 2021, Vol. 41
2021, Vol. 41文章信息
- 丁敬坤, 李加琦, 薛素燕, 张雯雯, 霍恩泽, 马占飞, 于文涵, 毛玉泽
- DING Jingkun, LI Jiaqi, XUE Suyan, ZHANG Wenwen, HUO Enze, MA Zhanfei, YU Wenhan, MAO Yuze
- 莱州湾小清河口近岸海域底栖生境健康评价
- Health assessment for benthic habitats of macrobenthos in the sea area adjacent to the Xiaoqing River estuary, Laizhou Bay
- 生态学报. 2021, 41(12): 4806-4817
- Acta Ecologica Sinica. 2021, 41(12): 4806-4817
- http://dx.doi.org/10.5846/stxb202002290369
-
文章历史
- 收稿日期: 2020-02-29
- 网络出版日期: 2021-04-21

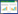




2. 青岛海洋科学与技术试点国家实验室海洋生态与环境科学功能实验室, 青岛 266237;
3. 上海海洋大学水产科学国家级实验教学示范中心, 上海 201306
2. Laboratory for Marine Ecology and Environmental Science, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266237, China;
3. National Demonstration Center for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306, China
海洋生态系统是生物圈的重要组成部分, 不仅为人类提供了大量宝贵的食物来源与服务, 而且对维持全球生态系统的动态平衡具有至关重要的作用, 这对人类的生存和发展具有非常重要的意义。因此, 海洋生态系统健康日益受到国内外学者的关注, 海洋生态系统健康评价成为全球生态学和海洋学领域研究的热点问题。21世纪后, 生态系统健康评价在河口、海湾、近海等海洋生态系统中的应用逐渐增多。2005年国家海洋局在《近岸海洋生态健康评价指南》中提出生态系统健康是指生态系统保持其自然属性, 维持生物多样性和关键生态过程稳定并持续发挥其服务功能的能力, 提出用水环境、沉积环境、生物残毒、栖息地和生物5类指标评价河口及海湾生态系统健康[1]。基于底栖群落的生物指数已经成为生态系统健康评估和相关决策的有效工具, 根据底栖生物群落和数量的空间变化与环境扰动之间有可预测的关系, 目前已经建立了多种底栖生物指数, 主要包括一些比较传统的生物指数, 如物种丰富度、多样性指数和优势度, 以及新开发的生物指数, 如海洋生物指数(BI), AZTI海洋生物指数(AMBI), 多变量AMBI(M-AMBI)[2]和底栖质量指数(BQI), 特别是基于大型底栖生物的AMBI和M-AMBI指数已经广泛应用于河口和沿海水域[3]。
莱州湾是渤海三大海湾之一, 位于山东半岛北部, 湾内水深较浅, 大部分海域水深小于10m, 最大水深仅18m, 是鱼、虾、蟹以及贝类等经济生物的主要繁育场, 也是山东省重要的水产品养殖基地。河口是地球上最具生产力和最重要的环境之一, 为人类社会提供了许多具有巨大价值的服务和利益[4], 莱州湾有黄河和小清河等低盐水体注入, 其河口附近的环境受到自然环境和人类的双重影响[5-8]。由于淡水和海水混合的地球化学过程, 河口处往往会截留住河流所携带的沉积物和有机物质, 但这些沉积物和有机物质也可能包括各种污染物, 且大多数河口都靠近人类活动区域, 易受人为干扰。以往的研究主要集中在对莱州湾小清河口近岸海域大型底栖动物的季节调查, 其中很容易忽略一些关键因素, 如河口附近盐度的剧烈变化对大型底栖动物分布的影响。本研究于2019年7月、8月和9月对莱州湾小清河口近岸海域大型底栖动物群落结构、物种多样性进行调查与分析, 并利用底栖生物评价指标——丰度/生物量比较曲线法、生物多样性指数、AZTI海洋生物指示(AMBI)和多元-AMBI指数[9-10]对莱州湾小河口近岸海域底栖生态系统的健康状况进行评价, 以其为了解人类活动和气候变化对河口底栖生态系统的影响提供理论参考。
1 材料与方法 1.1 站位布设与样品采集研究区域位于莱州湾南部小清河入海口的岸海域, 设置4条大致垂直于海岸线分布的调查断面, 每个断面设置4个调查站位, 共16个站位(图 1), 断面Ⅰ:1—4号站位, 断面Ⅱ:5—8号站位, 断面Ⅲ:9—12号站位, 断面Ⅳ:13—16号站位, 其中断面Ⅲ和断面Ⅳ之间有一条拦沙大坝, 分别于2019年7月、8月和9月份进行三个航程的调查。使用抓斗采泥器(型号为Van Veen grab 1000cm2)采集2—3次合并为一个样品, 现场用0.5 mm的取样筛过滤筛选沉积物, 仔细挑选大型底栖动物样品, 用5%的甲醛溶液固定, 样品带回实验室, 更换固定液, 对样品进行鉴定分类、个体计数和湿质量称量(精确到0.0001g), 最后依据采样面积折算成丰度(个/m2)和生物量(g/m2)。样品的采集、处理、保存、计数、称重等均按照《海洋调查规范第6部分:海洋生物调查》(GB/T 12763.6—2007)的调查方法进行。

|
| 图 1 莱州湾小清河口近岸海域调查站位示意图 Fig. 1 The sketch map of stations near the Xiaoqing Estuary of the Laizhou Bay |
大型底栖动物群落特征分析主要通过优势度指数(Y)[11]、Shannon-Wiener多样性指数(H′)、Margalef丰富度指数(D)及Pielou′s均匀度指数(J′)[12]进行分析, 公式如下:
