 2021, Vol. 41
2021, Vol. 41文章信息
- 曹恭祥, 王云霓, 季蒙, 郭中, 李艳慧, 耿庆春, 包瑞, 刘佳
- CAO Gongxiang, WANG Yunni, JI Meng, GUO Zhong, LI Yanhui, GENG Qingchun, BAO Rui, LIU Jia
- 呼伦贝尔沙地樟子松林净初级生产力对气候变化的响应
- Response of net primary productivity of Pinus sylvestris var. mongolica plantation to climate change in Hulunbuir sandy land
- 生态学报. 2021, 41(13): 5352-5359
- Acta Ecologica Sinica. 2021, 41(13): 5352-5359
- http://dx.doi.org/10.5846/stxb201908201738
-
文章历史
- 收稿日期: 2019-08-20
- 修订日期: 2021-04-12





2. 呼和浩特市赛罕区林业局, 呼和浩特 010010;
3. 红花尔基林业局, 海拉尔 021112
2. The Forestry Bureau of SaiHan District in Huhhot, Hohhot 010010, China;
3. The Forestry Bureau of Honghuaerji, Hailaer 021112, China
气候变化会深刻影响着陆地生态系统的结构、组成、功能[1-2], 森林生态系统作为陆地生态系统的重要组成部分, 其对气候变化的响应对维持陆地生态系统平衡具有重要意义[3-4]。森林生物量和生产力是森林生态系统的重要特性, 可用于指示气候变化对森林的影响[5-6]。已有关于森林生物量或生产力的研究主要集中在不同植被类型的静态对比、利用空间代替时间法分析同一植被生产力变化特征及少量森林净初级生产力对气候因子的响应。如, 王云霓等[5]对宁夏六盘山华山松(Pinus armandii)林、油松(Pinus tabulaeformis)林、华北落叶松(Larix principis-rupprechtii)林、程瑞梅等[6]对鸡公山马尾松(Pinus massoniana)林、于健等[7]对长白山红松(Pinus koraiensis)林的年净初级生产力变化特征及其对气候因子的响应进行了分析。然而, 受立地条件、树种、林龄、人为干扰等影响, 森林生物量和生产力表现出明显的种间差异[8-10]、空间差异[11-12]和时间差异[6]。为了全面认识森林生产力对气候变化的响应, 就需扩大研究不同区域不同树种的生物量和生产力变化特征及其对气候变化的响应。
在气候变化的背景下, 呼伦贝尔沙地樟子松林净初级生产力的变化特征及其对气候变化的响应如何, 在我国东北地区森林-草原过渡带上, 仍缺乏森林生态系统与气候变化响应关系研究的相关报道。为此, 本研究以内蒙古呼伦贝尔沙地樟子松(Pinus sylvestris var. mongolica)人工林为研究对象, 利用生物量经验方程和年轮生态学方法, 计算樟子松林的生物量和净初级生产力, 并分析樟子松林净初级生产力对气候因子变化的响应特征, 对探索全球气候变化下我国半干旱区森林草原过渡带上樟子松林生长特征具有重要意义, 也为应对气候变化的森林精准经营提供理论基础。
1 研究区概况本研究的采样点位于内蒙古呼仑贝尔红花尔基自然保护区(47°36′—48°35′N, 118°58′—120°32′E)。该地区属于寒温带大陆性气候, 地处干旱地区;冬季漫长, 夏季短暂。多年(1965—2014年)平均气温-1.3 ℃, 1月最冷, 极端最低气温-46.5 ℃;7月最热, 极端最高气温37.7 ℃, 平均相对湿度为61.7%;降水集中在6—8月, 多年平均降水量为330.1 mm;日照时数为2857.3 h, 年蒸发量1480.2 mm。土壤一般呈酸性或微酸性, 质地以沙土、淋溶黑土、薄层粗骨土等为主。
