 2021, Vol. 41
2021, Vol. 41文章信息
- 张丙行, 朱韩, 江姗, 黄克强, 潘玲, 席贻龙
- ZHANG Bingxing, ZHU Han, JIANG Shan, HUANG Keqiang, PAN Ling, XI Yilong
- 青霉素钠对萼花臂尾轮虫适合度的促进作用及其与藻密度的关系
- Promoting effects of sublethal concentrations of penicillin sodium on survival, reproduction and population growth of Brachionus calyciflorus in relation to Scenedesmus obliquus density
- 生态学报. 2021, 41(11): 4418-4427
- Acta Ecologica Sinica. 2021, 41(11): 4418-4427
- http://dx.doi.org/10.5846/stxb201907251568
-
文章历史
- 收稿日期: 2019-07-25
- 网络出版日期: 2021-04-06





抗生素的发现被认为是20世纪最重要的科学进步之一。60余年来, 抗生素被广泛应用于人和动物疾病的防治, 并作为生长促进剂被应用于畜牧业和水产养殖业[1]。抗生素在造福人类的同时, 也对环境造成了污染[1-5]。抗生素在杀灭靶向生物病原菌的同时, 也造成了部分病原菌抗性的形成[6], 并对部分非靶向生物产生毒性作用[1, 7-9]。
轮虫(Rotifer)是各类水体中广泛分布的一类浮游动物, 是抗生素作用的非靶向生物。由于其世代时间短、个体小、易于培养、繁殖速度快、对常见的水体污染物较敏感, 在水生态系统的物质循环和能量流动过程中具有重要的作用, 自上个世纪70年代起就被广泛应用于水生态毒理学研究[10-11]。迄今为止, 以轮虫为受试生物研究抗生素的慢性毒性作用已有一些报道。就轮虫种群增长率而言, 抗生素的毒性作用总体上呈现出以下三种格局:(1)12.5 mg/L的氯霉素、250和500 mg/L的盐酸氟哌酸均显著促进褶皱臂尾轮虫(B. plicatilis)的种群增长, 而浓度高于50 mg/L的氯霉素和750 mg/L的盐酸氟哌酸均起显著的抑制作用[12]。6.25 mg/L的氯霉素和红霉素[13]、180—200 μg/L的氟西汀(Fluoxetine)[14]和30—90 mg/L的盐酸土霉素[15]等均显著促进萼花臂尾轮虫的种群增长, 而浓度高于12.5 mg/L的氯霉素和红霉素[13]、浓度高于230 μg/L的氟西汀(Fluoxetine)[14]和180 mg/L的盐酸土霉素[15]等均起显著的抑制作用。(2)在2.0—10.0 mg/L的测试浓度范围内, 利福平(Rifampicin)显著地促进萼花臂尾轮虫的种群增长[16]。(3)在5.6—2000 mg/L的测试浓度范围内, 仅2000 mg/L的硫酸链霉素(streptomycin sulfate)、盐酸四环素(tetracycline hydrochloride)和酒石酸泰乐霉素(tylosin tartrate)显著抑制萼花臂尾轮虫和褶皱臂尾轮虫的种群增长[17];在50—200 μg/L的测试浓度范围内, 各浓度的阿莫西林(amoxicillin)显著抑制萼花臂尾轮虫和哈瓦那臂尾轮虫(B. havanaensis)的种群增长[18]。不同作用格局的呈现, 究竟是由于测试的抗生素浓度范围造成的, 还是由抗生素的种类决定的?