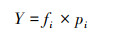
|
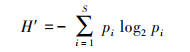
|
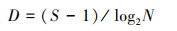
|
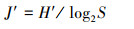
|
式中, S为总物种数;N为总个体数, pi=ni/N, pi为i种在总数量中的比例, ni为第i种个体数;fi为i种在采样点中出现频率, Y≥0.02时, 定为优势种。
1.2.2 群落结构分析利用PRIMER 5.0软件进行大型底栖动物群落结构分析, 对大型底栖动物的物种丰度进行log(x+1)转换后, 计算站位间的Bray-Curtis相似性系数, 构建相似性矩阵, 并采用等级聚类方法和非度量多维尺度分析大型底栖动物群落的空间分布, 通过相似性百分比分析不同物种对各群落组内相似性和组间差异性的平均贡献率[13]。
1.2.3 底栖生境健康评价底栖生境健康评价采用丰度/生物量比较曲线法、多样性指数法、AMBI指数和M-AMBI指数法。本研究数据处理中多样性指数评价参照蔡立哲等人划分的评价标准[14], AMBI指数根据各种底栖动物对环境的敏感程度, 将其分为5个生态组(Ecological Group, EG), EGⅠ即干扰敏感种(disturbance-sensitive species), 对富营养化非常敏感;EGⅡ即干扰不敏感种(disturbance-indifferent species), 对有机物过剩不敏感;EGⅢ即干扰耐受种(disturbance-tolerant species), 可以忍耐过量的有机物;EGⅣ即二阶机会种(the second-order opportunistic species), 生存在显著失衡的环境状态下;EGⅤ即一阶机会种(the first-order opportunistic species), 生存在显著失衡的环境下, 皆是食用沉积物的动物。M-AMBI指数结合欧盟对生态环境质量的定义, 将“差”生态质量的参照状态值选自与无生命沉积物下的群落结构参数和AMBI值, 其多样性指数和物种数参考值取0, AMBI的参照值取6[15-16];评价标准详见表 1。
| AMBI值 AMBI value |
扰动等级 Classification |
M-AMBI值 M-AMBI value |
底栖生境健康状况 The benthic habitat health |
| 0.0<AMBI≤1.2 | 无扰动 | 0.77<M-AMBI≤1 | 高等 |
| 1.2<AMBI≤3.3 | 轻度扰动 | 0.53<M-AMBI≤0.77 | 良好 |
| 3.3<AMBI≤5.0 | 中度扰动 | 0.38<M-AMBI≤0.53 | 一般 |
| 5.0<AMBI≤6.0 | 重度扰动 | 0.20<M-AMBI≤0.38 | 较差 |
| 6.0<AMBI≤7.0 | 极端扰动 | 0<M-AMBI≤0.20 | 差 |
| AMBI:AZTI海洋生物指数AZTI′s marine biotic index; M-AMBI: 多变量海洋生物指数Multivariate-AZTI′s marine biotic index | |||
共获得大型底栖动物64种, 其中多毛类24种, 占37.5%, 甲壳类19种, 占30.0%, 软体动物17种, 占26.6%, 其它动物4种, 占6.0%。本研究中3个航次的优势种及其所属类群和生态分组的情况如表 2所示, 根据优势度大于0.02的物种为优势种, 可以看出主要的优势种是软体动物的光滑河蓝蛤(Potamocorbula laevis)、彩虹明樱蛤(Moerella iridescens)、半褶织纹螺(Nassarius semiplicatus)、四角蛤蜊(Mactra veneriformis), 多毛类的寡鳃齿吻沙蚕(Nephthys oligobranchia)、中蚓虫(Mediomastus californiensis), 甲壳类的尖齿拳蟹(Philyra acutidens)、东方长眼虾(Ogyrides orientalis)、短角双眼钩虾(Ampelisca brevicornis)。各个航次的优势种不同, 其中彩虹明樱蛤和寡鳃齿吻沙蚕在三个航次的调查中均为优势物种。三个航次的优势种均为多毛类、软体动物和甲壳动物, 其生态分组优势种以EGⅠ、EGⅡ为主。
| 航次(年-月) Voyage |
类群 Group |
种名 Species |
生态分组 Ecological group |
优势度(Y) Dominance |
| 2019-07 | 软体动物 | 光滑河蓝蛤Potamocorbula laevis | EGⅤ | 0.11 |
| 软体动物 | 彩虹明樱蛤Moerella iridescens | EGⅠ | 0.08 | |
| 多毛类 | 寡鳃齿吻沙蚕Nephthys oligobranchia | EGⅡ | 0.03 | |
| 2019-08 | 软体动物 | 彩虹明樱蛤Moerella iridescens | EGⅠ | 0.11 |
| 多毛类 | 寡鳃齿吻沙蚕Nephthys oligobranchia | EGⅡ | 0.06 | |
| 甲壳类 | 尖齿拳蟹Philyra acutidens | NA | 0.02 | |
| 甲壳类 | 东方长眼虾Ogyrides orientalis | EGⅠ | 0.03 | |
| 软体动物 | 托氏鲳螺Umbonium thomasi | EGⅡ | 0.02 | |
| 2019-09 | 软体动物 | 彩虹明樱蛤Moerella iridescens | EGⅠ | 0.08 |