本文的研究对象为樟子松, 该树种深根性、抗逆性强、生长较快, 对立地条件要求不高, 适应性强, 是欧亚温带广泛分布赤松(Pinus sylvestris)的变种。红花尔基是樟子松的最大天然分布区, 樟子松面积13.58×104 hm2, 其中天然林10.91×104 hm2, 人工林2.67×104 hm2。红花尔基樟子松多为纯林, 在阴坡凹地混生有白桦(Betula platyphylla)、山杨(Populus davidiana)等伴生树种。灌木主要有山刺玫(Rosa davurica)、小叶杜香(Ledum palustre)、绣线菊(Spiraea salicifolia)、兴安杜鹃(Rhododendron dauricum)等。草本植物发育良好, 盖度较大, 主要有苔草(Carex stenophylloides)、兴安野青茅(Calamagrostic turczaninowii)、莎草(Cyperus rotundus)、羊草(Leymus chinensis)、广布野豌豆(Vicia cracca)、百蕊草(Thesium chinense)、蕨(Pteridium aquilum)、小叶章(Calamagrostis angustifolia)、矮山黧豆(Lathyrus humilis)等。

|
| 图 1 研究区1965—2014年的年平均温度和年降水量 Fig. 1 Changes of annual mean temperature and precipitation from 1965 to 2014 |
在红花尔基樟子松林区选择生长良好、林龄较大的樟子松人工林, 设置观测样地3个, 样地大小30 m×30 m, 调查样地的海拔、坡度等立地因子, 并进行每木检尺, 调查胸径、树高、冠幅、枝下高等植被特征, 样地的基本特征见表 1和图 2。樟子松人工林均为43年生, 三个样地的林分密度分别为782株/hm2、667株/hm2、625株/hm2, 密度相对较小, 但林冠郁闭度较大, 达到0.86以上。如图 2所示, Ⅰ样地中, 樟子松个体的平均胸径为25.83 cm, 以分布在24—28 cm径级范围内最多, 比例为60.00%, 胸径在31 cm以上的个体比例为8.57%。Ⅱ样地, 樟子松个体的平均胸径为22.53 cm, 以分布在22—28 cm径级范围内最多, 比例为70.00%, 胸径在29 cm以上的个体比例为1.67%, 没有出现径级31 cm以上的个体, 胸径在20 cm径阶及以下的个体比例为28.33%。Ⅲ样地, 樟子松个体的平均胸径为26.54 cm, 以分布在24—28 cm径级范围内最多, 比例为58.93%, 胸径在31 cm以上的个体比例为14.29%;该样地内, 出现了径级为33 cm以上的个体, 比例为5.36%。平均树高为12.89—13.58 m, 冠幅直径平均为5.21—5.40 m, 最大冠幅直径达到9.00 m。林下灌木零星分布或者没有, Ⅰ号样地内有2丛绣线菊, 高度分别为46.60 cm和51.90 cm, 其他样地无灌木分布。林下草本层发育较好, 盖度达到85%以上, 主要种类有东方草莓(Fragaria orientalis)、苔草、蕨、百蕊草、兴安野青茅、广布野豌豆等。
| 编号 Plot No. |
海拔 Altitude/m |
坡度 Slope degree/(°) |
乔木层Tree layer | 草本层Herbaceous layer | |||||
| 密度 Density/(株/hm2) |
平均胸径 Mean DBH/cm |
树高 Mean tree height/m |
郁闭度 Canopy density |
优势种 Dominant species |
盖度 Coverage/% |
||||
| Ⅰ | 810 | < 3 | 782 | 25.83±2.98 | 12.9±1.25 | 0.90 | 东方草莓Fragaria orientalis | 86 | |
| Ⅱ | 835 | < 3 | 667 | 22.53±3.24 | 13.4±1.09 | 0.86 | 苔草Carex stenophylloides | 85 | |
| Ⅲ | 829 | < 3 | 625 | 26.54±3.75 | 13.6±1.30 | 0.89 | 苔草Carex stenophylloides | 89 | |
| DBH: Diameter breast height | |||||||||

|