作为水生态系统中重要的初级生产者, 藻类在水体中可加速污染物降解, 降低其对浮游动物的毒性;升高的藻类密度可提高轮虫等浮游动物对污染物的耐受能力[19]。因此, 有关污染物浓度和食物密度对轮虫存活、生殖和种群增长的交互作用研究持续受到国内外学者的关注[20-26]。然而, 近年来, 已有的研究所揭示出的有关抗生素对轮虫存活、生殖和种群增长的影响与食物密度之间的关系并不一致[15-16, 26]。如亚致死浓度的利福平对萼花臂尾轮虫生命期望、净生殖率和种群内禀增长率的促进作用幅度随着藻密度由1.0×106个细胞/mL升高至4.0×106个细胞/mL而逐渐显著地增大[16], 但亚致死浓度的盐酸土霉素对萼花臂尾轮虫生命期望、净生殖率和种群内禀增长率的促进作用幅度在2.0×106个细胞/mL的斜生栅藻密度下显著高于1.0×106和4.0×106个细胞/mL的斜生栅藻密度下[15]。其他亚致死浓度的抗生素对轮虫存活、生殖和种群增长的影响与食物密度之间的关系如何仍需研究。
青霉素钠(penicillin sodium salt), 又名苄基青霉素钠盐、青霉素G钠盐、苄青霉素钠、6-苯乙酰胺基青霉烷酸, 是由青霉素G(苄青霉素)和钠结合形成的盐。它对革兰氏阳性细菌等致病微生物有较好的杀菌效果, 具有抗菌作用强、疗效高及毒性低等优点, 故至今仍被广泛应用。然而, 作为一种新兴污染物[1-3], 其对轮虫的慢性毒性作用格局是什么?慢性毒性作用的强度与食物密度之间的关系如何?所有这些尚无从知晓。为此, 本文以萼花臂尾轮虫为受试生物, 采用生命表试验方法, 研究了不同浓度的青霉素钠在3个食物密度下对萼花臂尾轮虫生命表统计学参数的影响, 旨在揭示青霉素类抗生素对轮虫实验种群动态的影响, 并为青霉素类污染物的生物监测提供适宜的指标。
1 材料与方法 1.1 轮虫的来源和预培养受试生物萼花臂尾轮虫采于芜湖市镜湖, 在实验室内进行克隆培养。所用培养液为EPA, 由96mg NaHCO3、60mg CaSO4、60mg MgSO4和4mg KCl溶解于1L蒸馏水中配制而成[27];所用饵料为经HB-4培养基[28]培养的、处于指数增长期的斜生栅藻, 离心浓缩并经显微计数后使用。实验前, 将轮虫置于(25±1)℃、自然光照(光照强度约1300 lx, L∶D约12∶12)的恒温培养箱内进行为期1周的预培养。期间, 每天投喂密度分别为1.0×106、2.0×106和4.0×106个细胞/mL的斜生栅藻并更换轮虫培养液一次, 同时通过去除一部分个体使轮虫种群始终处于指数增长期。
1.2 测试液的配置实验所用的青霉素钠购于Sigma公司, 分析纯(纯度≥98%)。测试液通过母液稀释法配置。实验前用蒸馏水配置0.1g/L的原液, 再用蒸馏水稀释成10mg/L的母液, 放置于4℃冰箱中保存以供实验使用。实验时用EPA将其配置成所需浓度梯度的测试液。
1.3 慢性毒性试验鉴于水环境中青霉素钠的浓度仅为80—125 ng/L[29], 不会对轮虫产生显著的毒性效应[1, 4-5], 因此实验时参照阿莫西林影响萼花臂尾轮虫生命表统计学参数的LOEC值[18], 将青霉素钠浓度设置为200、400、600、800和1000 μg/L, 另设置1个空白对照组;斜生栅藻密度设置为1.0×106(较低)、2.0×106(中等)和4.0×106个细胞/mL(较高)。每个藻密度下、每个青霉素钠浓度组设置3个重复, 实验共计45个重复(5个青霉素钠浓度×3个食物密度×3个重复)。实验时, 分别从预培养一周后的各组轮虫中随机吸取龄长在4h内的轮虫幼体, 放入8 mL玻璃杯中作为受试母体, 并加入5 mL测试液(其中分别含有相应浓度的青霉素钠和相应密度的斜生栅藻), 每个玻璃杯中放置10只轮虫幼体。实验在(25±1)℃、自然光照的恒温培养箱中进行。实验过程中, 每间隔12 h观察1次并记录母体的存活数和孵化出的幼体数, 移去死亡母体, 并将新生的幼体移至另一玻璃杯中继续培养, 待其产卵后确定雌体类型, 用于计算轮虫一生所产全部后代中混交雌体所占的比例;每间隔24 h更换1次含有相应密度的斜生栅藻的测试液。实验进行至轮虫母体全部死亡为止。
1.4 研究参数的定义与计算方法特定年龄存活率(lx):X年龄组开始时存活个体所占的比例。