| 软体动物 | 半褶织纹螺Nassarius semiplicatus | EGⅡ | 0.07 | |
| 多毛类 | 中蚓虫Mediomastus californiensis | EGⅢ | 0.05 | |
| 多毛类 | 寡鳃齿吻沙蚕Nephthys oligobranchia | EGⅡ | 0.03 | |
| 软体动物 | 四角蛤蜊Mactra veneriformis | EGⅠ | 0.04 | |
| 甲壳类 | 短角双眼钩虾Ampelisca brevicornis | NA | 0.02 | |
| EGⅠ: 干扰敏感种Disturbance-sensitive species; EGⅡ: 干扰不敏感种Disturbance-indifferent species; EGⅢ: 干扰耐受种Disturbance-tolerant species; EGⅣ: 二阶机会种The second-order opportunistic species; EGⅤ: 一阶机会种The first-order opportunistic species; NA: 未分组Not assigned | ||||
根据2019年的3个航次16个取样站位的大型底栖动物数据显示, 丰度最高的是7月份的16号站位, 1710个/m2, 其丰度高的原因是采集到了大量的光滑河蓝蛤, 丰度最低的是7月份的3号和11号站位, 仅为45个/m2。大型底栖动物生物量最高的是9月份的8号站位, 611.8g/m2, 该站位采集到了大量的四角蛤蜊。生物量最低的9月份的15号站位, 仅为0.4g/m2。丰度和生物量的空间分布如图 2, 7月份丰度高的站位主要集中在靠近小清河口近岸海域北部的16号附近;8月份小清河口北部丰度减小, 南部丰度增加, 主要集中在1、2号站位附近;9月份丰度空间分布情况与8月份相似, 丰度值较8月份有所增加;7、8、9月份生物量的分布情况大体相同, 主要位于靠近小清河河口近岸海域附近, 并随着远离河口, 生物量呈现出下降的趋势。

|
| 图 2 不同月份大型底栖动物丰度和生物量空间分布 Fig. 2 The spatial distributions of species biomass and abundance of macrobenthos in different months |
根据大型底栖动物丰度和站位间聚类结果, 按照Bray-Curtis相似性系数10%、18%和19%, 对三个航次16个站位的大型底栖动物群落结构进行分组和分析。相似性检验表明, 三次调查的不同聚类组间均存在显著性差异(7月, R=0.509, P<0.01;8月, R=0.627, P<0.01;9月, R=0.662, P<0.01)。
2019年7月大型底栖动物群落分为3组, Ⅰ组7个站位(1、2、7、8、9、10和13), 组内平均相似性为30.4%, 主要贡献种为寡节甘吻沙蚕、彩虹明樱蛤、寡鳃齿吻沙蚕、四角蛤蜊、菲律宾蛤仔, 贡献率依次为26.3%、19.5%、11.5%、9.9%和9.6%;Ⅱ组包括站位15和16组成, 组内平均相似性为36.9%, 主要贡献种为光滑河篮蛤、半褶织纹螺, 贡献率为85.9%和14.1%;Ⅲ组7个站位(3、4、5、6、11、12和14), 组内平均相似性为34.3%, 主要贡献种为彩虹明樱蛤、索沙蚕、棘刺锚参、寡鳃齿吻沙蚕、尖锥虫、短角双眼钩虾, 贡献率依次为23.4%、14.6%、14.3%、10.0%、9.9%和9.3%。
8月研究区域的大型底栖动物群落可分为4组, Ⅰ组包括6个站位(2、4、11、12、14和15), 组内平均相似性为42.7%, 主要贡献种为彩虹明樱蛤、尖锥虫、寡鳃齿吻沙蚕和中蚓虫, 贡献率依次为27.5%、25.0%、16.5%和9.6%。Ⅱ组由站位5、9和16号站位组成, 组内平均相似性为27.7%, 主要贡献种为尖齿拳蟹、托氏鲳螺、彩虹明樱蛤、半褶织纹螺, 贡献率依次为35.6%、28.0%、15.7%和11.4%。Ⅲ组包括6个站位(1、3、6、7、8和10), 组内平均相似性为38.0%, 主要贡献种为寡鳃齿吻沙蚕、东方长眼虾、彩虹明樱蛤、四角蛤蜊、尖齿拳蟹, 贡献率依次为31.8%、19.4%、13.4%、12.0%、10.5%;13号站位为单独一组。
9月研究区域的大型底栖动物群落可分为3组, Ⅰ组由站位1、8、9、10和16, 5个站位组成, 组内平均相似性为42.1%, 主要贡献种为四角蛤蜊、半褶织纹螺、托氏鲳螺、尖齿拳蟹、彩虹明樱蛤, 贡献率依次为34.1%、12.6%、12.0%、11.1%、9.7%。Ⅱ组包括站位2、4、6、7和15, 组内平均相似性为45.7%, 主要贡献种为彩虹明樱蛤、寡鳃齿吻沙蚕、中蚓虫、短角双眼钩虾, 贡献率依次为31.6%、19.9%、11.4%和9.7%。Ⅲ组包括站位3、5、11、12、13和14, 组内平均相似性为44.1%, 主要贡献种为纽虫、中蚓虫、扁玉螺、短角双眼钩虾和尖锥虫, 贡献率依次为21.2%、16.0%、15.3%、13.3%和12.6%。

|
| 图 3 不同月份大型底栖动物等级聚类和非度量多维度排序分析图 Fig. 3 The CLUSTER picture and NMDS picture of macrobenthos in different months |
莱州湾小清河口近岸海域3个航次的大型底栖动物的物种Shannon-Wiener多样性指数(H′)、丰富度指数(D)和均匀度指数(J′)空间分布情况见图 4。三个航次的整体趋势为多样性指数上升, 丰富度下降, 均匀度上升, 但其差异不显著(P>0.05), 其空间分布情况为随着远离河口, 多样性指数和均匀度指数的值均逐渐增大, 丰富度指数变化不明显。其中H′最高出现在7月、8月的1号站位和9月的2号站位, 其位置均在远离小清河口的南部区域, 为2.2, 最低为7月份的16号站位, 为0.1。D最高为7月份和8月份的1号站位和9月份的6号站位, 为1.6, 最低为7月份的16号站位, 为0.2。J′最高为8月份的13、14号站位和9月份的5、6、13号站位, 为1.0, 最低为7月份的16号站位, 为0.1。

