 2021, Vol. 41
2021, Vol. 41文章信息
- 陈星, 刘正祥, 段兴德, 邵宗体, 浦恩念, 苏超, 杜春红, 李玉琼, 高子厚
- CHEN Xing, LIU Zhengxiang, DUAN Xingde, SHAO Zongti, PU Ennian, SU Chao, DU Chunhong, LI Yuqiong, GAO Zihou
- 滇西北高原藏区小型兽类体表寄生蚤的生态位特征
- Niche characteristics of parasitic fleas on the body surface of small mamals in Deqin County, the Tibetan area of Northwestern Yunnan Plateau
- 生态学报. 2021, 41(13): 5476-5486
- Acta Ecologica Sinica. 2021, 41(13): 5476-5486
- http://dx.doi.org/10.5846/stxb202001200153
-
文章历史
- 收稿日期: 2020-01-20
- 修订日期: 2021-04-07





2. 云南省地方病防治所, 大理 671000
2. Yunnan Institute for Endemic Diseases Control and Prevention, Dali 671000, China
生态位指物种在生态系统中的位置, 包括生境需求及其功能作用, 既能反映该物种在各生态因子中的位置, 又能反映其在物质循环与能量流动以及信息传递时所扮演的角色, 一般主要运用生态位宽度和生态位重叠指数两大指标来分析特定种群的生态位特征[1-3]。生态位宽度可以解释物种的丰度及其分布的特征与种间差异, 生态位重叠则是度量物种间在生态因子联系上的相似性。
寄生蚤作为蚤传疾病、鼠传疾病的主要传播媒介, 在鼠疫耶尔森菌等病菌的储存及传播过程中扮演重要角色, 且多次从云南西北部高原藏区德钦县鼠疫指示动物犬血清中检测出F1抗体阳性, 提示德钦县可能存在鼠疫[4-5]。目前国内对植物、海洋生物、哺乳动物的生态学研究较多, 小兽寄生蚤的生态学分析还存在很大的空白, 因此明确德钦县寄生蚤的生态学特征既对该地蚤传疾病的防控至关重要, 又是对该领域生态学研究的补充。生态位作为一个宏观概念, 在特定种群中, 受多个生态因子的影响, 故本文从海拔、生境和寄主3个资源轴对德钦县小兽体表寄生蚤的生态位特征进行分析, 了解不同生态因子中蚤种间分化、共存特征, 为蚤传疾病防控提供可靠生态学依据。
1 材料与方法 1.1 研究区域概况德钦县(98°3′56″—99°32′20″E, 27°33′44″—29°15′2″N)位于云南省迪庆藏族自治州, 地处滇西北横断山脉地区的滇川藏三省交界处, 与鼠疫自然疫源地芒康县、巴塘县、德格县、玉龙县相邻, 为三江(澜沧江、怒江、金沙江)并流核心区, 是云南省地势最高的县城。境内最高海拔6740m, 最低1464m, 属高山深谷地貌, 立体气候明显, 年均气温6.6℃, 年均降水量631.7mm[6], 干湿季分明, 为蚤类等媒介生物的繁殖提供了有利条件[7]。2012年6月—2014年5月, 依据海拔、生境等自然环境特征, 在德钦县境内选取42个调查点, 进行群落生态学调查。
1.2 资源划分海拔范围:2000—4500m, 以500m为一个梯度, 将其划分为5个海拔梯度带, 依次以A—E表示;将生境划分为4类:林地、耕地、灌丛、居民区, 以Ⅰ—Ⅳ表示。
1.3 样本采集与处理以鲜炸油条为诱饵, 野外采用铗夜法[8]布放鼠铗, 居民区采用笼日法[9]布放鼠笼。根据《中国动物志》和《中国脊椎动物红色名录》[10-11]依据外部特征对捕获的小兽进行物种鉴定, 梳捡其体表寄生蚤, 保存于75%的乙醇溶液, 依据《云南蚤类志》[12], 在显微镜下进行形态学特征鉴定并记录。
1.4 生态位指标测定 1.4.1 生态位宽度测定采用Shannon-Wiener指数[13]来计算寄生蚤的生态位宽度, 计算公式:

|
其中, Bi=i种的生态位宽度, Nij=i种利用j资源等级的数值, r=生态位的资源等级数。生态位宽度范围:0—1, 0表示没有利用, 1表示对所有等级都同样的利用了。
根据李德志[14]等的研究, 对生态位进行整合, 将多个单一资源轴的生态位宽度值进行PCA(主成分分析)处理后整合成2个或单个新的资源轴:

|
其中, Bin为多维空间生态位宽度指数, Bi为第i个生态因子轴上的生态位宽度值, n为生态因子轴个数。
1.4.2 生态位重叠指数测定采用Pianka. E.R生态位重叠指数[15]公式来计算蚤的生态位重叠指数, 计算公式:
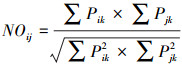
|
其中, NOij= i种和j种之间的生态位重叠指数, Pik=物种i种在k资源等级的数量与物种i在所有资源等级的总数量之比, Pjk=物种j种在k资源等级的数量与物种j在所有资源等级的总数量之比。生态位重叠指数的变化范围:0—1, 0表示完全不重叠, 1表示百分之百重叠, NO>0.6为有意义的重叠, 说明重叠较为严重[16]。
1.4.3 统计分析各种指标计算均在EXCEL、SPSS 21.0中完成。
2 结果 2.1 寄生蚤构成及数量特征本次调查共捕获小兽1371只, 隶属4目7科18属28种, 共梳捡蚤1275头, 隶属于4科9亚科17属33种。按照徐正会[17]将物种构成比大于10%定义为优势种, 1%—10%位常见种, 小于1%为稀有种的定义, 本次调查结果中迪庆额蚤(Frontopsylla(Frontopsylla)diqingensis)、云南栉眼蚤(Ctenophthalmus(Sinoctenophthalmus)yunnanus)为优势种, 分别占所捡蚤匹的25.02%、22.82%;特新蚤德钦亚种(Neopsylla specialis dechingensis)、特新蚤指名亚种(Neopsylla specialis specialis)、相关新蚤指名亚种(Neopsylla affinis affinis)、喜山二刺蚤川滇亚种(Peromyscopsylla himalaica sichuanoyunnana)、结实茸足蚤(Geusibia(Geusibia)torosa)、棕形额蚤(Frontopsylla(Frontopsylla)spadix spadix)、绒鼠怪蚤(Paradoxopsylla custodies)、直缘双蚤德钦亚种(Amphipsylla tuta deqinensis)、无值大锥蚤(Macrostylophora euteles)、二刺大锥蚤(Macrostylophora bispiniforma bispiniforma)、卷带倍蚤(Amphalius spirataenius spirataenius)以及细钩盖蚤(Callopsylla(Callopsylla)sparsilis sparsilis)为常见种;其余均为稀有种。在海拔选择上, 蚤匹主要分布在中高海拔带, 3500—4000m海拔带捡蚤最多。在生境选择上, 林地捡蚤数量、种类最多(表 1)。在寄主选择上, 优势蚤种云南栉眼蚤主要寄生于西南绒鼠(Eothenomys custos), 迪庆额蚤则以姬鼠属中的大耳姬鼠(Apodemus latronum)和中华姬鼠(Apodemus draco)为主要寄主。所有寄主中, 西南绒鼠体外寄生蚤数量最多, 达383头, 其次为大耳姬鼠、藏鼠兔(Ochotona thibetana)和珀氏长吻松鼠(Dremomys pernyi);大耳姬鼠体外寄生蚤种类最多, 达15种, 其次为西南绒鼠、中华姬鼠、白尾松田鼠(Phaiomys leucurus)(表 2)。