特定年龄繁殖率(mx):X年龄组平均每个个体所产的雌性后代数。
生命期望值(e0):每个个体出生时能活多久的估计值。
净生殖率(R0):种群经过一个世代后的净增长率, R0=∑lxmx。
世代时间(T):完成一个世代所需要的时间, T=∑lxmxx/R0。
种群内禀增长率(rm):种群在特定试验条件下的最大增长率;根据方程rm=lnR0/T在粗略计算的基础上, 再根据方程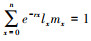
后代混交率:所有雌性后代中混交雌体所占的比例。
1.5 数据统计和分析采用SPSS 19.0分析软件对数据进行分析。对所得数据作正态性检验后, 对符合正态分布的各组数据通过单因素方差分析(one-way ANOVA)和多重比较(LSD检验)分析各青霉素钠浓度处理组与空白对照组间的差异显著性, 通过双因素方差分析检测青霉素钠浓度、斜生栅藻密度及其交互作用对轮虫各生命表统计学参数的影响;运用生存分析中的Log-Rank分析各藻密度下各青霉素钠浓度处理组轮虫存活率与对照组之间的差异显著性;对轮虫各生命表统计学参数与青霉素钠浓度之间的关系进行回归分析。
2 结果与分析 2.1 不同藻密度下青霉素钠浓度对轮虫存活率和繁殖率的影响各食物密度下, 对照组和5个青霉素钠浓度处理组中萼花臂尾轮虫的特定年龄存活率和特定年龄繁殖率如图 1所示。各藻密度下, 各青霉素钠浓度对萼花臂尾轮虫的存活率均无显著性影响(P > 0.05)。当藻密度为1.0×106个细胞/mL时, 轮虫的繁殖率峰值在200 μg/L的青霉素钠溶液中最高, 为5.27个后代(雌体)-1 (12h)-1;当藻密度为2.0×106个细胞/mL时, 轮虫的繁殖率峰值在800 μg/L的青霉素钠溶液中最高, 为4.67个后代(雌体)-1 (12h)-1;当藻密度为4.0×106个细胞/mL时, 轮虫的繁殖率峰值在1000 μg/L的青霉素钠溶液中最高, 为4.42个后代(雌体)-1 (12h)-1。

|
| 图 1 不同食物密度和青霉素钠浓度下萼花臂尾轮虫的存活率和繁殖率 Fig. 1 The age-specific survivorship (filled square) and fecundity curves (open square) of B. calyciflorus exposed to five concentrations of penicillin sodium and three S. obliquus densities |
当藻密度为1.0×106个细胞/mL时, 青霉素钠浓度对萼花臂尾轮虫的生命期望、世代时间和净生殖率均具有显著影响(P < 0.05)。与对照组相比, 400 μg/L青霉素钠处理组中轮虫的生命期望显著延长, 200和400 μg/L青霉素钠处理组中轮虫的世代时间显著延长(P < 0.05), 200—600和1000 μg/L青霉素钠处理组中轮虫的净生殖率显著提高(P < 0.05)(表 1)。
| 食物密度 Food density/ (个细胞/mL) |
青霉素浓度 Penicillin sodium salt concentration /(μg/L) |
生命期望 Lifeexpectancy at hatching /h |
世代时间 Generation time/h |
净生殖率 Net reproductive rate/ (后代(雌体)-1生-1) |
内禀增长率 Intrinsic rate of population increase/h |
后代混交率 Proportion of sexual offspring |
| 1.0×106 | 0 | 80.0±0.4 | 50.1±0.4 | 9.5±0.1 | 0.0506±0.0003 | 0.03±0.01 |
| 200 | 89.6±8.3 | 61.7±5.2* | 14.2±1.9* | 0.0551±0.0006 | 0.04±0.02 | |
| 400 | 96.8±0.8* | 66.7±2.0** | 17.2±1.0** | 0.0514±0.0006 | 0.03±0.01 | |
| 600 | 86.0±2.8 | 59.6±4.1 | 13.3±1.0* | 0.0518±0.0017 | 0.03±0.01 | |