|
| 图 4 不同月份大型底栖动物生物多样性指数空间分布 Fig. 4 The spatial distribution of H′, J′ and D of macrobenthos in different months H′: Shannon-Wiener多样性指数; D: Margalef丰富度指数; J′: Pielou′s均匀度指数 |
大型底栖动物群落丰度-生物量ABC曲线如图 5所示。7月、8月和9月份大型底栖动物群落生物量优势度曲线均在丰度优势度曲线之上, 说明大型底栖动物群落未受干扰, 大型底栖动物生物量由大个体的保守种占据优势, 丰度较小, 而小个体机会种如多毛类则占据丰度上的优势, 种间丰度差异较生物量差异小, 显示底栖动物群落生态系统处于健康的状态。

|
| 图 5 不同月份大型底栖动物群落丰度/生物量比较曲线 Fig. 5 The abundance/biomass comparison of macrobenthic communities in different months |
对莱州湾小清河河口近岸海域的大型底栖动物进行AMBI和M-AMBI指数分析(见表 3), 7月份调查海域AMBI值的范围为0.5—5.9, 均值为1.4, 结果显示, 该航次有9个站位为无扰动状态, 6个站位处于轻度扰动, 只有16号站位为重度扰动, 其空间分布上受小清河口影响, 大多数处于轻度扰动的站位处于靠近小清河口的近岸海域;M-AMBI值的范围为0.1—0.9, 均值为0.7, M-AMBI生态环境状况评价结果显示, 该调查航次有6个站位处于高等状态, 8个站位处于好的状态, 1个站位处于一般, 1个站位处于差。8月份调查海域AMBI值的范围为0.2—2.6, 均值为1.1, 结果显示, 该航次有10个站位处于无扰动状态, 6个站位处于轻度扰动, 其空间分布规律不明显;M-AMBI值的范围为0.4—0.9, 均值为0.7, M-AMBI生态环境状况评价结果显示, 该调查航次有9个站位处于高等状态, 6个站位处于好, 1个站位处于一般状态。9月份调查海域AMBI值的范围为0.1—1.6, 均值为1.0, 结果显示, 有13个站位处于无扰动状态, 其余站位处于轻度扰动状态, 且处于轻度干扰状态的站位均位于靠近小清河口附近海域;M-AMBI值的范围为0.6—1.0, 均值为0.8, M-AMBI生态环境状况评价结果显示, 该调查海域有5个站位处于高等状态, 11个站位处于好的状态。
| 航次(年-月) Voyage |
站位 Station |
EGⅠ/% | EGⅡ/% | EGⅢ/% | EGⅣ/% | EGⅤ/% | NA/% | AMBI | 扰动等级 Disturbance classification |
M-AMBI | 生态环境状况 Ecological status |
| 2019-07 | 1 | 57.9 | 42.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.6 | 无扰动 | 0.8 | 高等 |
| 2 | 20.6 | 67.6 | 11.8 | 0.0 | 0.0 | 0.0 | 1.9 | 轻度扰动 | 0.9 | 良好 | |
| 3 | 11.1 | 88.9 | 0.0 | 0.0 | 0.0 | 0.0 | 1.3 | 轻度扰动 | 0.4 | 一般 | |
| 4 | 61.5 | 30.8 | 7.7 | 0.0 | 0.0 | 0.0 | 0.7 | 无扰动 | 0.9 | 良好 | |
| 5 | 52.9 | 35.3 | 11.8 | 0.0 | 0.0 | 0.0 | 0.9 | 无扰动 | 0.8 | 良好 | |
| 6 | 57.1 | 39.3 | 0.0 | 0.0 | 3.6 | 0.0 | 0.8 | 无扰动 | 0.9 | 良好 | |
| 7 | 25.0 | 45.0 | 30.0 | 0.0 | 0.0 | 0.0 | 1.6 | 轻度扰动 | 0.8 | 良好 | |
| 8 | 48.6 | 42.9 | 5.7 | 2.9 | 0.0 | 0.0 | 0.9 | 无扰动 | 0.8 | 良好 | |
| 9 | 3.2 | 88.7 | 8.1 | 0.0 | 0.0 | 0.0 | 1.6 | 轻度扰动 | 0.6 | 高等 | |
| 10 | 40.5 | 16.2 | 43.2 | 0.0 | 0.0 | 2.6 | 1.5 | 轻度扰动 | 0.9 | 良好 | |
| 11 | 55.6 | 33.3 | 11.1 | 0.0 | 0.0 | 0.0 | 0.8 | 无扰动 | 0.7 | 高等 | |
| 12 | 79.3 | 10.3 | 10.3 | 0.0 | 0.0 | 0.0 | 0.5 | 无扰动 | 0.7 | 高等 | |
| 13 | 66.7 | 33.3 | 0.0 | 0.0 | 0.0 | 0.0 | 0.5 | 无扰动 | 0.8 | 良好 | |
| 14 | 55.6 | 44.4 | 0.0 | 0.0 | 0.0 | 0.0 | 0.7 | 无扰动 | 0.7 | 高等 | |
| 15 | 31.4 | 20.0 | 2.9 | 0.0 | 45.7 | 0.0 | 3.1 | 轻度扰动 | 0.7 | 高等 | |
| 16 | 0.0 | 1.8 | 0.0 | 0.0 | 98.2 | 0.0 | 5.9 | 重度扰动 | 0.1 | 差 | |