| 蚤种编号 Species No. | 海拔/m Altitude gradient | 生境 Habitat | 合计 Total | 构成比 Cr% | 生态位宽度指数 Index of niche breadth | |||||||||
| 2000—2500 | 2500—3000 | 3000—3500 | 3500—4000 | 4500—5000 | 林地 | 耕地 | 灌丛 | 居民区 | Bi1 | Bi2 | ||||
| 1 | 2 | 2 | 2 | 0.16 | 0 | 0 | ||||||||
| 2 | 10 | 10 | 10 | 0.78 | 0 | 0 | ||||||||
| 3 | 5 | 5 | 5 | 0.39 | 0 | 0 | ||||||||
| 4 | 1 | 5 | 18 | 2 | 18 | 6 | 2 | 26 | 2.04 | 0.556 | 0.570 | |||
| 5 | 5 | 5 | 27 | 68 | 9 | 84 | 20 | 10 | 114 | 8.94 | 0.698 | 0.537 | ||
| 6 | 7 | 7 | 7 | 0.55 | 0 | 0 | ||||||||
| 7 | 3 | 14 | 17 | 17 | 1.33 | 0.290 | 0 | |||||||
| 8 | 1 | 5 | 1 | 1 | 2 | 6 | 8 | 0.63 | 0.667 | 0.406 | ||||
| 9 | 1 | 1 | 1 | 0.08 | 0 | 0 | ||||||||
| 10 | 2 | 2 | 2 | 0.16 | 0 | 0 | ||||||||
| 11 | 1 | 2 | 1 | 2 | 2 | 4 | 0.31 | 0.646 | 0.5 | |||||
| 12 | 1 | 1 | 2 | 2 | 0.16 | 0.431 | 0 | |||||||
| 13 | 2 | 2 | 2 | 0.16 | 0 | 0 | ||||||||
| 14 | 9 | 9 | 9 | 0.71 | 0 | 0 | ||||||||
| 15 | 1 | 1 | 1 | 0.08 | 0 | 0 | ||||||||
| 16 | 9 | 28 | 229 | 25 | 251 | 25 | 15 | 291 | 22.82 | 0.455 | 0.354 | |||
| 17 | 13 | 5 | 11 | 8 | 21 | 29 | 2.27 | 0.640 | 0.425 | |||||
| 18 | 1 | 1 | 2 | 2 | 0.16 | 0.431 | 0 | |||||||
| 19 | 1 | 18 | 13 | 26 | 6 | 32 | 2.51 | 0.496 | 0.348 | |||||
| 20 | 2 | 18 | 23 | 6 | 7 | 42 | 49 | 3.84 | 0.690 | 0.296 | ||||
| 21 | 5 | 14 | 38 | 199 | 63 | 241 | 42 | 36 | 319 | 25.02 | 0.665 | 0.523 | ||
| 22 | 2 | 1 | 1 | 2 | 3 | 0.24 | 0.395 | 0.459 | ||||||
| 23 | 4 | 17 | 1 | 2 | 1 | 3 | 21 | 1 | 25 | 1.96 | 0.631 | 0.382 | ||
| 24 | 1 | 3 | 1 | 3 | 2 | 5 | 0.39 | 0.590 | 0.485 | |||||
| 25 | 5 | 1 | 5 | 1 | 6 | 0.47 | 0.280 | 0.325 | ||||||
| 26 | 1 | 1 | 1 | 0.08 | 0 | 0 | ||||||||
| 27 | 1 | 4 | 5 | 5 | 0.39 | 0.311 | 0 | |||||||
| 28 | 21 | 2 | 21 | 2 | 23 | 1.80 | 0.184 | 0.213 | ||||||
| 29 | 3 | 32 | 39 | 1 | 45 | 24 | 6 | 75 | 5.88 | 0.553 | 0.630 | |||
| 30 | 3 | 60 | 7 | 70 | 70 | 5.49 | 0.309 | 0 | ||||||
| 31 | 1 | 70 | 30 | 94 | 1 | 6 | 101 | 7.92 | 0.410 | 0.202 | ||||
| 32 | 13 | 12 | 23 | 2 | 25 | 1.96 | 0.430 | 0.201 | ||||||
| 33 | 4 | 4 | 4 | 0.31 | 0 | 0 | ||||||||
| 合计 Total | 40 | 97 | 175 | 785 | 178 | 949 | 221 | 104 | 1 | 1275 | 100 | 0.715 | 0.529 | |
| 1:人蚤Pulex irritans; 2:鼠兔角头蚤Echidnophaga ochotona; 3:圆凹多毛蚤Hystrichopsylla(Hystroceras)zii; 4:特新蚤指名亚种Neopsylla specialis specialis; 5:特新蚤德钦亚种Neopsylla specialis dechingensis; 6:二毫新蚤指名亚种Neopsylla biseta biseta; 7:相关新蚤指名亚种Neopsylla affinis affinis; 8:斯氏新蚤川滇亚种Neopsylla stevensi sichuanyunnana; 9:后棘新蚤Neopsylla honora honora; 10:棒突继新蚤Genoneopsylla claviprocera; 11:高山狭臀蚤Stenischia montanis montanis; 12:五侧纤蚤Rhadinopsylla (Actenophthalmus)dahurica vicina; 13:纪氏叉蚤Doratopsylla jii; 14:无突栉眼蚤Ctenophthalmus(Sinoctenophthalmus)aprojectus; 15:泸水栉眼蚤Ctenophthalmus(Sinoctenophthalmus)lushuiensis; 16:云南栉眼蚤: Ctenophthalmus(Sinoctenophthalmus)yunnanus; 17:喜山二刺蚤川滇亚种Peromyscopsylla himalaica sichuanoyunnana; 18:云南茸足蚤Geusibia(Geusibia)yunnanensis; R19:结实茸足蚤Geusibia(Geusibia)torosa; 20:棕形额蚤Frontopsylla(Frontopsylla)spadix spadix; 21:迪庆额蚤Frontopsylla(Frontopsylla)diqingensis; 