| 800 | 75.2±3.6 | 55.0±2.9 | 11.6±0.4 | 0.0515±0.0006 | 0.02±0.02 | |
| 1000 | 81.6±4.2 | 55.9±1.3 | 13.4±1.3* | 0.0560±0.0024 | 0.02±0.01 | |
| 2.0×106 | 0 | 60.0±3.0 | 42.6±1.9 | 4.6±0.7 | 0.0364±0.0036 | 0.00±0.00 |
| 200 | 87.6±13.4* | 61.9±5.0** | 14.0±2.9** | 0.0496±0.0030** | 0.02±0.01 | |
| 400 | 84.4±1.1* | 57.1±0.9** | 16.7±1.4** | 0.0577±0.0008** | 0.03±0.0 | |
| 600 | 101.6±6.2** | 64.0±2.6** | 21.0±1.3** | 0.0589±0.0009** | 0.02±0.01 | |
| 800 | 102.4±7.7** | 63.0±3.5** | 21.1±2.0** | 0.0590±0.0011** | 0.02±0.01 | |
| 1000 | 99.6±4.3** | 62.1±1.6** | 20.8±1.6** | 0.0589±0.0013** | 0.02±0.00 | |
| 4.0×106 | 0 | 84.4±8.2 | 56.6±7.9 | 7.4±1.2 | 0.0382±0.0014 | 0.02±0.01 |
| 200 | 82.8±11.1 | 59.0±9.6 | 11.2±2.4 | 0.0490±0.0016 | 0.00±0.00 | |
| 400 | 75.6±1.8 | 51.0±4.8 | 11.0±1.4 | 0.0572±0.0063 | 0.00±0.00 | |
| 600 | 74.0±5.1 | 53.3±2.5 | 10.3±0.6 | 0.0488±0.0025 | 0.01±0.02 | |
| 800 | 79.6±5.2 | 56.331±4.6 | 10.8±1.8 | 0.0476±0.0012 | 0.00±0.00 | |
| 1000 | 63.2±2.9 | 50.1±2.2 | 9.2±1.1 | 0.0476±0.0029 | 0.00±0.00 | |
| */** 与同一食物密度下的对照组相比有显著性差异(* P < 0.05, ** P < 0.01) | ||||||
当藻密度为2.0×106个细胞/mL时, 青霉素钠浓度对轮虫的生命期望、世代时间、净生殖率和内禀增长率均具有显著影响(P < 0.05)。与对照组相比, 200—1000 μg/L青霉素钠处理组中轮虫的生命期望和世代时间显著延长(P < 0.05), 净生殖率和内禀增长率显著升高(P < 0.01)(表 1)。
当藻密度为4.0×106个细胞/mL时, 青霉素钠浓度对轮虫的所有生命表统计学参数均无显著性影响(P>0.05)(表 1)。
双因素方差分析结果表明, 食物密度对轮虫的生命期望、净生殖率、内禀增长率和后代混交率均具有显著性影响(P < 0.05), 青霉素钠浓度对轮虫的世代时间、净生殖率和内禀增长率有显著性影响(P < 0.05), 食物密度和青霉素钠浓度的交互作用对轮虫的生命期望、净生殖率和内禀增长率具有显著性影响(P < 0.05)。
2.3 不同食物密度下轮虫生命表统计学参数与青霉素钠浓度间的关系回归分析结果表明, 当食物密度为1.0×106个细胞/mL时, 青霉素钠浓度与轮虫的生命期望、世代时间和净生殖率间均具有显著的剂量—效应关系;当食物密度为2.0×106个细胞/mL时, 青霉素钠浓度与轮虫的生命期望、世代时间和内禀增长率间均具有显著的剂量—效应关系(表 3)。
| 参数 Parameter |
差异源 Source |
平方和 SS |
df | 均方 MS |
F | P |
| 生命期望 | 食物密度(A) | 1488.85 | 2 | 744.43 | 3312.70 | P < 0.01 |