| 2020-08 | 1 | 42.3 | 55.1 | 2.6 | 0.0 | 0.0 | 0.0 | 0.9 | 无扰动 | 0.9 | 良好 |
| 2 | 47.1 | 47.1 | 5.9 | 0.0 | 0.0 | 0.0 | 0.9 | 无扰动 | 0.9 | 良好 | |
| 3 | 28.6 | 71.4 | 0.0 | 0.0 | 0.0 | 0.0 | 1.1 | 无扰动 | 0.6 | 高等 | |
| 4 | 77.8 | 22.2 | 0.0 | 0.0 | 0.0 | 0.0 | 0.3 | 无扰动 | 0.8 | 良好 | |
| 5 | 10.3 | 5.1 | 84.6 | 0.0 | 0.0 | 0.0 | 2.6 | 轻度扰动 | 0.4 | 一般 | |
| 6 | 75.0 | 20.8 | 4.2 | 0.0 | 0.0 | 0.0 | 0.4 | 无扰动 | 0.9 | 良好 | |
| 7 | 61.5 | 38.5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.6 | 无扰动 | 0.8 | 高等 | |
| 8 | 7.1 | 50.0 | 42.9 | 0.0 | 0.0 | 0.0 | 2.0 | 轻度扰动 | 0.6 | 高等 | |
| 9 | 0.0 | 78.3 | 21.7 | 0.0 | 0.0 | 0.0 | 1.8 | 轻度扰动 | 0.6 | 高等 | |
| 10 | 64.3 | 35.7 | 0.0 | 0.0 | 0.0 | 0.0 | 0.5 | 无扰动 | 0.8 | 高等 | |
| 11 | 65.0 | 25.0 | 10.0 | 0.0 | 0.0 | 0.0 | 0.7 | 无扰动 | 0.8 | 良好 | |
| 12 | 92.9 | 0.0 | 7.1 | 0.0 | 0.0 | 0.0 | 0.2 | 无扰动 | 0.7 | 高等 | |
| 13 | 33.3 | 16.7 | 50.0 | 0.0 | 0.0 | 0.0 | 1.8 | 轻度扰动 | 0.6 | 高等 | |
| 14 | 50.0 | 14.3 | 21.4 | 7.1 | 7.1 | 0.0 | 1.6 | 轻度扰动 | 0.8 | 良好 | |
| 15 | 75.0 | 12.5 | 0.0 | 12.5 | 0.0 | 0.0 | 0.8 | 无扰动 | 0.7 | 高等 | |
| 16 | 21.1 | 57.9 | 0.0 | 0.0 | 21.1 | 0.0 | 2.1 | 轻度扰动 | 0.7 | 高等 | |
| 2021-09 | 1 | 34.4 | 62.5 | 3.1 | 0.0 | 0.0 | 0.0 | 1.0 | 无扰动 | 0.9 | 良好 |
| 2 | 16.4 | 61.2 | 22.4 | 0.0 | 0.0 | 0.0 | 1.6 | 轻度扰动 | 0.8 | 良好 | |
| 3 | 60.0 | 20.0 | 20.0 | 0.0 | 0.0 | 0.0 | 0.9 | 无扰动 | 0.8 | 良好 | |
| 4 | 81.3 | 15.6 | 0.0 | 3.1 | 0.0 | 0.0 | 0.4 | 无扰动 | 0.8 | 高等 | |
| 5 | 41.7 | 41.7 | 16.7 | 0.0 | 0.0 | 0.0 | 1.1 | 无扰动 | 0.8 | 良好 | |
| 6 | 37.5 | 50.0 | 12.5 | 0.0 | 0.0 | 0.0 | 1.1 | 无扰动 | 0.7 | 高等 | |
| 7 | 50.0 | 40.6 | 9.4 | 0.0 | 0.0 | 0.0 | 0.9 | 无扰动 | 1.0 | 良好 | |
| 8 | 39.6 | 50.0 | 4.2 | 0.0 | 6.3 | 0.0 | 1.3 | 轻度扰动 | 0.8 | 良好 | |
| 9 | 28.6 | 66.7 | 0.0 | 0.0 | 4.8 | 0.0 | 1.3 | 轻度扰动 | 0.7 | 高等 | |
| 10 | 92.9 | 7.1 | 0.0 | 0.0 | 0.0 | 0.0 | 0.1 | 无扰动 | 0.6 | 高等 | |
| 11 | 57.7 | 34.6 | 7.7 | 0.0 | 0.0 | 0.0 | 0.8 | 无扰动 | 0.9 | 良好 | |
| 12 | 60.0 | 33.3 | 6.7 | 0.0 | 0.0 | 0.0 | 0.7 | 无扰动 | 0.9 | 良好 | |
| 13 | 75.0 | 8.3 | 8.3 | 8.3 | 0.0 | 0.0 | 0.8 | 无扰动 | 0.9 | 良好 | |
| 14 | 52.6 | 15.8 | 31.6 | 0.0 | 0.0 | 0.0 | 1.2 | 无扰动 | 0.8 | 良好 | |
| 15 | 44.4 | 44.4 | 11.1 | 0.0 | 0.0 | 0.0 | 1.0 | 无扰动 | 0.7 | 高等 | |
| 16 | 35.7 | 50.0 | 14.3 | 0.0 | 0.0 | 6.7 | 1.2 | 无扰动 | 0.8 | 良好 |