22:毛额蚤Frontopsylla(Frontopsylla)tomentosa; 23:绒鼠怪蚤Paradoxopsylla custodies; 24:介中怪蚤Paradoxopsylla intermedius; 25:金沙江怪蚤Paradoxopsylla jinshajiangensis jinshajiang jinshajiangensis; 26:长突怪蚤Paradoxopsylla longiprojectus; 27:直缘双蚤察里亚种Amphipsylla tuta chaliensis; 28:直缘双蚤德钦亚种Amphipsylla tuta deqinensis; 29:无值大锥蚤Macrostylophora euteles; 30:二刺大锥蚤Macrostylophora bispiniforma bispiniforma; 31:卷带倍蚤Amphalius spirataenius spirataenius; 32:细沟盖蚤Callopsylla(Callopsylla)sparsilis sparsilis; 33:昌都盖蚤Callopsylla(callopsylla)changduensis; Bi1:以海拔资源等级计算的海拔生态位宽度指数, altitude niche breadth calculated based on altitude;Bi2:以生境为资源等级计算的生境生态位宽度指数, habitat niche breadth calculated based on habit | ||||||||||||||
在海拔资源轴上, 总体生态位宽度值为0.715, 蚤类生态位宽度范围为0—0.698, 其中生态位宽度值最大的是特新蚤德钦亚种, 为0.698;其次是棕形额蚤、斯氏新蚤川滇亚种、迪庆额蚤、高山狭臀蚤(Stenischia montanis montanis)、喜山二刺蚤川滇亚种和绒鼠怪蚤, 生态位宽度值分别为0.690、0.667、0.665、0.646、0.640、0.631;而人蚤(Pulex irritans)等11种蚤类在该资源轴上的生态位宽度值为0, 仅分布在单一海拔带, 未利用海拔等级资源(表 1)。
2.2.2 生境生态位宽度(Bi2)在生境资源轴上, 总体生态位宽度值为0.529, 相对海拔资源轴的总体生态位宽度较窄。蚤类生态位宽度范围:0—0.630, 其中无值大锥蚤的生态位最宽, 为0.630;其次是特新蚤指名亚种、特新蚤德钦亚种和迪庆额蚤, 生态位宽度分别为0.570、0.537、0.523;而人蚤等16种蚤的生态位宽度为0, 仅分布在单一生境, 未利用生境等级资源(表 1)。
2.2.3 寄主生态位宽度(Bi3)在寄主资源轴上, 总体生态位宽度为0.749, 是三种资源轴的总体生态位宽度中的最大值。蚤类的寄生生态位宽度值范围为0—0.584, 其中绒鼠怪蚤的生态位较宽, 宽度值为0.584;其次是迪庆额蚤、棕形额蚤、喜山二刺蚤川滇亚种、特新蚤德钦亚种和斯氏新蚤川滇亚种(Neopsylla stevensi sichuanyunnana), 其生态位宽度值分别为:0.567、0.553、0.550、0.537和0.530;而鼠兔角头蚤(Echidnophaga ochotona)和圆凹多毛蚤(Hystrichopsylla(Hystroceras)zii)等9种蚤类的生态位宽度为0, 仅寄生在单一宿主, 未利用小兽等级资源(表 2)。
2.2.4 多维生态位宽度(Bin)利用李德志对多个单维生态位宽度值整合的方法将三个生态因子进行整合, 得到多维生态位宽度值Bin, 为寄生蚤在海拔、生境、寄主3个资源轴上的整体生态位宽度。结果显示多维生态位宽度值范围为0—1.031, 特新蚤德钦亚种和迪庆额蚤生态位宽度值最大, 分别为1.031、1.019, 在所有蚤种中占据相对最优的分布形式;其次为喜山二刺蚤川滇亚种、斯氏新蚤川滇亚种、绒鼠怪蚤、棕形额蚤, 分别为0.945、0943、0.941、0.932, 也显示出较优分布。鼠兔角头蚤、圆凹多毛蚤、后棘新蚤(Neopsylla honora honora)、纪氏叉蚤(Doratopsylla jii)、泸水栉眼蚤(Ctenophthalmus(Sinoctenophthalmus)lushuiensis)、长突怪蚤(Paradoxopsylla longiprojectus)、昌都盖蚤(Callopsylla(callopsylla)changduensis)7种蚤类生态位宽度值为0, 在三种生态资源轴中均为单一分布形式(表 2)。
| 蚤种编号 No. | 寄主 host | 合计 Total | 生态位宽度指数 | |||||||||||||||||||
| R. ni | N.c | A.c | A.l | A.d | N.a | O.t | T.s | M.c | V.f | E.c | P.l | C.r | D.p | R.t | O.k | T.b | R.f | R.no | Bi3 | Bin | ||
| 1 | 1 | 1 | 2 | 0.235 | 0.235 | |||||||||||||||||
| 2 | 10 | 10 | 0 | 0 | ||||||||||||||||||
| 3 | 5 | 5 | 0 | 0 | ||||||||||||||||||
| 4 | 2 | 2 | 5 | 17 | 26 | 0.336 | 0.864 | |||||||||||||||
| 5 | 2 | 3 | 8 | 26 | 21 | 3 | 47 | 4 | 114 | 0.537 | 1.031 | |||||||||||
| 6 | 1 | 5 | 1 | 7 | 0.27 | 0.27 | ||||||||||||||||
| 7 | 17 | 17 | 0 | 0.29 | ||||||||||||||||||
| 8 | 2 | 2 | 1 | 2 | 1 | 8 | 0.53 | 0.943 | ||||||||||||||
| 9 | 1 | 1 | 0 | 0 | ||||||||||||||||||
| 10 | 1 | 1 | 2 | 0.235 | 0.235 | |||||||||||||||||
| 11 | 2 | 2 | 4 | 0.235 | 0.85 | |||||||||||||||||
| 12 | 1 | 1 | 2 | 0.235 | 0.491 | |||||||||||||||||
| 13 | 2 | 2 | 0 | 0 | ||||||||||||||||||
| 14 | 2 | 1 | 6 | 9 | 0.288 | 0.288 | ||||||||||||||||
| 15 | 1 | 1 | 0 | 0 | ||||||||||||||||||
| 16 | 1 | 2 | 1 | 286 | 1 | 291 | 0.037 | 0.578 | ||||||||||||||
| 17 | 3 | 6 | 10 | 1 | 7 | 1 | 1 | 29 | 0.55 | 0.945 | ||||||||||||
| 18 | 2 | 2 | 0 | 0.431 | ||||||||||||||||||
| 19 | 30 | 1 | 1 | 32 | 0.094 | 0.613 | ||||||||||||||||
