σ为标准化方差;zm为最大归一化藻细胞对应的深度。
2 结果与分析 2.1 藻类生长率廓线谱型特征图 1呈现了归一化藻类生长率廓线形状随温度、漫射消光系数的变化。

|
| 图 1 不同温度和漫射消光系数的归一化藻类生长率 Fig. 1 Normalized algae growth rates at different temperatures and diffuse attenuation coefficients |
尽管水温是影响藻类生长率的关键因素之一, 但均匀的水温通常不会造成生长率的垂向差异, 因而先分析消光作用对藻类生长率廓线的影响。
弱消光作用(K=0.1 m-1)的水体中光能充足, 进而抑制藻类生长。随深度的增大, 光能稍有减弱, 从而导致藻类生长率随深度逐渐增大, 直至底层(图 1);随消光作用的加强, 光能衰减加剧, 表层光能依然抑制藻类的生长, 但随随深度的增大, 光能逐渐衰减到难以满足藻类的生长需求(即光限制)。光抑制向光限制的转变导致次表层生长率峰值的形成(图 1)。该峰出现的深度随水体消光作用的增大而逐渐上移(图 1)。底层, 藻类生长率随深度基本保持不变, 说明藻类的生长可能已经停止, 且水体消光能力越强, 藻类生长停止的深度越浅。
在水温升高至藻类最适宜温度的过程中, 低温对藻类生长的限制逐渐减弱, 生长率峰值出现的深度逐渐向上迁移;在温度大于藻类最适宜温度并继续升高时, 高温对藻类生长的限制逐渐增强, 峰值出现的深度逐步下移。
2.2 垂向湍流、温和光对藻类垂向分布的影响图 2—图 5分别描述了不同垂向湍流(0.010 m2/h、1.000 m2/h、10.00 m2/h、100.0 m2/h)作用下藻类的垂向谱型对消光作用和温度的响应;其中各子图描述了在确定垂向湍流和温度的条件下垂向谱型随消光作用的变化。

|
| 图 2 垂向湍流扩散为0.100 m2/h的归一化藻类廓线 Fig. 2 Normalized algae profiles with vertical turbulent Diffusion of 0.100 m2/h |

|
| 图 3 垂向湍流扩散为1.000 m2/h的归一化藻类廓线 Fig. 3 Normalized algae profiles with vertical turbulent Diffusion of 1.000 m2/h |

|
| 图 4 垂向湍流扩散为10.00 m2/h的归一化藻类廓线 Fig. 4 Normalized algae profiles with vertical turbulent Diffusion of 10.00 m2/h |

|
| 图 5 垂向湍流扩散为100.0 m2/h的归一化藻类廓线 Fig. 5 Normalized algae profiles with vertical turbulent Diffusion of 100.00 m2/h |
在各垂向湍流和消光作用下的垂向谱型分布存在如下共同点(图 2—图 5):其一, 水温介于8—16℃区间, 各垂向湍流和消光作用下的垂向谱型均是先递减再递增(SCMin)。8℃时SCMin的深度最低, 其值为1.7 m左右, 且SCMin的深度差异也最弱;其二, 随水温的升高, SCMin的深度上移, 该现象在低消光作用条件下尤为突出(如消光系数为0.1 m-1, 其深度略大于1 m)。其三, 除消光系数为0.1 m-1外, 不同消光作用下SCMin的差异(强度和深度两方面)随温度增加而增大、随垂向湍流增强而减弱。另, 当消光系数为0.1 m-1, 藻类生长因光能过强而出现光抑制现象, 温度的升高提升藻类的最优光强进而弱化光抑制, 强化不同温度条件下的垂向谱型的差异。
在8—16℃之间, 所有垂向谱型均存在SCMin, 可见温度是限制下沉藻生长的主要因素(图 2—图 5)。温度小于藻类的最适宜温度, 限制藻类的生长, 导致浅层中藻类生长小于下沉作用的减少, 即藻细胞数随深度减少;随深度的递增, 下沉的藻类渐渐聚集在下层, 且大于呼吸作用的损失, 导致藻类数量随深度递增, 即使在最强的消光作用下, 其随深度递增的规律依然维持, 证实了温度是决定下沉藻SCMin形成的关键因素。
可见, 低温(即水温小于藻类最适宜温度)对藻类生长的限制作用促进了下沉藻类SCMin的形成;温度的升高(低温向最适宜温度逼近)将增大不同消光条件下SCMin的差异, 垂向湍流的作用则相反。