| Life expectancy | 青霉素钠浓度(B) | 1016.13 | 5 | 203.22 | 1.82 | P>0.05 |
| at hatching | A×B | 4695.47 | 10 | 469.55 | 4.19 | P < 0.01 |
| Error | 4032.00 | 36 | 112.00 | |||
| 世代时间 | 食物密度(A) | 183.56 | 2 | 91.78 | 1.73 | P>0.05 |
| Generation time | 青霉素钠浓度(B) | 672.85 | 5 | 134.57 | 2.54 | P < 0.05 |
| A×B | 1003.95 | 10 | 100.40 | 1.90 | P>0.05 | |
| Error | 1907.49 | 36 | 52.99 | |||
| 净生殖率 | 食物密度(A) | 365.46 | 2 | 182.73 | 26.83 | P < 0.01 |
| Net | 青霉素钠浓度(B) | 413.30 | 5 | 82.66 | 12.14 | P < 0.01 |
| reproductive | A×B | 343.27 | 10 | 34.33 | 5.04 | P < 0.01 |
| rate | Error | 245.17 | 36 | 6.81 | ||
| 内禀增长率 | 食物密度(A) | 0.00 | 2 | 0.00 | 7.10 | P < 0.01 |
| Intrinsic rate of population increase | 青霉素钠浓度(B) | 0.00 | 5 | 0.00 | 9.20 | P < 0.01 |
| A×B | 0.00 | 10 | 0.00 | 2.45 | P<0.05 | |
| Error | 0.00 | 36 | 0.00 | |||
| 后代混交率 | 食物密度(A) | 0.01 | 2 | 0.00 | 13.07 | P < 0.01 |
| Proportion of sexual offspring | 青霉素钠浓度(B) | 0.00 | 5 | 0.00 | 0.88 | P> 0.05 |
| A×B | 0.00 | 10 | 0.00 | 1.31 | P>0.05 | |
| Error | 0.01 | 36 | 0.00 |
| 食物密度 Food density/(个细胞/mL) |
参数 Parameters |
回归方程 Regression equation |
显著性检验 Significant test |
| 1.0×106 | 生命期望 | y=-6.071×10-5x2+0.098x+62.629 | R2 =0.614, P < 0.01 |
| 世代时间 | y=-4.1×10-5x2+0.041x+52.55 | R2 =0.352, P < 0.05 | |
| 净生殖率 | y=-2.64×10-5x2+0.042x+5.238 | R2 =0.826, P < 0.01 | |
| 2.0×106 | 生命期望 | y=-6.187x2+0.098x+51.464 | R2 =0.548, P < 0.01 |
| 世代时间 | y=-3.821×10-5x2+0.054x+45.679 | R2 =0.587, P < 0.01 | |
| 内禀增长率 | y=-4.389×10-8x2+6.417×10-5x+0.037 | R2 =0.866, P < 0.01 |
自然水体中, 抗生素的浓度通常在ng/L—μg/L, 因此环境相关浓度的抗生素对轮虫的存活、生殖和种群增长等无显著性影响[1, 4-5]。然而, 亚致死浓度的抗生素却对其具有显著性影响[15-18, 30], 本研究结果亦然。从已报道的研究结果来看, 在亚致死浓度范围内, 一定浓度范围内的抗生素对轮虫存活、生殖和种群增长的影响总体上呈现出“抑制”、“促进”和“低促高抑”等三种格局。首先, 一定浓度的硫酸链霉素、盐酸四环素、酒石酸泰乐霉素[17]和阿莫西林[18]对萼花臂尾轮虫、哈瓦那臂尾轮虫和褶皱臂尾轮虫等的存活、生殖和种群增长具有抑制作用, 且最低可观察效应浓度(LOEC)与抗生素和轮虫的种类有关。