| 20 | 3 | 18 | 17 | 1 | 2 | 2 | 2 | 1 | 3 | 49 | 0.553 | 0.932 | ||||||||||
| 21 | 11 | 14 | 146 | 70 | 32 | 11 | 1 | 19 | 8 | 1 | 1 | 5 | 319 | 0.567 | 1.019 | |||||||
| 22 | 2 | 1 | 3 | 0.216 | 0.643 | |||||||||||||||||
| 23 | 9 | 4 | 3 | 1 | 1 | 3 | 4 | 25 | 0.584 | 0.941 | ||||||||||||
| 24 | 2 | 1 | 2 | 5 | 0.358 | 0.844 | ||||||||||||||||
| 25 | 1 | 1 | 4 | 6 | 0.295 | 0.52 | ||||||||||||||||
| 26 | 1 | 1 | 0 | 0 | ||||||||||||||||||
| 27 | 1 | 1 | 1 | 2 | 5 | 0.452 | 0.549 | |||||||||||||||
| 28 | 1 | 1 | 21 | 23 | 0.121 | 0.306 | ||||||||||||||||
| 29 | 2 | 1 | 71 | 1 | 75 | 0.09 | 0.843 | |||||||||||||||
| 30 | 2 | 11 | 1 | 56 | 70 | 0.215 | 0.376 | |||||||||||||||
| 31 | 1 | 97 | 1 | 2 | 101 | 0.071 | 0.463 | |||||||||||||||
| 32 | 6 | 1 | 18 | 25 | 0.24 | 0.532 | ||||||||||||||||
| 33 | 4 | 4 | 0 | 0 | ||||||||||||||||||
| 合计 Total | 20 | 47 | 57 | 194 | 111 | 64 | 144 | 11 | 11 | 1 | 383 | 60 | 2 | 129 | 6 | 17 | 2 | 15 | 1 | 1275 | 0.749 | 1.163 |
| Bi3:以小兽为资源等级计算的小兽生态位宽度指数host niche breadth calculated based on small mammals; Bin: 整合Bi1、Bi2、Bi3计算的小兽生态位宽度Bin was calculated by integrating Bi1Bi2 and Bi3;R.ni: 大足鼠Rattus nitidus; N.c: 社鼠Niviventer confucianus; A.c: 齐氏姬鼠Apodemus chevrieri; A.L: 大耳姬鼠Apodemus latronum; A.d: 中华姬鼠Apodemus draco; N.a: 安氏白腹鼠Niviventer andersoni; O.t: 藏鼠兔Ochotona thibetana; T.s: 隐纹花松鼠Tamipps swinhoei; M.c: 克氏田鼠Microtus clarkei; V.f: 滇攀鼠Vernaya fulva; E.c: 西南绒鼠Eothenomys custos; P.l: 白尾松田鼠Phaiomys leucurus; C.r: 中麝鼩Crocidura rapax; D.p: 珀氏长吻松鼠Dremomys pernyi; R.t: 中亚鼠Rattus turkestanicus; O.k: 柯氏鼠兔Ochotona koslowi; T.b: 树鼩Tupaia belangeri; R.f: 黄胸鼠Rattus flavipectus; R.no: 斑胸鼠Rattus norvegicus | ||||||||||||||||||||||
海拔生态位重叠指数中, 0.6<NO<1的种对有188对, 为有意义的重叠, 占所有种对的35.6%, 多为优势种、常见种、稀有种两者之间交叉重叠;NO=0的有142对, 占比26.89%, 表示完全不重叠, 多为稀有种与稀有种之间的种对, 最多的是与人蚤和鼠兔角头蚤的种对;NO=1的有12对, 占比2.27%, 表示完全重叠, 均为稀有种之间的重叠, 最多的是与圆凹多毛蚤、无突栉眼蚤(Ctenophthalmus(Sinoctenophthalmus)aprojectus)的重叠, 其次是与泸水栉眼蚤和棒突继新蚤(Genoneopsylla claviprocera)的重叠。(表 3)
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | |
| 1 | 0 | 0 | 0 | △ | 0 | 0 | △ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | □ | 0 | 0 | △ | △ | 0 | △ | △ | □ | 1 | 0 | 0 | 0 | 0 | △ | 0 | 0 | |
| 2 | 0 | 0 | △ | △ | 0 | 0 | □ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | △ | △ | 0 | △ | □ | △ | □ | □ | □ | 0 | 0 | 0 | 0 | △ | 0 | 0 | 0 | 1 | |
| 3 | 0 | 0 | □ | □ | 1 | □ | △ | 1 | 0 | □ | □ | 0 | 1 | 0 | □ | 0 | □ | □ | △ | □ | 0 | △ | 0 | 0 | 0 | △ | □ | □ | □ | □ | □ | 0 | |
| 4 | △ | △ | □ | □ | □ | □ | △ | □ | △ | □ | □ | △ | □ | △ | □ | △ | □ | □ | △ | □ | △ | △ | △ | △ | 0 | △ | □ | □ | □ | □ | □ | △ | |
| 5 | △ | △ | □ | □ | □ | □ | △ | □ | △ | □ | □ | △ | □ | △ | □ | △ | □ | □ | △ | □ | △ | △ | △ | △ | △ | △ | □ | □ | □ | □ | □ | △ | |
| 6 | 0 | 0 | 1 | □ | □ | □ | △ | 1 | 0 | □ | □ | 0 | 1 | 0 | □ | 0 | □ | □ | △ | □ | 0 | △ | 0 | 0 | 0 | △ | □ | □ | □ | □ | □ | 0 | |
| 7 | 0 | 0 | 1 | □ | □ | 1 | △ | □ | 0 | □ | □ | 0 | □ | 0 | □ | △ | □ | □ | △ | □ | 0 | △ | △ | △ | 0 | △ | □ | □ | □ | □ | □ | 0 | |
| 8 | □ | 0 | △ | □ | △ | △ | △ | △ | 0 | △ | △ | 0 | △ | 0 | △ | △ | △ | △ | □ | △ | □ | □ | □ | △ | △ | △ | △ | △ | △ | △ | △ | □ | |