水温介于20—28℃, 当消光作用弱(消光系数介于0.1—3.1 m-1)时, 水柱中光能相对充足, 随深度递增, 光抑制现象减弱, 生长作用增强。与下沉作用相比, 藻类生长作用占主导, 导致藻类数量随深度递增:垂向湍流越弱, 该现象越明显(图 2—图 5);当消光作用逐渐强时, 在浅层, 随深度递增, 光抑制作用减弱, 藻类随深度递增, 当深度继续增加, 消光作用导致藻类生长受到光能限制, 使得藻细胞数随深度递减、SCMax的形成。随着水体消光作用的增大, 藻类生长受到光限制的深度变浅, 因而SCMax对应的深度(DSCMax: Depth of Subsurface Chlorophyll Maxinmum)逐渐向浅表层迁移(图 2—图 5);另一方面, 在弱垂向湍流的水体中, 下沉作用的积累造成近底层藻类数量随深度递增, 即形成垂向谱型的第二拐点(图 2—图 5中的20—28℃的子图)。
随着垂向湍流的裹挟作用的加强(图 2—图 5), 藻类的光抑制和限制作用逐渐减弱, SCMax也逐渐减弱;湍流的裹挟作用减少浅层的藻类获取光能, 弱化表层的藻细胞数随深度增大的强度, 进而可能导致SCMax的消失, 致使垂向谱型中仅存谷值拐点。随消光作用的增强, 该现象越明显(见图 2—图 5中消光系数为6.1 m-1和7.6 m-1的藻类垂向谱型的变化)。
水温介于32—36℃, 温度大于藻类最适宜温度, 限制藻类的生长速率和降低藻类的最优光强, 加剧光对藻类生长的抑制和限制, 促进藻类生长的光抑制向限制的转变, 形成或加剧SCMax, 且温度越高, 该趋势越明显。
垂向湍流作用对藻类垂向谱型的影响:随着垂向湍流作用增强, 裹挟作用促进藻类在垂向分布均匀, 减弱藻类在深层的积聚(1.6 m-1和3.1 m-1), 弱化垂向谱型的拐点变化(图 2—图 5)。强消光作用(4.6—7.6 m-1)导致藻类获取的光量子数减少, 弱化藻类数量随深度增强的趋势, 逐渐形成藻类数量随深度递减的趋势。且消光作用越强, 浅层藻类廓线减弱的趋势和程度越明显。
总之, 低温是下沉藻类深层谷区形成的关键;由光抑制下沉藻类的生长转变为限制其生长是SCMax形成的关键;SCMax的强度及其对应的深度分别受控于垂向湍流强度和消光作用。
2.3 藻类垂向谱型的特征藻类垂向谱型受水体的温度、消光作用及垂向湍流的综合影响。基于垂向谱型的模糊聚类结果(5类), 分析藻类垂向谱型特征及其共性, 以便评估湍流、温、光对它的定量影响。
五类垂向谱型中存在一个共同特征:底层, 所有藻类造谱型均随深度递增, 但递增程度的差异显著(图 6)。

|
| 图 6 归一化藻类廓线的模糊聚类结果 Fig. 6 Fuzzy clustering results of normalized algae profiles 图例中的温度、消光作用和垂向湍流属于第三类垂向谱型 |
第4、五类的水温和消光作用相同, 分别为36℃和3.6 m-1, 不同之处仅为垂向湍流:分别为0.005—0.010 m2/h和0.05—100m2/h。与藻类生长的最适宜温度相比, 水温较高, 它通过降低藻类的最优光强、酶的活性等途径降低藻类生长速率, 加剧浅层光抑制。
第四类垂向湍流作用最弱(分别为0.005 m2/h和0.010 m2/h), 因而藻类的下沉作用占主导:光抑制藻生长的深度较第五类明显;垂向谱型为∑形(图 6)。表层, 因温度和光对藻类生长的抑制较强, 偏弱的垂向湍流导致的藻类损失大于藻类的生长, 使得表层藻类随深度递减;随深度增加, 光抑制得到进一步缓解, 使得藻类生长大于下沉等的损失, 形成藻类随深度递增;当随深度继续增加, 导致光能不足, 藻类再次随深度递减;最后因下沉作用导致底层藻类随深度递增, 且弱湍流作用造成该递增趋势尤为突出。
第五类的垂向湍流强于第四类, 因而第五类中垂向谱型的趋势(递减或递增)及峰值强度弱于第四类;第四、五类中的垂向谱型均存在SCMax, 其对应的深度在1 m左右。
第三类:水温介于20—32℃, 漫射衰减系数为1.6 m-1和3.1 m-1, 垂向湍流作用为0.005 m2/h和0.010 m2/h。水温和光能对藻类生长的抑制偏弱, 藻类生长率偏强(图 1), 导致藻类随深度递增;随深度增加, 藻类生长受到温度抑制和光能不足的限制, 促进SCMax的形成。
通过比较20℃和1.6 m-1、24℃和1.6 m-1、28℃和1.6 m-1、32℃和1.6 m-1条件下垂向湍流扩散分别是0.005 m2/h和0.010 m2/h的藻类垂向谱型:0.010 m2/h的SCMax相对偏弱, 且该DSCMax也呈弱抬升特点, 说明了垂向湍流降低降低DSCMax, 同时也减弱了SCMax的强度(图 6)。
第二类:温度介于20—36℃、漫射消光系数不大于3.1 m-1。垂向谱型的总体特征为随深度递增:当漫射消光作用为3.1 m-1时, 其拐点表现向右突起, 为强递增、弱递减的状态;但消光作用和湍流混合作用都弱时, 藻类下沉作用向底层聚集形成的拐点, 表现为左突起。上述两种状况在湍流作用加强时, 其随深度的变化趋势逐渐接近线性。
另, 在第二类垂向谱型中, 主要包括温度为32℃、漫射消光系数介于1.6—3.1 m-1。和温度为36℃时、漫射消光系数为1.6 m-1的两类组合。此组合证实了三者对藻类垂向谱型的综合影响(尤其是对藻类SCMax的影响)。