| 图 2 樟子松林样地径级分布 Fig. 2 The trees diameter distribution of Pinus sylvestris var. mongolica plantation plot |
在样地调查的基础上, 按照径级选择生长良好、干形较直的标准木20株, 在标准木胸高处(1.3 m)垂直交叉钻取2个树芯。按照树木年轮处理的基本程序, 即自然干燥、固定、打磨[5], 并初步检查可能存在的伪轮和遗失年轮。利用Lintab 5年轮分析仪(Frank Rinn, 德国)测量宽度(精度为0.01 mm), 并利用折线图比法和COFECHA程序[13]对定年准确性和测量结果进行合理性检验, 选择与主序列相关系数较大、奇异点少的样芯, 之后利用ARSTAN程序去趋势。
2.3 生物量和净初级生产力的估算利用调查时测定胸径D2017依次减去历年2个树芯年轮宽度之和dn(cm), 得到历年单木第n年的胸径Dn(cm), 见公式1。应用研究区38棵解析木数据拟合樟子松带皮胸径(Db, cm)和去皮胸径(Dw, cm)函数关系(Db=1.6815+1.0118Dw, R2=0.99), 将樟子松历年去皮胸径转换成带皮胸径。基于红花尔基林场樟子松解析木数据建立的单株各部分(干、枝、叶)生物量(kg)与胸径D(cm)的函数关系[14], 计算样树的地上生物量, 见公式(2)—(4);根据单株根系生物量W根(kg)与胸径D(cm)的关系计算根系生物量[15], 见公式(5);最后结合样地面积S(m2)和株数m(棵)计算单位面积上生物量W(t/hm2), 见公式(6)。
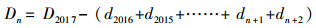
|
(1) |
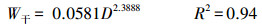
|
(2) |
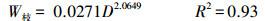
|
(3) |

|
(4) |
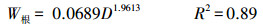
|
(5) |
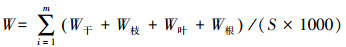
|
(6) |
式中, Dn是第n年的胸径, dn是第n年的两个不同树芯年轮宽度之和, Db为樟子松带皮直径, Dw为樟子松去皮胸径, W为样地生物量, W干为单株樟子松树干生物量, W枝为单株樟子松树枝生物量, W叶为单株樟子松树叶生物量, W根为单株樟子松树根生物量, m为每个调查样地的樟子松株数。
样地的年净初级生产力NPP为样地历年单位面积上的生物量之差[5]。
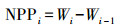
|
(7) |
式中, Wi是第i年的生物量(t/hm2), NPPi是第i年的净初级生产力(t hm-2 a-1), i表示年份。
2.4 数据处理气象数据为鄂温克旗气象站多年的观测数据, 包括月均气温、月均最高气温、月均最低气温、月降水量和月均相对湿度等5个气象因素。利用Mann-Kandell和Double-mass analysis方法对气象数据的均一性进行检验, 结果表明气象资料变化相对均一, 无随机突变和随机变化, 可以用来代表研究区自然气候的变化。考虑到上一年气象特征对当年的树木生长的影响[12, 16-17], 本文在分析净初级生产力与气象因子的关系时, 选择上一年6月到当年12月的气象指标。
应用Excel 2010和SPSS 18软件进行数据处理, 利用Pearson相关分析樟子松人工林净初级生产力与气象因子的关系。
3 结果 3.1 生物量的年际变化樟子松林生物量的年际变化如图 3所示。随着林龄的增加, 樟子松林生物量变化总体呈现缓慢增加—快速增加—缓慢增加的趋势, 樟子松林生物量从1977年的4.83 t/hm2到2017年的生物量为167.95 t/hm2, 近41年间增长超过了34倍, 生物量W(t/hm2)的增长随林龄N变化以Logistic模型(W=170/(1+11.081EXP(-0.129N)), R2=0.99)拟合较好。

|
| 图 3 樟子松人工林生物量的年际变化 Fig. 3 Interannual changes of biomass of Pinus sylvestris var. mongolica plantation |