| 9 | 0 | 0 | 1 | □ | □ | 1 | 1 | △ | 0 | □ | □ | 0 | 1 | 0 | □ | 0 | □ | □ | △ | □ | 0 | △ | 0 | 0 | 0 | △ | □ | □ | □ | □ | □ | 0 | |
| 10 | 0 | 0 | 1 | □ | □ | 1 | 1 | △ | 1 | △ | □ | 1 | 0 | 1 | △ | 0 | □ | □ | 0 | △ | △ | △ | 0 | 0 | 0 | □ | △ | △ | △ | △ | □ | 0 | |
| 11 | 0 | □ | □ | □ | □ | □ | □ | △ | □ | □ | □ | △ | □ | △ | □ | △ | □ | □ | △ | □ | △ | △ | △ | △ | 0 | △ | □ | □ | □ | □ | □ | 0 | |
| 12 | 0 | 0 | 1 | □ | □ | 1 | 1 | △ | 1 | 1 | □ | □ | □ | □ | □ | 0 | 1 | □ | △ | □ | △ | △ | 0 | 0 | 0 | □ | □ | △ | □ | □ | □ | 0 | |
| 13 | 0 | 0 | 1 | □ | □ | 1 | 1 | △ | 1 | 1 | □ | 1 | 0 | 1 | △ | 0 | □ | △ | 0 | △ | △ | △ | 0 | 0 | 0 | □ | △ | △ | △ | △ | □ | 0 | |
| 14 | 0 | 0 | 1 | □ | □ | 1 | 1 | △ | 1 | 1 | □ | 1 | 1 | 0 | □ | 0 | □ | □ | △ | □ | 0 | △ | 0 | 0 | 0 | △ | □ | □ | □ | □ | □ | 0 | |
| 15 | 0 | 0 | 1 | □ | □ | 1 | 1 | △ | 1 | 1 | □ | 1 | 1 | 1 | △ | 0 | □ | △ | 0 | △ | △ | △ | 0 | 0 | 0 | □ | △ | △ | △ | △ | □ | 0 | |
| 16 | △ | △ | □ | □ | □ | □ | □ | △ | □ | □ | □ | □ | □ | □ | □ | △ | □ | □ | △ | □ | △ | △ | △ | △ | 0 | △ | □ | □ | □ | □ | □ | △ | |
| 17 | □ | 0 | △ | □ | △ | △ | △ | □ | △ | △ | △ | △ | △ | △ | △ | △ | 0 | △ | □ | △ | △ | △ | □ | □ | □ | 0 | 0 | △ | △ | △ | 0 | △ | |
| 18 | 0 | 0 | 1 | □ | □ | 1 | 1 | △ | 1 | 1 | □ | 1 | 1 | 1 | 1 | □ | △ | □ | △ | □ | △ | △ | 0 | 0 | 0 | □ | □ | △ | □ | □ | □ | 0 | |
| 19 | 0 | △ | □ | □ | □ | □ | □ | △ | □ | □ | □ | □ | □ | □ | □ | □ | △ | □ | △ | □ | △ | △ | △ | 0 | 0 | □ | □ | □ | □ | □ | □ | △ | |
| 20 | □ | 0 | △ | △ | △ | △ | △ | □ | △ | △ | △ | △ | △ | △ | △ | △ | □ | △ | △ | △ | △ | □ | □ | △ | △ | △ | △ | □ | △ | △ | △ | □ | |
| 21 | △ | △ | □ | □ | □ | □ | □ | △ | □ | □ | □ | □ | □ | □ | □ | □ | △ | □ | □ | △ | △ | △ | △ | △ | △ | △ | □ | □ | □ | □ | □ | △ | |
| 22 | □ | 0 | △ | □ | □ | △ | △ | □ | △ | △ | △ | △ | △ | △ | △ | △ | □ | △ | △ | □ | △ | □ | □ | 0 | 0 | △ | △ | △ | △ | △ | △ | □ | |
| 23 | □ | △ | △ | △ | △ | △ | △ | □ | △ | △ | △ | △ | △ | △ | △ | △ | □ | △ | △ | □ | △ | □ | □ | △ | △ | △ | △ | △ | △ | △ | △ | □ | |
| 24 | □ | △ | 0 | △ | △ | 0 | 0 | □ | 0 | 0 | △ | 0 | 0 | 0 | 0 | △ | □ | 0 | △ | □ | △ | □ | □ | △ | △ | 0 | 0 | △ | △ | △ | 0 | □ | |
| 25 | □ | 0 | 0 | △ | △ | 0 | 0 | □ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | △ | □ | 0 | 0 | □ | △ | □ | □ | □ | □ | 0 | 0 | △ | △ | △ | 0 | 0 | |
| 26 | 1 | 0 | 0 | △ | △ | 0 | 0 | □ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | △ | □ | 0 | 0 | □ | △ | □ | □ | □ | □ | 0 | 0 | 0 | 0 | △ | 0 | 0 | |
| 27 | 0 | 0 | 1 | □ | □ | 1 | 1 | △ | 1 | 1 | □ | 1 | 1 | 1 | 1 | □ | △ | 1 | □ | △ | □ | △ | △ | 0 | 0 | 0 | △ | △ | △ | □ | □ | 0 | |
| 28 | 0 | △ | □ | □ | □ | □ | □ | △ | □ | □ | □ | □ | □ | □ | □ | □ | △ | □ | □ | △ | □ | △ | △ | △ | 0 | 0 | □ | □ | □ | □ | □ | 0 | |
| 29 | △ | △ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | □ | △ | △ | △ | △ | □ | □ | □ | □ | △ | △ | |
| 30 | 0 | 0 | 1 | □ | □ | 1 | 1 | △ | 1 | 1 | □ | 1 | 1 | 1 | 1 | □ | △ | 1 | □ | △ | □ | △ | △ | 0 | 0 | 0 | 1 | □ | □ | □ | □ | 0 | |
| 31 | △ | △ | □ | □ | □ | □ | □ | △ | □ | □ | □ | □ | □ | □ | □ | □ | △ | □ | □ | △ | □ | △ | △ | △ | △ | △ | □ | □ | □ | □ | □ | 0 | |