其次, 一定浓度范围内的利福平[16]、诺氟沙星[30]和青霉素钠(本研究)对萼花臂尾轮虫的存活、生殖和种群增长具有促进作用。再次, 一定浓度的盐酸土霉素[15]对轮虫存活、生殖和种群增长的影响具有“低促高抑”特点(表 4)。鉴于较低浓度(低于Araujo和McNair[17]所设计的最低测试浓度5.6 mg/L)的利福平[16]、诺氟沙星[30]和青霉素钠(本研究)对萼花臂尾轮虫的存活、生殖和种群增长所具有的促进作用, 以及较高浓度的硫酸链霉素、盐酸四环素、酒石酸泰乐霉素[17]、阿莫西林[18]和盐酸土霉素[15]等抗生素对轮虫的存活、生殖和种群增长所具有的抑制作用, 我们推测“低促高抑”可能是抗生素对轮虫存活、生殖和种群增长影响的主要格局。当然, 这一推测仍需要通过对更多种类的抗生素在更宽的测试浓度和食物密度范围内进行研究加以证实。
| 抗生素 Antibiotics |
测试的浓度范围 Range of tested concentrations/ (mg/L) |
受试轮虫种类 Tested rotifer species |
食物种类和密度 Algal species and density/ (个/L) |
生命期望 Life expectancy at hatching |
净生殖率 Net reproductive rate |
内禀增长率 Intrinsic rate of population increase |
文献来源 References |
|||||
| 促进作用 Prompting effect |
抑制作用 Inhibitory effect |
促进作用 Prompting effect |
抑制作用 Inhibitory effect |
促进作用 Prompting effect |
抑制作用 Inhibitory effect |
|||||||
| 硫酸链霉素 Streptomycin sulfate |
5.6—2000 | 萼花臂尾轮虫B. calyciflorus 褶皱臂尾轮虫B. plicatilis |
CV*, 3×106 NO, 7×106 |
— — |
5.6 90 |
— — |
5.6 90 |
— — |
2000 2000 |
[17] | ||
| 盐酸四环素 Tetracycline hydrochloride |
5.6—2000 | 萼花臂尾轮虫B. calyciflorus 褶皱臂尾轮虫B. plicatilis |
CV, 3×106 NO, 7×106 |
— — |
90 90 |
— — |
5.6 90 |
— — |
2000 2000 |
[17] | ||
| 酒石酸泰乐菌素 Tylosin tartrate |
5.6—2000 | 萼花臂尾轮虫B. calyciflorus 褶皱臂尾轮虫B. plicatilis |
CV, 3×106 NO, 7×106 |
— — |
90 90 |
— — |
90 90 |
— — |
2000 2000 |
[17] | ||
| 阿莫西林 Amoxicillin |
0.05—0.2 | 萼花臂尾轮虫B. calyciflorus 哈瓦那臂尾轮虫B. havanaensis |
CV, 1×106 | — — |
0.05 0.1 |
— — |
0.05 0.05 |
— — |
0.05 0.05 |
[18] | ||
| 利福平 | 2—10 | 萼花臂尾轮虫B. calyciflorus | SO, 1×106 | 2 | — | — | — | — | — | [16] | ||
| Rifampicin | SO, 2×106 | 2 | — | 2 | — | 2 | — | |||||
| SO, 4×106 | 2 | — | 2 | — | 2 | — | ||||||
| 诺氟沙星 | 5—80 | 萼花臂尾轮虫B. calyciflorus | SO, 1×106 | 5 | — | 5 | — | 5 | — | [30] | ||
| Norfloxacin | SO, 2×106 | 5 | — | 5 | — | 5 | — | |||||