樟子松林净初级生产力年际变化总体表现出先增加后减小、最后趋于稳定的趋势, 但年际间波动存在差异, 见图 4。在1977—1987年是快速上升, 1987年净初级生产力达到5.99 t hm-2 a-1, 但1988—1991年间净初级生产力呈持续降低趋势, 降低到4.33 t hm-2 a-1;在1992回升到6.01 t hm-2 a-1, 1993年的净初级生产力最高, 达到6.13 t hm-2 a-1;1994—2006年是下降的趋势, 但是在下降中又有波动, 净初级生产力在2006年降到最低, 为2.63 t hm-2 a-1;之后呈现波动上升的趋势, 但上升幅度很小。多年平均年净初级生产力为4.08 t hm-2 a-1, 变异系数为0.26。

|
| 图 4 樟子松人工林净初级生产力的年际变化 Fig. 4 Interannual changes of net primary productivity of Pinus sylvestris var. mongolica plantation |
樟子松林净初级生产力与气象指标的相关性见图 5。樟子松林净初级生产力对气象因子的响应具有一定的“滞后”。净初级生产力与上一年8月、12月的降水分别呈显著性正、负相关(P<0.05), 相关系数分别为0.336和0.324;与上一年6月的月均最高温、8月的月均最低温及7月、9月的月均温和月均最高温均呈极显著(P<0.01)或显著(P<0.05)负相关, 相关系数均大于0.33;与上一年8月、9月的月均湿度呈极显著负相关(P<0.01), 相关系数分别达到0.602和0.505。说明前一年12月降水、8—9月湿度和6—9月的温度指标对樟子松林净初级生产力有着显著影响。

|
| 图 5 樟子松林生产力与月份气象因子的相关系数 Fig. 5 Correlation coefficients between productivity of Pinus sylvestris var. mongolica plantation and monthly meteorological factors 虚线表示相关分析显著性水平线(P<0.05), 实线表示相关分析极显著性水平线(P<0.01), 月份前的“-”表示前一年 |
从图 5还可以看出, 樟子松林净初级生产力与当年3月降水呈显著正相关(P<0.05), 说明生长季前期降水对樟子松林生产力的影响较大;当年1月、6月、8月、9月、12月的月均湿度显著影响着樟子松林生产力。樟子松林净初级生产力与当年大多月份平均气温成负相关, 与8月、9月的平均气温分别显著、极显著负相关, 相关系数分别达到0.348、0.414;与8月的月均最低温、6月月均最高温呈显著负相关(P<0.05)。
4 讨论 4.1 樟子松林的生物量和生产力研究区樟子松林生物量从1977年的4.83 t/hm2到2017年的生物量为167.95 t/hm2(地上生物量为133.55 t/hm2), 近41年间增长超过了34倍;多年间净初级生产力最高达到6.13 t hm-2 a-1, 最低为2.63 t hm-2 a-1, 多年平均净初级生产力为4.08 t hm-2 a-1。本研究中樟子松林的地上生物量和生产力低于黑龙江省西部地区的[18], 后者23年生樟子松林的地上生物量和生产力分别为154.07 t/hm2和14.72 t hm-2 a-1, 但远高于黑龙江大兴安岭45年生的(94.87 t/hm2)、小兴安岭45年生的(80.50 t/hm2)[19]、塞罕坝地区45年生左右樟子松林地上生物量(94.20 t/hm2)[20]和内蒙古科尔沁沙地35年生的(86.45 t/hm2)[21]。
生产力差异的可能是由立地条件、林龄、林分密度、气候等因素综合引起。如, 袁立敏等[21]研究认为:科尔沁沙地樟子松林生物量随着林龄的增加而增大;王云霓等[22]发现华北落叶松同龄人工林生物量具有显著的坡位差异;贾忠奎等[23]研究发现塞罕坝华北落叶松同龄人工林生产力具有显著的坡向差异;彭舜磊等[24]对天童地区常绿阔叶林、吕振刚等[25]对河北省华北落叶松的研究都认为:气候变化显著影响着林分生产力;李俊霞等[12]对大兴安岭林区南北部樟子松生长研究表明:高温是导致南部樟子松生长量降低的原因, 而北部樟子松生长量增加的原因是4—9月平均最低温与降水量的相互作用;李露露等[26]研究发现樟子松径向生长指示的森林生产力在地理空间水平上随着降水的增加而显著增加, 呈明显的梯度变化。