| 32 | 0 | △ | □ | □ | □ | □ | □ | △ | □ | □ | □ | □ | □ | □ | □ | □ | △ | □ | □ | △ | □ | △ | △ | △ | 0 | 0 | □ | 1 | □ | □ | 1 | 0 | |
| 33 | 0 | 1 | 0 | △ | △ | 0 | 0 | 0 | 0 | 0 | □ | 0 | 0 | 0 | 0 | △ | 0 | 0 | △ | 0 | △ | 0 | △ | △ | 0 | 0 | 0 | △ | △ | 0 | △ | △ | |
| △:0<NO<0.6, □:0.6≤NO<1; 主对角线上、下分别是以海拔和生境为资源等级计算的蚤生态位重叠指数 | |||||||||||||||||||||||||||||||||
生境生态位重叠指数中, 有意义的重叠有208对, 占比39.39%, 多为特新蚤指名亚种、特新蚤德钦亚种、云南栉眼蚤、迪庆额蚤、直缘双蚤、卷带倍蚤、细沟盖蚤与其他蚤种的重叠。NO=0的有98对, 占比18.56%, 多为稀有种之间的重叠, 最多的是与昌都盖蚤的重叠, 其次是与鼠兔角头蚤、介中怪蚤(Paradoxopsylla intermedius)、金沙江怪蚤(Paradoxopsylla jinshajiangensis jinshajiang jinshajiangensis)和长突怪蚤的重叠;NO=1的有70对, 占比13.26%, 最多的是与圆凹多毛蚤的重叠, 其次是二刺大锥蚤、直缘双蚤察里亚种(Amphipsylla tuta chaliensis)和二毫新蚤指名亚种(Neopsylla biseta biseta)的重叠, 除相关新蚤指名亚种和二次大锥蚤为常见种外, 其他均为稀有种。(表 3)
寄主生态位重叠指数中, 有意义的重叠有49对, 占比9.28%, NO=0的有270对, 占比51.14%, 最多的是与昌都盖蚤的种对, 其次是与鼠兔角头蚤和二刺大锥蚤的种对;NO=1的仅有4个种对, 占比0.19%, 分别是鼠兔角头蚤和昌都盖蚤、圆凹多毛蚤和纪氏叉蚤、泸水栉眼蚤和云南栉眼蚤、云南茸足蚤(Geusibia(Geusibia)yunnanensis)和卷带倍蚤的生态位重叠。(表 4)
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | 32 | 33 | |
| 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | △ | 0 | 0 | 0 | △ | 0 | △ | 0 | □ | □ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | △ | 0 | 0 | 0 | 0 | □ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | ||
| 3 | △ | △ | △ | 0 | △ | 0 | □ | □ | 0 | 1 | 0 | 0 | △ | 0 | 0 | 0 | △ | □ | 0 | △ | 0 | 0 | 0 | △ | △ | △ | 0 | 0 | 0 | 0 | |||
| 4 | □ | △ | 0 | △ | 0 | △ | □ | 0 | △ | □ | □ | □ | △ | 0 | △ | △ | △ | △ | △ | △ | △ | 0 | △ | △ | △ | 0 | 0 | △ | 0 | ||||
| 5 | △ | △ | △ | △ | △ | □ | △ | △ | □ | □ | □ | △ | 0 | △ | △ | □ | △ | △ | △ | △ | 0 | △ | △ | △ | △ | △ | △ | 0 | |||||
| 6 | 0 | △ | 0 | △ | △ | 0 | △ | △ | △ | △ | △ | 0 | △ | △ | △ | □ | △ | 0 | 0 | 0 | △ | △ | △ | 0 | 0 | △ | 0 | ||||||
| 7 | △ | 0 | 0 | 0 | 0 | 0 | △ | 0 | 0 | △ | 0 | 0 | 0 | △ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | △ | 0 | 0 | 0 | |||||||
| 8 | 0 | △ | △ | 0 | △ | △ | 0 | △ | □ | 0 | 0 | △ | △ | 0 | □ | △ | △ | 0 | △ | △ | △ | △ | △ | 0 | 0 | ||||||||
| 9 | 0 | 0 | □ | 0 | 0 | 0 | △ | 0 | 0 | 0 | △ | △ | 0 | 0 | 0 | 0 | 0 | □ | □ | 0 | 0 | △ | □ | 0 | |||||||||
| 10 | △ | 0 | □ | 0 | 0 | △ | 0 | 0 | 0 | △ | □ | △ | △ | 0 | 0 | 0 | △ | △ | △ | 0 | △ | △ | 0 | ||||||||||
| 11 | 0 | □ | □ | □ | □ | △ | 0 | △ | △ | □ | 0 | △ | 0 | 0 | 0 | △ | △ | △ | 0 | 0 | △ | 0 | |||||||||||
| 12 | 0 | 0 | 0 | △ | 0 | □ | □ | △ | △ | 0 | 0 | 0 | 0 | 0 | □ | □ | 0 | 0 | □ | □ | 0 | ||||||||||||
| 13 | 0 | 0 | △ | 0 | 0 | 0 | △ | □ | 0 | △ | 0 | 0 | 0 | △ | △ | △ | 0 | 0 | 0 | 0 | |||||||||||||
| 14 | □ | □ | △ | 0 | △ | △ | △ | △ | △ | 0 | 0 | 0 | 0 | △ | △ | △ | 0 | △ | 0 | ||||||||||||||
| 15 | 1 | △ | 0 | △ | 0 | △ | 0 | 0 | 0 | 0 | 0 | 0 | △ | 0 | 0 | 0 | △ | 0 | |||||||||||||||
| 16 | △ | 0 | △ | △ | △ | △ | △ | △ | △ | 0 | △ | △ | △ | 0 | △ | △ | 0 | ||||||||||||||||
| 17 | 0 | △ | □ | △ | △ | △ | △ | △ | 0 | 0 | △ | △ | △ | △ | △ | 0 | |||||||||||||||||
| 18 | □ | △ | △ | 0 | 0 | 0 | 0 | 0 | △ | 0 | 0 | 0 | 1 | 0 | 0 | ||||||||||||||||||
| 19 | △ | △ | 0 | 0 | △ | 0 | 0 | △ | △ | 0 | 0 | □ | △ | △ | |||||||||||||||||||
| 20 | △ | △ | △ | □ | △ | 0 | △ | △ | △ | △ | △ | △ | 0 | ||||||||||||||||||||
| 21 | △ | △ | △ | △ | △ | △ | △ | △ | △ | △ | △ | 0 | |||||||||||||||||||||