第一类垂向谱型的主体特征是藻类随深度递减的趋势, 它的样本数占73.9%。尽管符合第一类特征的样本较多, 但均具备两种共性(图 6):其一, 因衰减作用弱, 水体光能相对充足, 使得表层藻类数量密度最大, 随深度递减, 最后因下沉作用在底层集聚增大。其二, 光抑制转变为光限制, 形成藻类的SCMax。
3 讨论光能是藻类生长的能量来源, 是影响藻类生长的直接因素。温度通过影响酶的活性, 进而影响藻类的生长率和最优光强[20]。垂向湍流通过影响藻类对光能获取而影响藻类的生长。它与温度一样是影响藻类生长的间接因素。
为进一步定量评估垂向湍流、温、光对垂向谱型的影响, 用逐步回归的方法筛选影响DSCMax的关键因素:该因素为漫射衰减系数和水温, 对应的标准化系数分别是-0.341和0.27(P<0.01)。图 7刻画了DSCMax随水温和漫射衰减系数的变化, 其特征如下:DSCMax为0.5 m左右;当水温小于17℃左右、或漫射衰减系数小于0.3 m-1, 不存在SCMax;当水温大于17℃, 随着消光系数的增大, DSCMax逐渐增大。当消光系数介于2—4 m-1时, DSCMax最大;随消光衰减的持续增大, DSCMax减小;温度对DSCMax的影响弱于漫射消光系数。

|
| 图 7 叶绿素峰值对应深度随水温和漫射消光系数的变化 Fig. 7 Variation of SCML with temperature and diffuse attenuation (色标代表DSCMax) (color bar represented DSCMax) |
本文敏感性试验的背景是太湖的物理场:水体混合作用较强, 即水温垂向均匀;水体消光作用较强烈, 远大于海洋;水体深度浅(2 m)等, 因而浅水湖泊的DSCMax浅, 其值介于15—50 cm, 远小于海洋中DSCMax, 如楚科奇海的DSCMax为30 m、加拿大海盆的DSCMax为56 m [33]。
消光作用对DSCMax的影响同时还存在两个极端:其一, 对于浅水湖泊而言, 弱消光作用(如小于1 m-1)使得水体中光能充足, 难以存在因光能不足而限制藻类的生长的现象, 即难以形成SCMax;海洋中水体消光作用较弱, 导致光限制藻类生长的深度(DSCMax)远大于浑浊的太湖。图 2—图 5中藻类数量随深度基本呈现递增状态、或低温状态时藻类数量呈现谷分布状态证实了上述规律。其二, 当水体漫射消光作用过强时, 光能衰减过快, 因光能过强而导致藻类生长受到抑制的深度变浅、厚度变薄(图 2—图 5)。上述规律从侧面也进一步证实了介于2—4 m-1的消光系数更有利于SCMax的形成和维持。
依据高斯分布的特征, 式(7)的σ既可以表征藻类次表层峰值的强度, 也可表征次表层峰的厚度。同样采用逐步回归的方法评估垂向湍流、温、光对σ的影响:影响σ的关键因素是垂向湍流和温度, 对应的标准化系数分别为-0.263和0.217(P<0.012)。垂向湍流是垂向上各种尺度的涡旋运动的叠加, 其裹挟作用导致藻类数量在垂直方向上尽可能趋于均匀分布, 弱化σ;当水温向藻类生长的最适宜温度逼近时, 它一方面促进藻类生长速率, 另一方面促进藻类生长对光的依赖程度, 强化藻类生长受光抑制或限制的程度, 进而增大σ。同理, 随温度升高(初始温度小于17℃), 不同消光作用下藻类垂向分布的谷区差异程度得以逐渐增强。可见, σ的峰形随垂向湍流强度的增加变得更扁平, 即:垂向湍流的作用是弱化σ强度, 增大其厚度;当水温向藻类生长最适宜温度逼近时, 加剧了藻类的光限制或抑制的程度, 进而凸显σ。
湍流、温、光不仅通过直接或间接的方式改变下沉藻类的垂向分布特征, 而且它们之间还存在复杂的相互作用:如光能影响水温及水温的垂向结构, 进而影响水体的湍流混合强度;湍流促进水体混合、弱化水温等的垂向差异;在浅水湖泊中, 湍流通过沉积物再悬浮过程影响水下光场结构等。该相互作用机制在浅水湖泊中尤为复杂, 有待于后续的研究。
4 结论藻类的垂向分布是描述水生态系统特征及其演变过程的重要参数, 垂向湍流、温、光对藻类垂向分布、SCMax强度和位置的影响规律如下:
1) 当温度小于17℃, 温度对藻类生长的限制作用促进下沉藻类垂向分布中谷区的形成;温度的升高(低温向最适宜温度逼近)增大SCMin间的差异。
2) 当温度大于17℃, 藻类生长的光抑制向光限制的转变是形成SCMax的关键过程, DSCMax介于20—50 cm。
3) 介于2 m-1至4 m-1的漫射衰减作用更有利于形成藻类的SCMax。
4) 垂向湍流的裹挟作用影响藻类对光的获取, 弱化藻类SCMax或SCMin的强度。
| [1] |