4.2 气候因子对生产力的影响 4.2.1 降水和湿度对生产力的影响本研究中, 樟子松林净初级生产力与上一年8月、12月和当年3月的月降水量显著相关;与上一年8月、9月、12月和当年1月、6月、8月、9月的月均湿度显著相关, 这与前人研究较为一致。如王云霓等[5]研究发现;华北落叶松林净初级生产力与上一年9月和当年4月的月降水量呈显著正相关;尚建勋等[27]认为:樟子松径向生长与各月月降水量多呈正相关关系, 特别当年5—8月降水量呈显著正相关关系。研究区3月气温回升, 树木细胞开始分裂, 此时较多的降水, 可为樟子松萌芽生长储备较多的土壤水分[28], 有利于提高树木生产潜力。此外, 虽然在樟子松关键生长时期(5—8月), 当年降水量与林分生产力呈正相关关系, 但没有达到显著或极显著相关, 这可能与研究样地立地条件有关, 本研究样地均位于土壤层相对较厚的坡下, 受降水坡面再分配影响, 土壤层水分相对丰富, 立地条件优越, 所以樟子松林净初级生产力较高, 大气直接降水量的多少不是制约净初级生产力的主导因素。另外, 樟子松林净初级生产力与生长季月均湿度之间的正相关关系及与生长季月均温负相关系数的存在, 也进一步说明温度可能通过植被的蒸腾作用及土壤水分的蒸发而间接影响树木的生长[29]。
在本研究中, 上一年6—9月降水量与樟子松林净初级生产力呈正相关, 而上年10月至当年2月降水量却与生产力呈负相关。这可能与降水形态有关[28], 6—9月降水充足, 樟子松树体内储存水分就较多, 为下一年生长提供贮备, 且适量的土壤水分也可保证树木安全越冬[28];而上一年10月到翌年2月降水以固态形式为主[27], 容易形成雪害, 对树木造成物理伤害, 引起树木倾斜、折断和倒伏, 在一定程度上影响其生长[5]。
4.2.2 温度对生产力的影响樟子松林净初级生产力与上一年及当年5—9月的各项温度指标均成负相关。这与前人的研究基本一致[30-31]。王云霓等[5]研究发现:华山松天然林净初级生产力与当年1—8月月均温和月均最高温及2—7月月均最低温呈负相关关系;程瑞梅等[6]等发现北亚热带马尾松净生产力与当年5月平均气温呈负相关;张先亮等[3]对大兴安岭山地樟子松的研究认为:樟子松径向生长与上一年6—9月和当年4—8月的月均气温、月均最低温、月均最高温均呈显著负相关, 温度是樟子松生长的主要限制因子;尚建勋等[27]认为:樟子松径向生长与生长季前期和生长季的各项温度指标均成负相关, 月均最高气温的升高不利于樟子松的径向生长;李露露等[26]认为生长季温度升高不利于辽宁省樟子松生长, 是因为升温导致的水分亏缺及高温胁迫抑制樟子松的生长;吴祥定和邵雪梅[32]研究结果显示:上一年10月到当年4月的月均气温与树木径向生长呈负相关, 并认为引起以上响应关系可能与树木生理过程有关;而Mäkinen[30]和Rolland[31]认为:温度升高导致蒸散增多, 加剧水分胁迫, 此时降水若不能及时补充土壤水分, 只能使用树木体内储存水分, 进而影响次年的生长。
此外, 可能与树种的生物学特性有关, 樟子松虽耐寒抗旱, 但温度生理阈值较小, 生长季气温升高导致蒸腾加强, 容易引起树木生理干旱, 即高温干燥会限制其生长, 本研究中樟子松林净初级生产力与降水的相关性较低、而温度及生长季月均湿度的增加显著影响着樟子松林生产力就是例证, 所以温度升高引起的干旱胁迫或树木生理干旱是抑制研究区樟子松林净初级生产力的主要原因, 在其他研究中也发现类似现象。如:Liu等[4]研究认为:由于温度升高导致的生长季水分需求增加, 导致半干旱地区的树木生长速率下降, 且树木死亡率增加;张先亮等[3]对大兴安岭山地樟子松、李俊霞等[12]对大兴安岭南侧樟子松、李露露等[26]对辽宁省樟子松、Yu等[11]对长白山低海拔地区红松、Schuster和Oberhuber[33]对奥地利中部地区三种针叶林、Buermann等[34]对欧亚北方森林的研究均显示:温度升高引起的生理干旱是抑制树木生长的主要原因。