| 22 | △ | 0 | 0 | 0 | △ | 0 | △ | 0 | △ | △ | 0 | ||||||||||||||||||||||
| 23 | △ | △ | △ | △ | △ | △ | 0 | △ | 0 | 0 | |||||||||||||||||||||||
| 24 | △ | 0 | 0 | 0 | 0 | 0 | △ | 0 | □ | ||||||||||||||||||||||||
| 25 | □ | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||||||||||||||||||||||||
| 26 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||||||||||||||||||||||||||
| 27 | □ | △ | 0 | △ | □ | 0 | |||||||||||||||||||||||||||
| 28 | △ | 0 | △ | □ | 0 | ||||||||||||||||||||||||||||
| 29 | □ | 0 | 0 | 0 | |||||||||||||||||||||||||||||
| 30 | 0 | 0 | 0 | ||||||||||||||||||||||||||||||
| 31 | △ | 0 | |||||||||||||||||||||||||||||||
| 32 | 0 | ||||||||||||||||||||||||||||||||
| 主对角线以上是以寄主为资源等级计算的蚤生态位重叠指数 | |||||||||||||||||||||||||||||||||
生态位(Niche)理论作为探讨群落演变的经典理论, 在种间关系、群落结构、物种多样性及种群进化等研究领域被广泛应用[18]。生态位宽度和种间生态位重叠被认为是物种多样性及群落结构的决定因素, 也反映了所在群落的稳定性[19]。
本次调查结果显示, 迪庆额蚤和云南栉眼蚤为优势种, 但在三种资源轴中的生态位宽度值均不是最大, 这是因为生态位宽度值除了与数量有关之外, 更重要的是与分布有关, 某些具有数量优势的蚤种在分布中并不具有分配优势, 如云南栉眼蚤98.3%寄生于单一宿主西南绒鼠。三种资源轴中, 具有较大生态位宽度值的寄生蚤均为常见蚤种, 如特新蚤德钦亚种, 棕形额蚤等, 这些蚤种除了在数量上具有相对优势外, 在资源轴内部分布也具有优势, 为广生态位物种, 跨境分布广, 生态幅宽, 数量相对均匀, 具有很强的竞争优势。而生态位宽度值较小甚至为0的蚤种, 如人蚤、鼠兔角头蚤等蚤类, 仅在单一生境、海拔、寄主中有分布, 为狭生态位物种, 生态幅窄, 适应能力弱, 未利用等级资源, 在资源竞争中不具备竞争优势, 生态脆性增加[20], 容易因外部环境变化导致物种灭绝。
生境因子、海拔因子与资源的利用、种间种内的竞争、捕食与寄生、及生物的繁衍等息息相关[21-22]。本次调查中, 小兽体表寄生蚤生境选择以灌木丛为主, 这可能是因为该生境中水和食物资源充足, 环境隐蔽, 适宜小兽生存, 而寄生蚤的分布很大程度上依赖于小兽的分布[23]。在海拔选择上, 集中在3500m以上的中、高海拔带, 多为云南栉眼蚤和迪庆额蚤, 这与其主要宿主大耳姬鼠、西南绒鼠分布一致, 也是依赖小兽分布的另一种表现形式。在寄主资源轴上绒鼠怪蚤、棕形额蚤、迪庆额蚤占优势, 分别寄生于7、9、12种小兽, 是具有最多寄主的蚤种。而在寄主资源轴上, 蚤的生态位宽度值越大, 其生态幅越宽, 对寄主选择的特异性越低, 可寄生的宿主越多, 越倾向于是一个泛化种。生态位越窄, 对寄主选择性越高, 特化程度越高, 更倾向于是一个特化种, 甚至专一寄生。该资源轴与寄生蚤本身的特性, 如寄生方式、对寄主的选择性等高度相关。多维生态位宽度显示特新蚤德钦亚种和迪庆额蚤生态位宽度值最大, 而特新蚤作为云南省齐氏姬鼠-大绒鼠鼠疫疫源地的媒介蚤, 其在鼠疫传播中发挥着重要作用, 应重点关注, 加强防控。
生态位重叠指数反映的不仅是不同蚤种对于资源利用的相似度, 更表明了潜在的竞争程度, 重叠指数越高, 其选择越近似[24]。以海拔和生境为资源轴计算的生态位重叠指数显示:完全重叠和完全不重叠的种对, 多为稀有种与稀有种之间, 而有意义的重叠多为优势种、常见种、稀有种两两之间交叉重叠。这是因为优势蚤种和常见蚤种依赖于小兽的分布, 除了具有数量优势外, 还具有分布优势, 较宽的生态幅和较强的跨带分布能力让其竞争优势明显, 因此和其他蚤种重叠指数较高。而稀有种数量较少, 寄主相对单一, 甚至只寄生于某一种小兽, 导致选择同一小兽作为寄主的蚤种之间重叠指数高, 甚至完全重叠, 而选择不同小兽作为寄主的蚤种间重叠指数低, 甚至完全不重叠。在以寄主为资源轴的重叠指数表明:完全重叠的种对很少, 完全不重叠的种对很多, 更进一步表明蚤类寄生方式、对宿主的差异性选择对其生态位重叠指数的的影响较大。
重叠指数较高甚至完全重叠的物种能够在同一资源轴上同时存在是因为在其他资源轴上存在性状替代或分离, 如取食时间不同、活动时间不同、食性偏好不同[25-27]等。这种通过生态位分化减少种间竞争, 更合理的利用生态资源, 实现共存而没有导致相互驱离的现象[28]在物种间普遍存在。某一蚤种能与其他多个蚤种重叠, 说明其竞争能力强, 在生态竞争中更容易存活, 在外力导致群落重构过程中, 具有更大的竞争优势[29]。这种不同物种的适应性差异构成的群落物种的生态位分化, 是不同物种对环境适应性的结果[30]。随着资源轴的增加, 物种的生态位分化会逐渐增大, 以达到共存[31]。
在任一生态系统中, 物种间的相互作用都始终处于动态变化中, 通过不同功能生物间激烈的竞争或共生驱动形成不同资源轴上的生态位分化, 减少种间竞争, 进而实现共存, 形成相对稳定的群落结构[29]。但当环境中原有优势种因外部原因突然锐减, 其占位种会快速占据优势地位, 成为新的优势种, 如果其为重要的疾病传播媒介蚤种时, 可以导致疾病的爆发甚至快速流行。因此, 相对稳定的蚤类群落结构和其生态位格局对于蚤传疾病的防控相当重要。
4 结论本文运用生态位宽度值和生态位重叠指数分析了德钦县小型兽类体表寄生蚤的生态学特征。结果表明不同资源轴上寄生蚤的生态位宽度值不同, 重叠指数也不同, 这除了与调查环境本身的海拔、生境相关外, 还与寄生蚤的寄生方式和对寄主的差异性选择有关。该结果揭示了寄生蚤在不同生态因子中的生态位格局, 为鼠蚤监测提供指导, 对于有效防控蚤传疾病具有重大现实意义。
致谢: 云南省地方病防治所和德钦县疾病预防控制中心帮助调查,特此致谢。| [1] |
李登武, 张文辉, 任争争. 黄土沟壑区狼牙刺群落优势种群生态位研究. 应用生态学报, 2005, 16(12): 2231-2235. DOI:10.3321/j.issn:1001-9332.2005.12.002 |
| [2] |
陈林, 辛佳宁, 苏莹, 李月飞, 宋乃平, 王磊, 杨新国, 卞莹莹, 田娜. 异质生境对荒漠草原植物群落组成和种群生态位的影响. 生态学报, 2019, 39(17): 6187-6205. |
| [3] |
董雪, 李永华, 辛智鸣, 姚斌, 包岩峰, 脱登峰, 扆凡, 段瑞兵, 李新乐, 汪静, 孙志成, 王海, 陈旭. 敦煌西湖荒漠——湿地生态系统优势物种生态位研究. 生态学报, 2020, 40(19): 6841-6849. |