根据本研究结果, 2019年莱州湾近岸海域三个航次共发现大型底栖动物64种, 7月份物种数最多, 为45种, 9月份最少, 为32种, 三个航次所获物种各类群所占的比例与林颂报道的结果一致, 为多毛类环节动物>软体动物>甲壳动物>其他类群[18]。有研究表明, 盐度是影响河口区大型底栖动物物种分布的重要环境因子, 并且随着盐度的降低, 大型底栖动物丰度会下降[19], 本研究的7、8、9月份是小清河径流量变化较大的时期, 其中8月份出现了小清河径流量增加, 导致调查海域盐度急剧下降, 从生物量和丰度的空间分布可以看出, 盐度的降低, 导致小清河河口附近海域大型底栖动物丰度和生物量下降, 物种数在三个调查航次中也有明显的下降。生物群落中优势种的优势度与群落稳定性密切相关, 一般来说, 生物群落的物种优势度越高, 说明群落内各物种的生态地位越不平衡, 生物群落也就更加脆弱;反之, 物种优势度越低, 则群落内各物种的生态地位更加趋于平衡, 生物群落就越稳定[20]。三个航次的调查均出现了高优势度的物种, 优势物种的数量也有所增加, 由此推测莱州湾小清河河口近海区域大型底栖动物群落的不稳定性。彩虹明樱蛤在三个航次调查中均为优势物种, 因其分布在淡水和海水之间, 能够忍受一定范围的盐度变化, 繁殖和发育的适宜盐度为7—42之间[21], 小清河口近岸海域的盐度范围适宜其繁殖与生长, 使其在调查中始终位于优势地位。
3.2 莱州湾小清河口近岸海域大型底栖动物多样性及群落结构特征三个航次的调查中大型底栖动物Shannon-Wiener多样性指数、丰富度指数、均匀度指数均显示靠近河口的多样性比近岸海域低, 生物多样性自河口向外海逐渐升高, 并在盐度较为稳定的区域出现丰度和生物量升高的现象。其原因可能与该区域相对于河口内盐度的变化幅度相对较窄有关, 既能维持河口生物的生存, 也能满足近岸生物的生存[22]。根据大型底栖动物群落结构聚类和非度量多维尺度分析结果表明, 调查海域大型底栖动物群落的相似性较低。7月份群落的空间分布以小清河河口为中心, 靠近河口的近海区域划分为组Ⅰ、组Ⅱ, 远离河口的海域划分为组Ⅲ。8月份由于小清河径流量增加, 群落的空间分布发生改变, 群落空间分布呈现斑块化。9月份其群落空间分布情况与7月份相似。水环境和沉积环境影响着河口近岸海域大型底栖动物群落的组成和分布, 造成各调查站位大型底栖动物群落低相似性的原因可能与河口近岸海域不稳定的环境因素有关, 特别是河流径流量的变化对河口近岸海域环境的影响。孙凯静等对黄河口及邻近海域底栖群落研究发现, 水深、盐度和溶解无机氮等是限制底栖生境健康的主要因子, 盐度较低的河口区不适于底栖动物的栖息[23]。周细平等对珠江河口区大型底栖群落结构研究发现, 盐度和pH是影响群落多样性分布的主要环境因子[24]。莱州湾小清河口近岸海域不稳定的环境因素造成河口近岸海域的复杂性, 形成了不同的小生境, 不同的小生境栖息着不同结构的底栖动物群落, 导致各个站位的底栖动物群落结构相似性都较低。
3.3 莱州湾小清河口近岸海域底栖生境健康状况丰度/生物量比较曲线、多样性指数H′、AMBI指数及M-AMBI指数法均能在一定程度上反映底栖动物受扰动状态, 近年来, 已被用于港湾、近岸海域及河口区的底栖生态环境质量评价[25-26], 其中AMBI指数及M-AMBI指数法是欧盟水框架指令评价方法之一, 它们被广泛应用于可口和海岸带水域的水环境质量检测和生态研究中[2]。这些生物指数的应用, 提升了人们对海洋底栖无脊椎动物群落的生态功能的认知, 使得底栖生物群落水平上的环境指示作用更加迅捷有效。2008—2013年, 林颂运用M-AMBI指数对小清河近岸海域底栖生境进行评价, 其健康状况为中等[18], 而本次的调查中, 近岸海域均处于高等或者良好的状态, 底栖生境的健康状况要优于以前。其原因与小清河水质状况有所改善有关, 范新凤对近十年小清河口入海口十年水质变化研究表明, 小清河入海口水质呈现好转状态, 污染状况大大减轻, 污水处理率的提高和水环境保护政策的实施是小清河水质明显改善的重要原因[27]。
由于海洋生态系统本身的复杂性以及人为环境压力的影响, 不同的评价指数在同一区域通常给出不同的结果[28]。在本研究中, 不同生物指数的评价结果存在一定的差异, 由表 5可以看出, 多样性指数H′研究结果表明, 莱州湾小清河口近岸海域的底栖生境状况处于一般和差之间, 而M-AMBI指数的评价结果为大部分站位处于高等和良好状态。二者在莱州湾小清河河口近海区域评价结果有较大的差异, 多样性指数H′比M-AMBI指数评价结果低两个等级, 与林颂等[18]对小清河口近岸海域调查结果一致。其原因可能是由于河口环境因子的变化会导致河口近岸海域较低的生物多样性, 因为很少有生物能够适应和忍受生存条件的快速变化[29], 特别是盐度是河口区生物分布的主要限制因子[30]。同时ABC曲线表明莱州湾小清河口近岸海域底栖群落处于未干扰状态, 与M-AMBI指数分析结果相似。
由此看来, 多样性指数评价并不能很好的反应小清河口近岸海域的底栖生境健康状况, 评价结果与实际情况不符合, 因此该方法具有一定的局限性[31]。M-AMBI指数为综合了AMBI、生物多样性和物种丰度的复合指数, 其评价结果综合多种因素, 包括底栖动物自身的敏感程度、周边生态环境变化情况及人类对环境造成的压力情况等, 因此其结果更能真实反映该区域的底栖生境质量状况[32], 通过海洋生物指数M-AMBI评价可以看出莱州湾小清河河口近海区域底栖生境处于高等或者良好的状态, 底栖群落处于未干扰或者轻度干扰的状态。
| 航次(年-月) Voyage |
站位 Station |
底栖生境健康 Benthic habitat health |
航次(年-月) Voyage |
站位 Station |
底栖生境健康 Benthic habitat health |
航次(年-月) Voyage |
站位 Station |
底栖生境健康 Benthic habitat health |
|||||
| H′ | M-AMBI | H′ | M-AMBI | H′ | M-AMBI | ||||||||
| 2019-07 | 1 | 一般 | 高等 | 2019-08 | 1 | 一般 | 差 | 2019-09 | 1 | 一般 | 良好 | ||
| 2 | 良好 | 良好 | 2 | 一般 | 良好 | 2 | 差 | 良好 | |||||
| 3 | 较差 | 一般 | 3 | 差 | 良好 | 3 | 差 | 良好 | |||||