5 结论随着林龄的增加, 樟子松人工林生物量逐渐增加, 从1977年的4.83 t/hm2到2017年的167.95 t/hm2。樟子松林净初级生产力的年际差异较大, 总体趋势呈先增加后减小、最后趋于稳定, 多年平均生产力为4.08 t hm-2 a-1。气候因子是影响樟子松林净初级生产力年际变化的主要原因, 特别是3月和12月的降水、8—9月的湿度及6—9月的温度, 即生长季初期及冬季的降水、生长季的温度及生长季后期的湿度是影响呼伦贝尔沙地樟子松林净初级生产力的主要因素。根据中国及研究区气候变化的趋势来看, 该地区未来温度将会升高, 气候暖干化, 将加重生长季温度升高引起的干旱胁迫, 限制樟子松的生长, 但由于降水格局不同, 气候变暖对樟子松林净初级生产力的影响还需要进一步的研究。另外, 有研究发现:树木生长量下降可能因为碳获取不足[35-36], 后续的研究还需要考虑二氧化碳浓度升高对樟子松林净初级生产力的影响。
| [1] |
Liang E Y, Leuschner C, Dulamsuren C, Wagner B, Hauck M. Global warming-related tree growth decline and mortality on the north-eastern Tibetan plateau. Climatic Change, 2016, 134(1/2): 163-176. |
| [2] |
晏寒冰, 彭丽潭, 唐旭清. 基于气候变化的东北地区森林树种分布预测建模与影响分析. 林业科学, 2014, 50(5): 132-139. |
| [3] |
张先亮, 何兴元, 陈振举, 崔明星, 黎娜. 大兴安岭山地樟子松径向生长对气候变暖的响应——以满归地区为例. 应用生态学报, 2011, 22(12): 3101-3108. |
| [4] |
Liu H Y, Williams A P, Allen C D, Guo D L, Wu X C, Anenkhonov O A, Hang E Y, Sandanov D V, Yin Y, Qi Z H, Badmaeva N K. Rapid warming accelerates tree growth decline in semi-arid forests of Inner Asia. Global Change Biology, 2013, 19(8): 2500-2510. DOI:10.1111/gcb.12217 |
| [5] |
王云霓, 熊伟, 王彦辉, 于澎涛, 曹恭祥, 郭浩, 徐丽宏, 左海军. 宁夏六盘山三种针叶林初级净生产力年际变化及其气象因子响应. 生态学报, 2013, 33(13): 4002-4010. |
| [6] |
程瑞梅, 封晓辉, 肖文发, 王瑞丽, 王晓荣, 杜化堂. 北亚热带马尾松净生产力对气候变化的响应. 生态学报, 2011, 31(8): 2086-2095. |
| [7] |
于健, 罗春旺, 徐倩倩, 孟盛旺, 李俊清, 刘琪璟. 长白山原始林红松径向生长及林分碳汇潜力. 生态学报, 2016, 36(9): 2626-2636. |
| [8] |
Ni J. Forest productivity of the Altay and Tianshan Mountains in the dryland, northwestern China. Forest Ecology and Management, 2004, 202(1/3): 13-22. |
| [9] |
Yamashita T, Kasuya N, Nishimura S, Takeda H. Comparison of two coniferous plantations in central Japan with respect to forest productivity, growth phenology and soil nitrogen dynamics. Forest Ecology and Management, 2004, 200(1/3): 215-226. |
| [10] |
胡海清, 罗碧珍, 魏书精, 魏书威, 文正敏, 孙龙, 罗斯生, 王立明, 马洪斌. 大兴安岭5种典型林型森林生物碳储量. 生态学报, 2015, 35(17): 5745-5760. |
| [11] |