Mellard J P, Yoshiyama K, Litchman E, Klausmeier C A. The vertical distribution of phytoplankton in stratified water columns. Journal of Theoretical Biology, 2011, 269(1): 16-30. DOI:10.1016/j.jtbi.2010.09.041 |
| [2] |
Fu M Z, Sun P, Wang Z L, Wei Q S, Qu P, Zhang X L, Li Y. Structure, characteristics and possible formation mechanisms of the subsurface chlorophyll maximum in the Yellow Sea Cold Water Mass. Continental Shelf Research, 2018, 165: 93-105. DOI:10.1016/j.csr.2018.07.007 |
| [3] |
Richardson K, Visser A W, Pedersen F B. Subsurface phytoplankton blooms fuel pelagic production in the North Sea. Journal of Plankton Research, 2000, 22(9): 1663-1671. DOI:10.1093/plankt/22.9.1663 |
| [4] |
Cullen J J. Subsurface chlorophyll maximum layers: enduring enigma or mystery solved?. Annual Review of Marine Science, 2015, 7: 207-239. DOI:10.1146/annurev-marine-010213-135111 |
| [5] |
Hickman A E, Moore C M, Sharples J, Lucas M I, Tilstone G H, Krivtsov V, Holligan P M. Primary production and nitrate uptake within the seasonal thermocline of a stratified shelf sea. Marine Ecology Progress Series, 2012, 463: 39-57. DOI:10.3354/meps09836 |
| [6] |
Hanson C E, Pesant S, Waite A M, Pattiaratchi C B. Assessing the magnitude and significance of deep chlorophyll maxima of the coastal eastern Indian Ocean. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2007, 54(8/10): 884-901. |
| [7] |
Leterme S C, Jendyk J G, Ellis A V, Brown M H, Kildea T. Annual phytoplankton dynamics in the Gulf Saint Vincent, South Australia, in 2011. Oceanologia, 2014, 56(4): 757-778. DOI:10.5697/oc.56-4.757 |
| [8] |
Huisman J, Codd G A, Paerl H W, Ibelings B W, Verspagen J M H, Visser P M. Cyanobacterial blooms. Nature Reviews Microbiology, 2018, 16(8): 471-483. DOI:10.1038/s41579-018-0040-1 |
| [9] |
Lindemann C, Visser A, Mariani P. Dynamics of phytoplankton blooms in turbulent vortex cells. Journal of the Royal Society Interface, 2017, 16(8): 20170453. |
| [10] |
Havens K E, Paerl H W. Climate change at a crossroad for control of harmful algal blooms. Environmental Science & Technology, 2015, 49(21): 12605-12606. |
| [11] |