| 4 | 良好 | 良好 | 4 | 差 | 高等 | 4 | 差 | 高等 | |||||
| 5 | 一般 | 良好 | 5 | 极差 | 良好 | 5 | 一般 | 良好 | |||||
| 6 | 良好 | 良好 | 6 | 差 | 一般 | 6 | 差 | 高等 | |||||
| 7 | 良好 | 良好 | 7 | 差 | 良好 | 7 | 一般 | 良好 | |||||
| 8 | 一般 | 良好 | 8 | 差 | 高等 | 8 | 差 | 良好 | |||||
| 9 | 较差 | 高等 | 9 | 差 | 高等 | 9 | 差 | 高等 | |||||
| 10 | 良好 | 良好 | 10 | 差 | 高等 | 10 | 较差 | 高等 | |||||
| 11 | 一般 | 高等 | 11 | 差 | 高等 | 11 | 一般 | 良好 | |||||
| 12 | 一般 | 高等 | 12 | 差 | 良好 | 12 | 一般 | 良好 | |||||
| 13 | 一般 | 良好 | 13 | 差 | 高等 | 13 | 一般 | 良好 | |||||
| 14 | 一般 | 高等 | 14 | 一般 | 高等 | 14 | 差 | 良好 | |||||
| 15 | 一般 | 高等 | 15 | 差 | 良好 | 15 | 差 | 高等 | |||||
| 16 | 较差 | 差 | 16 | 差 | 高等 | 16 | 差 | 良好 | |||||
| H′: Shannon-Wiener多样性指数 | |||||||||||||
| [1] |
国家海洋局海洋环境保护司. HY/T 087-2005近岸海洋生态健康评价指南. 北京: 中国标准出版社, 2005. |
| [2] |
Borja A, Franco J, Pérez V. A marine biotic index to establish the ecological quality of soft-bottom benthos within European estuarine and coastal environments. Marine Pollution Bulletin, 2000, 40(12): 1100-1114. DOI:10.1016/S0025-326X(00)00061-8 |
| [3] |
Borja A, Tunberg B G. Assessing benthic health in stressed subtropical estuaries, eastern Florida, USA using AMBI and M-AMBI. Ecological Indicators, 2011, 11(2): 295-303. DOI:10.1016/j.ecolind.2010.05.007 |
| [4] |
Chevillot X, Tecchio S, Chaalali A, Lassalle G, Selleslagh J, Castelnaud G, David V, Bachelet G, Niquil N, Sautour B, Lobry J. Global changes jeopardize the trophic carrying capacity and functioning of estuarine ecosystems. Ecosystems, 2019, 22(3): 473-495. DOI:10.1007/s10021-018-0282-9 |
| [5] |
郭富, 王保栋, 辛明, 张文全, 厉丞烜, 谢琳萍, 孙霞. 2015年春、夏季莱州湾营养盐分布特征. 海洋科学进展, 2017, 35(2): 258-266. DOI:10.3969/j.issn.1671-6647.2017.02.010 |
| [6] |
任彬彬, 袁伟, 孙坚强, 陈瑞盛, 王俊. 莱州湾金城海域鱼礁投放后大型底栖动物群落变化. 应用生态学报, 2015, 26(6): 1863-1870. |
| [7] |
刘晓收, 赵瑞, 华尔, 路璐, 张志南. 莱州湾夏季大型底栖动物群落结构特征及其与历史资料的比较. 海洋通报, 2014, 33(3): 283-292. |
| [8] |
李少文, 刘元进, 李凡, 张莹, 徐宗法, 吕振波, 王田田, 张爱波. 莱州湾大型底栖动物功能群现状. 生态学杂志, 2013, 32(2): 380-388. |
| [9] |
Muxika I, Borja Á, Bald J. Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status, according to the European Water Framework Directive. Marine Pollution Bulletin, 2007, 55(1/6): 16-29. |
| [10] |
Ranasinghe J A, Weisberg S B, Smith R W, Montagne D E, Thompson B, Oakden J M, Huff D D, Cadien D B, Velarde R G, Ritter K J. Calibration and evaluation of five indicators of benthic community condition in two California bay and estuary habitats. Marine Pollution Bulletin, 2009, 59(1/3): 5-13. |
| [11] |
国家海洋局. GB/T 12763.6-2007海洋调查规范第6部分: 海洋生物调查. 北京: 中国标准出版, 2008.