Yu D P, Liu J Q, Lewis B J, Zhou L, Zhou W M, Fang X M, Wei Y W, Jiang S W, Dai L M. Spatial variation and temporal instability in the climate-growth relationship of Korean pine in the Changbai Mountain region of Northeast China. Forest Ecology and Management, 2013, 300: 96-105. DOI:10.1016/j.foreco.2012.06.032 |
| [12] |
李俊霞, 白学平, 张先亮, 常永兴, 陆旭, 赵学鹏, 陈振举. 大兴安岭林区南、北部天然樟子松生长对气候变化的响应差异. 生态学报, 2017, 37(21): 7232-7241. |
| [13] |
Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983, 43: 69-78. |
| [14] |
丛俊霞, 郑晓, 朱教君, 宋立宁, 高西宁, 李秀芬, 闫涛. 沙地樟子松天然林地上碳储量估算及其空间分布特征. 生态学杂志, 2017, 36(11): 2997-3007. |
| [15] |
董利虎. 黑龙江省主要树种相容性生物量模型研究[D]. 哈尔滨: 东北林业大学, 2012: 20-22.
|
| [16] |
高露双, 王晓明, 赵秀海. 长白山过渡带红松和鱼鳞云杉径向生长对气候因子的响应. 植物生态学报, 2011, 35(1): 27-34. |
| [17] |
王晓春, 宋来萍, 张远东. 大兴安岭北部樟子松树木生长与气候因子的关系. 植物生态学报, 2011, 35(3): 294-302. |
| [18] |
苑增武, 丁先山, 李成烈, 赵玉恒, 曹志伟. 樟子松人工林生物生产力与密度的关系. 东北林业大学学报, 2000, 28(1): 21-24. DOI:10.3969/j.issn.1000-5382.2000.01.006 |
| [19] |
胡海清, 罗碧珍, 魏书精, 魏书威, 孙龙, 罗斯生, 马洪斌. 小兴安岭7种典型林型林分生物量碳密度与固碳能力. 植物生态学报, 2015, 39(2): 140-158. |
| [20] |
李晓莎, 楚聪颖, 许中旗, 梅琳, 张岩, 吴强, 程顺, 崔同祥. 河北省塞罕坝地区樟子松人工林的生物量. 河北林果研究, 2016, 31(3): 230-234. |
| [21] |
袁立敏, 闫德仁, 王熠青, 姜鹏, 刘阳, 党晓宏. 沙地樟子松人工林碳储量研究. 内蒙古林业科技, 2011, 37(1): 9-13. DOI:10.3969/j.issn.1007-4066.2011.01.003 |
| [22] |
王云霓, 邓秀秀, 王彦辉, 曹恭祥, 于澎涛, 熊伟, 徐丽宏. 宁夏六盘山香水河小流域华北落叶松人工林乔木层生物量的坡面变化和尺度效应. 林业科学研究, 2015, 28(5): 701-707. DOI:10.3969/j.issn.1001-1498.2015.05.015 |
| [23] |
贾忠奎, 公宁宁, 姚凯, 司瑞雪, 程顺, 王立东, 张田田, 游伟斌, 王西洋, 刘杰. 塞罕坝华北落叶松人工林生产力对坡向的响应. 西北林学院学报, 2012, 27(4): 1-6. DOI:10.3969/j.issn.1001-7461.2012.04.01 |
| [24] |
彭舜磊, 由文辉, 郑泽梅, 王良衍. 近60年气候变化对天童地区常绿阔叶林净初级生产力的影响. 生态学杂志, 2011, 30(3): 502-507. |
| [25] |
吕振刚, 李文博, 黄选瑞, 张志东. 气候变化情景下基于潜在NPP的河北省华北落叶松生长适宜性. 林业科学, 2019, 55(11): 37-44. DOI:10.11707/j.1001-7488.20191105 |
| [26] |
李露露, 李丽光, 陈振举, 周永斌, 张先亮, 自学平, 常永兴, 肖建强. 辽宁省人工林樟子松径向生长对水热梯度变化的响应. 生态学报, 2015, 35(13): 4508-4517. |