Huo S L, He Z S, Ma C Z, Zhang H X, Xi B D, Xia X H, Xu Y, Wu F C. Stricter nutrient criteria are required to mitigate the impact of climate change on harmful cyanobacterial blooms. Journal of Hydrology, 2019, 569: 698-704. DOI:10.1016/j.jhydrol.2018.12.030 |
| [12] |
朱广伟, 秦伯强, 张运林, 许海, 朱梦圆, 杨宏伟, 李宽意, 闵屾, 沈睿杰, 钟春妮. 2005-2017年北部太湖水体叶绿素a和营养盐变化及影响因素. 湖泊科学, 2018, 30(2): 279-295. |
| [13] |
Zhao Q H, Wang J, Wang J J, Wang J X L. Seasonal dependency of controlling factors on the phytoplankton production in Taihu Lake, China. Journal of Environmental Sciences, 2019, 76: 278-288. DOI:10.1016/j.jes.2018.05.010 |
| [14] |
Wang C, Feng T, Wang P F, Hou J, Qian J. Understanding the transport feature of bloom-forming Microcystis in a large shallow lake: a new combined hydrodynamic and spatially explicit agent-based modelling approach. Ecological Modelling, 2017, 343: 25-38. DOI:10.1016/j.ecolmodel.2016.10.017 |
| [15] |
Backhaus J O, Hegseth E N, Wehde H, Irigoien X, Hatten K, Logemann K. Convection and primary production in winter. Marine Ecology Progress Series, 2003, 251: 1-14. DOI:10.3354/meps251001 |
| [16] |
Siswanto E, Ishizaka J, Yokouchi K. Estimating chlorophyll-a vertical profiles from satellite data and the implication for primary production in the Kuroshio front of the east china sea. Journal of Oceanography, 2005, 61(3): 575-589. DOI:10.1007/s10872-005-0066-7 |
| [17] |
Huisman J, Weissing F J. Competition for nutrients and light in a mixed water column: a theoretical analysis. The American Naturalist, 1995, 146(4): 536-564. DOI:10.1086/285814 |
| [18] |
Huisman J, van Oostveen P, Weissing F J. Critical depth and critical turbulence: two different mechanisms for the development of phytoplankton blooms. Limnology and Oceanography, 1999, 44(7): 1781-1787. DOI:10.4319/lo.1999.44.7.1781 |
| [19] |
Huisman J, Arrayás M, Ebert U, Sommeijer B. How do sinking phytoplankton species manage to persist?. The American Naturalist, 2001, 159(3): 245-254. |
| [20] |
Bouterfas R, Belkoura M, Dauta A. Light and temperature effects on the growth rate of three freshwater. Hydrobiologia, 2002, 489(1/3): 207-217. DOI:10.1023/A:1023241006464 |