|
| [12] |
唐得昊, 邹欣庆, 刘兴健. 海岸带生态系统健康评价中能质和生物多样性的差异——以江苏海岸带为例. 生态学报, 2012, 33(4): 1240-1250. |
| [13] |
周红, 张志南. 大型多元统计软件PRIMER的方法原理及其在底栖群落生态学中的应用. 青岛海洋大学学报, 2003, 33(1): 58-64. DOI:10.3969/j.issn.1672-5174.2003.01.023 |
| [14] |
蔡立哲, 马丽, 高阳, 郑天凌, 林鹏. 海洋底栖动物多样性指数污染程度评价标准的分析. 厦门大学学报: 自然科学版, 2002, 41(5): 641-646. DOI:10.3321/j.issn:0438-0479.2002.05.023 |
| [15] |
Muxika I, Borja Á, Bonne W. The suitability of the marine biotic index (AMBI) to new impact sources along European coasts. Ecological Indicators, 2005, 5(1): 19-31. DOI:10.1016/j.ecolind.2004.08.004 |
| [16] |
Forchino A, Borja A, Brambilla F, Rodríguez J G, Muxika I, Terova G, Saroglia M. Evaluating the influence of off-shore cage aquaculture on the benthic ecosystem in Alghero Bay (Sardinia, Italy) using AMBI and M-AMBI. Ecological Indicators, 2011, 11(5): 1112-1122. DOI:10.1016/j.ecolind.2010.12.011 |
| [17] |
Borja A, Josefson A B, Miles A, Muxika I, Olsgard F, Phillips G, Rodríguez J G, Rygg B. An approach to the intercalibration of benthic ecological status assessment in the North Atlantic ecoregion, according to the European Water Framework Directive. Marine Pollution Bulletin, 2007, 55(1/6): 42-52. |
| [18] |
林颂. 小清河口及邻近海域沉积环境特征及底栖生境质量评价[D]. 青岛: 中国海洋大学, 2015.
|
| [19] |
Ysebaert T, Herman P M J, Meire P, Craeymeersch J, Verbeek H, Heip C H R. Large-scale spatial patterns in estuaries: estuarine macrobenthic communities in the Schelde estuary, NW Europe. Estuarine, Coastal and Shelf Science, 2003, 57(1/2): 335-355. |
| [20] |
刘录三, 孟伟, 郑丙辉, 李新正, 雷坤, 李中宇. 辽东湾北部海域大型底栖动物研究: Ⅰ. 种类组成与数量分布. 环境科学研究, 2008, 21(6): 118-123. |
| [21] |
顾晓英, 尤仲杰, 王一农. 几种环境因子对彩虹明樱蛤Moerella iridescens不同发育阶段的影响. 东海海洋, 1998, 16(3): 41-47. |
| [22] |
张敬怀. 珠江口及邻近海域大型底栖动物多样性随盐度、水深的变化趋势. 生物多样性, 2014, 22(3): 302-310. |
| [23] |
孙凯静, 罗先香, 张龙军, 林颂, 王潇. 黄河口及邻近海域底栖群落健康及生境适宜性评价. 中国海洋大学学报, 2015, 45(12): 107-112. |
| [24] |
周细平, 李贞, 吴培芳, 吴茜, 陈逸欣, 刘康格, 刘东艳, 王玉珏, 王跃启. 珠江河口区大型底栖动物群落结构. 生物多样性, 2019, 27(10): 1112-1121. DOI:10.17520/biods.2018321 |
| [25] |
魏永杰, 杨耀芳, 费岳军, 张海波, 叶然. 象山港大型底栖生态状况的多指数互校法评价与分析. 海洋环境科学, 2015, 34(5): 769-776. |
| [26] |
蔡文倩, 孟伟, 刘录三, 朱延忠, 周娟. 长江口海域底栖生态环境质量评价——AMBI和M-AMBI法. 环境科学, 2013, 34(5): 1725-1734. |
| [27] |
范新凤, 韩美, 王磊, 李秀萍, 周璟. 小清河入海口近十年水质变化及驱动因素分析. 环境科学, 2020, 41(4): 1619-1628. |
| [28] |
吴海燕, 陈克亮, 张平, 傅世锋, 侯建平, 陈庆辉. 基于不同生物指数的罗源湾生态环境质量状况评价. 应用生态学报, 2013, 24(3): 825-831. |
| [29] |
Miller J M, Crowder L B, Moser M L. Migration and utilization of estuarine nurseries by juvenile fishes: an evolutionary perspective//University of Texas Marine Science Institute, ed. Contributions in Marine Science. Texas: Marine Science Institute, The University of Texas at Austin, 1985, 27: 338-352. |
| [30] |
Laprise R, Dodson J J. Nature of environmental variability experienced by benthic and pelagic animals in the St. Lawrence Estuary, Canada. Marine Ecology Progress Series, 1993, 94(2): 129-139. |
| [31] |
Warwick R M, Clarke K R. Comparing the severity of disturbance: a meta-analysis of marine macrobenthic community data. Marine Ecology Progress Series, 1993, 92(3): 221-231. |
| [32] |
梁淼, 姜倩, 李德鹏, 柳圭泽, 陈兆林, 崔雷, 于大涛. 曹妃甸近岸海域大型底栖动物群落特征. 水产科学, 2019, 38(4): 479-491. |