| [27] |
尚建勋, 时忠杰, 高吉喜, 徐丽宏, 吕世海, 冯朝阳, 王鲁秀. 呼伦贝尔沙地樟子松年轮生长对气候变化的响应. 生态学报, 2012, 32(4): 1077-1084. |
| [28] |
曹恭祥, 郭中, 王云霓, 季蒙, 韩珊珊, 耿庆春, 冯丽红, 刘海利. 呼伦贝尔沙地樟子松人工林乔木层固碳速率及其对气象因子的响应. 生态学杂志, 2020, 39(4): 1082-1090. |
| [29] |
Liang E Y, Shao X M, Hu Y X, Lin J X. Dendroclimatic evaluation of climate-growth relationships of Meyer spruce (Picea meyeri) on a sandy substrate in semi-arid grassland, North China. Trees, 2001, 15(4): 230-235. DOI:10.1007/s004680100097 |
| [30] |
Mäkinen H, Nöjd P, Mielikäinen K. Climatic signal in annual growth variation in damaged and healthy stands of Norway spruce[Picea abies (L.) Karst.] in southern Finland. Trees, 2001, 15(3): 177-185. DOI:10.1007/s004680100089 |
| [31] |
Rolland C. Tree-ring and climate relationships for Abies alba in the internal Alps. Tree-Ring Bulletin, 1993, 53: 1-11. |
| [32] |
吴祥定, 邵雪梅. 采用树轮宽度资料分析气候变化对树木生长量影响的尝试. 地理学报, 1996, 51(S1): 92-101. |
| [33] |
Schuster R, Oberhuber W. Drought sensitivity of three co-occurring conifers within a dry inner Alpine environment. Trees, 2013, 27(1): 61-69. DOI:10.1007/s00468-012-0768-6 |
| [34] |
Buermann W, Parida B, Jung M, MacDonald G M, Tucker C J, Reichstein M. Recent shift in Eurasian boreal forest greening response may be associated with warmer and drier summers. Geophysical Research Letters, 2014, 41(6): 1995-2002. DOI:10.1002/2014GL059450 |
| [35] |
Mencuccini M, Martínez-Vilalta J, Vanderklein D, Hamid H A, Korakaki E, Lee S, Michiels B. Size-mediated ageing reduces vigour in trees. Ecology Letters, 2005, 8(11): 1183-1190. DOI:10.1111/j.1461-0248.2005.00819.x |
| [36] |
Bond-Lamberty B, Wang C K, Gower S T. Net primary production and net ecosystem production of a boreal black spruce wildfire chronosequence. Global Change Biology, 2004, 10(4): 473-487. DOI:10.1111/j.1529-8817.2003.0742.x |