| [4] |
杜春红, 赵文红, 刘正祥, 刘云海, 提布只玛, 扎史品初, 宋志忠, 高子厚. 2012年云南省德钦县鼠疫疫源地调查实验室检测结果与分析. 疾病监测, 2013, 28(11): 917-919. DOI:10.3784/j.issn.1003-9961.2013.11.014 |
| [5] |
赵文红, 苏超, 高子厚, 洪梅, 段彪, 和星, 扎史品初. 云南省香格里拉和德钦两县鼠疫指示动物家犬F1抗体调查. 中国媒介生物学及控制杂志, 2015, 26(4): 411-413. |
| [6] |
唐梦琴, 孙娅蕾, 赵秋菊. 1971-2017年德钦县气候特点及变化特征分析. 现代农业科技, 2018(22): 218-218, 222-222. DOI:10.3969/j.issn.1007-5739.2018.22.131 |
| [7] |
杨丽天, 高子厚, 赵文红, 刘云海, 扎史品初, 提布只玛, 张文娟, 杜春红. 云南省德钦县宿主动物感染伯氏疏螺旋体情况初步调查. 中国热带医学, 2015, 15(4): 419-421, 433-433. |
| [8] |
陈星, 刘正祥, 杜春红, 洪梅, 龚正达, 邵宗体, 段兴德, 高子厚. 云南省农耕区小型兽类群落结构及分布格局. 应用生态学报, 2019, 30(6): 2039-2048. |
| [9] |
林元庆, 郑敏忠. 2001-2007年福州市鼠蚤调查分析. 海峡预防医学杂志, 2009, 15(2): 41-42. |
| [10] |
蒋志刚, 江建平, 王跃招, 张鹗, 张雁云, 李立立, 谢锋, 蔡波, 曹亮, 郑光美, 董路, 张正旺, 丁平, 罗振华, 丁长青, 马志军, 汤宋华, 曹文宣, 李春旺, 胡慧建, 马勇, 吴毅, 王应祥, 周开亚, 刘少英, 陈跃英, 李家堂, 冯祚建, 王燕, 王斌, 李成, 宋雪琳, 蔡蕾, 臧春鑫, 曾岩, 孟智斌, 方红霞, 平晓鸽. 中国脊椎动物红色名录. 生物多样性, 2016, 24(5): 501-551. |
| [11] |
罗泽珣; 陈卫; 高武; 王应祥; 李崇云; 李华. 《中国动物志》兽纲第六卷啮齿目下册仓鼠科. 北京市, 中国科学院动物研究所, 2000-11-17
|
| [12] |
解宝琦, 曾静凡. 云南蚤类志. 昆明: 云南科技出版社, 2000.
|
| [13] |
Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 1949: 3-55.
|
| [14] |
李德志, 石强, 臧润国, 王绪平, 盛丽娟, 朱志玲, 王长爱. 物种或种群生态位宽度与生态位重叠的计测模型. 林业科学, 2006, 42(7): 95-103. |
| [15] |
Pianka E R. The structure of lizard communities. Annual Review of Ecology and Systematics, 1973, 4: 53-74. DOI:10.1146/annurev.es.04.110173.000413 |
| [16] |
Wathne J A, Haug T, Lydersen C. Prey preference and niche overlap of ringed seals Phoca hispida and harp seals P. groenlandica in the Barents Sea. Marine Ecology Progress Series, 2000, 194: 233-239. DOI:10.3354/meps194233 |
| [17] |
徐正会, 杨比伦, 胡刚. 西双版纳片断山地雨林蚁科昆虫群落研究. 动物学研究, 1999, 20(4): 288-293. DOI:10.3321/j.issn:0254-5853.1999.04.009 |
| [18] |
Dewalt S J. Community Ecology. by Gary G. Mittelbach. Sunderland (Massachusetts): sinauer associates. $72.95(paper). xv+400 p.; ill.; author and subject indexes. ISBN: 978-0-87893-509-3. 2012. The Quarterly Review of Biology, 2013, 88(3): 237-238.
|
| [19] |
冯玉超, 郑晓阳, 王正宁, 刘博, 兰思仁. 杉木纯林和混交林林下草本层种群生态位特征. 生态与农村环境学报, 2019, 35(2): 217-224. |
| [20] |
徐高福, 余梅生, 章德三, 余世成, 黄丽飞. 千岛湖消落带生境现状及其不同类型植被重建与保护. 林业科技通讯, 2018(10): 21-26. |
| [21] |
肖厚贞, 辛吉武, 刘志威, 方佳, 贾德峰. 芒果炭疽病生境生态位. 热带农业科学, 2009, 29(6): 23-25. DOI:10.3969/j.issn.1009-2196.2009.06.006 |
| [22] |
王子迎, 檀根甲. 水稻纹枯病生境生态位研究. 安徽教育学院学报, 2003, 21(3): 66-69. DOI:10.3969/j.issn.1674-2273.2003.03.026 |
| [23] |
刘正祥, 高子厚, 尹家祥, 杜春红, 洪梅, 李玉琼, 宋志忠. 云南省居民区室内小型兽类体表蚤类与宿主关系研究. 中国媒介生物学及控制杂志, 2016, 27(2): 151-154. |
| [24] |
田杰. 剑川鼠疫自然疫源地27种蚤类生态位研究. 地方病通报, 1995, 10(3): 27-32. |
| [25] |
马英, 魏有文, 罗军, 鲁亮, 刘起勇. 青海省海东地区小型兽空间生态位分析. 动物学杂志, 2011, 46(04): 126-130. |
| [26] |
Lack D. Competition for food by birds of prey. Journal of Animal Ecology, 1946, 123-129. |
| [27] |
李凡, 徐炳庆, 吕振波, 王田田. 莱州湾鱼类群落优势种生态位. 生态学报, 2018, 38(14): 5195-5205. |
| [28] |
戎可, 司雨蕙, 潘麒嫣, 王欢. 同域分布3种啄木鸟冬季取食的生态位差异. 生态学报, 2018, 38(23): 8314-8323. |
| [29] |
盖珊珊, 赵文溪, 宋静静, 于道德, 刘莹, 王其翔, 周健. 小黑山岛人工鱼礁区许氏平鲉和大泷六线鱼的营养生态位研究. 生态学报, 2019, 39(18): 6923-6931. |
| [30] |
侯朝伟, 孙西艳, 刘永亮, 张晨, 张文静, 赵建民, 董志军. 烟台近海浮游动物优势种空间生态位研究. 生态学报, 2020, 40(16): 5822-5833. |
| [31] |
郭磊, 韦博良, 胡金涛, 贾宏汝, 叶永忠, 袁志良. 基于两个不同资源轴上苔藓植物生态位分析. 生态学报, 2017, 37(21): 7266-7276. |