| [21] |
周妍, 赵巧华, 刘鹏. 垂向湍流扩散和光耦合对下沉藻增长的影响——基于内陆混浊湖泊(太湖)分析. 中国环境科学, 2019, 39(2): 792-801. DOI:10.3969/j.issn.1000-6923.2019.02.043 |
| [22] |
秦伯强, 杨桂军, 马健荣, 邓建明, 李未, 吴挺峰, 刘丽贞, 高光, 朱广伟, 张运林. 太湖蓝藻水华"暴发"的动态特征及其机制. 科学通报, 2016, 61(7): 759-770. |
| [23] |
Zhou J, Qin B Q, Han X X, Zhu L. Turbulence increases the risk of microcystin exposure in a eutrophic lake (Lake Taihu) during cyanobacterial bloom periods. Harmful Algae, 2016, 55: 213-220. DOI:10.1016/j.hal.2016.03.016 |
| [24] |
朱广伟, 许海, 朱梦圆, 邹伟, 国超旋, 季鹏飞, 笪文怡, 周永强, 张运林, 秦伯强. 三十年来长江中下游湖泊富营养化状况变迁及其影响因素. 湖泊科学, 2019, 31(6): 1510-1524. |
| [25] |
朱伟, 陈怀民, 王若辰, 冯甘雨, 薛宗璞, 胡思远. 2017年太湖水华面积偏大的原因分析. 湖泊科学, 2019, 31(3): 621-632. |
| [26] |
蔡琳琳, 朱广伟, 朱梦圆, 杨桂军, 赵林林. 太湖梅梁湾湖岸带浮游植物群落演替及其与水华形成的关系. 生态科学, 2012, 31(4): 345-351. DOI:10.3969/j.issn.1008-8873.2012.04.001 |
| [27] |
张运林, 秦伯强, 陈伟民, 陈宇炜, 高光. 太湖梅梁湾春季浮游植物初级生产力. 湖泊科学, 2005a, 17(1): 81-86. |
| [28] |
Huisman J, Sommeijer B. Population dynamics of sinking phytoplankton in light-limited environments: simulation techniques and critical parameters. Journal of Sea Research, 2002, 48(2): 83-96. DOI:10.1016/S1385-1101(02)00137-5 |
| [29] |
Huisman J, Sharples J, Stroom J M, Visser P M, Kardinaal W E A, Verspagen J M H, Sommeijer B. Changes in turbulent mixing shift competition for light between phytoplankton species. Ecology, 2004, 85(11): 2960-2970. DOI:10.1890/03-0763 |
| [30] |
Paerl H W, Otten T G. Harmful cyanobacterial blooms: causes, consequences, and controls. Environmental Microbiology, 2013, 65(4): 995-1010. DOI:10.1007/s00248-012-0159-y |
| [31] |
徐德瑞, 周杰, 吴时强, 戴江玉, 吴修锋. 夏季东太湖光合有效辐射衰减特征及其对沉水植物恢复的指示. 湖泊科学, 2021, 33(1): 111-122. |
| [32] |
张运林, 秦伯强, 陈伟民, 胡维平, 高光, 朱广伟, 罗潋葱. 不同风浪条件下太湖梅梁湾光合有效辐射的衰减. 应用生态学报, 2005b, 16(6): 1133-1137. |
| [33] |
Zachary W, Brown Z B, Lowry K E, Palmer M A, van Dijken G L, Mills M M, Pickart R S, Arrigo K R. Characterizing the subsurface chlorophyll a maximum in the Chukchi sea and Canada Basin. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 2015, 118: 88-104. DOI:10.1016/j.dsr2.2015.02.010 |