| SO, 4×106 | 5 | — | 5 | — | 5 | — | ||||||
| 青霉素钠 | 0.2—1 | 萼花臂尾轮虫B. calyciflorus | SO, 1×106 | 0.4 | — | 0.2 | — | — | — | 本文 | ||
| Penicillin sodium | SO, 2×106 | 0.2 | — | 0.2 | — | 0.2 | — | |||||
| SO, 4×106 | — | — | — | — | — | — | ||||||
| 盐酸土霉素 | 30—180 | 萼花臂尾轮虫B. calyciflorus | SO, 1×106 | 90 | 180 | 30 | 150 | — | 180 | [15] | ||
| Oxytetracycline | SO, 2×106 | 30 | 180 | 30 | 180 | 30 | 180 | |||||
| hydrochloride | SO, 4×106 | 90 | 180 | 30 | 180 | 30 | 180 | |||||
| * CV: 普通小球藻Chlorella vulgaris. NO: 微绿球藻Nannochloropsis oculata. SO: 斜生栅藻Scenedesmus obliquus | ||||||||||||
已有研究结果表明, 与较低和较高的藻密度相比, 中等的藻密度可降低污染物对轮虫存活、生殖和种群增长的抑制作用[20-25]。Jiang等[15]的研究结果表明, 中等的斜生栅藻密度(2.0×106个细胞/mL)不仅降低了较高浓度的盐酸土霉素对萼花臂尾轮虫存活、生殖和种群增长的抑制作用幅度, 而且提高了较低浓度的盐酸土霉素对萼花臂尾轮虫存活、生殖和种群增长的促进作用幅度。本研究中, 中等的斜生栅藻密度也提高了较低浓度的青霉素钠对萼花臂尾轮虫存活、生殖和种群增长的促进作用幅度。翟盼等[16]研究发现较低浓度的利福平对萼花臂尾轮虫生命期望、净生殖率和种群内禀增长率的促进作用幅度随着藻密度由1.0×106个细胞/mL升高至4.0×106个细胞/mL而逐渐显著地增大;这一结果与上述结果的不一致是否可归因于藻类的生长状况, 还有待于进一步的研究。
3.3 抗生素影响轮虫存活、生殖和种群增长的可能机制有关抗生素影响轮虫存活、生殖和种群增长的机制, 目前尚不明确。就较高浓度的抗生素的抑制作用而言, 多数学者将其归结为抗生素的毒性作用[12-13, 15, 17-18, 26]。Wollenberger等[7]在研究盐酸土霉素等抗生素对大型溞(Daphnia magna)存活和生殖的影响时认为, 较高浓度的抗生素通过对藻类的毒性作用从而间接地对枝角类的存活和生殖起抑制作用。但是, 与中等的藻密度相比, 藻类密度的升高和降低均增强了较高浓度的盐酸土霉素对萼花臂尾轮虫存活、生殖和种群增长的抑制作用幅度[15], 提示较高浓度的抗生素的抑制作用可能主要源于其毒性作用。
就较低浓度的抗生素对轮虫存活、生殖和种群增长的促进作用而言, 三个主要的机制先后被提出。首先, 利福平和盐酸土霉素等抗生素通过有效杀灭轮虫体内和喂食的藻类中可能存在的一些有害细菌, 从而促进了轮虫的存活、生殖和种群增长[15-16]。其次, 鉴于利福平和布洛芬等可能具有的环境雌激素效应[16, 25], 而一部分已被证明具有环境雌激素效应的低浓度污染物如Cd2+[22]、Zn2+[24]、艾氏剂[31]、狄氏剂[32]和十氯丹[33]等可促进轮虫存活、生殖和种群增长, 因此较低浓度的利福平等抗生素的促进作用被部分归因于其雌激素效应[16]。再次, 由于盐酸四环素[26]等抗生素具有和Hg2+[34]等污染物相似的“低促高抑”作用, 毒物兴奋效应(hormesis)[35]被用于解释较低浓度的抗生素对轮虫存活、生殖和种群增长的促进作用[26]。包括青霉素钠等在内的较低浓度的抗生素究竟是通过哪个或那几个机制促进轮虫存活、生殖和种群增长, 仍需进一步研究。
3.4 轮虫有性生殖与抗生素种类和浓度以及食物密度之间的关系单巢纲轮虫具有典型的周期性孤雌生殖的繁殖方式。通常情况下, 单巢纲轮虫主要以孤雌生殖的方式繁殖;但在特定的条件下, 如较低的食物质量和较高的种群密度, 单巢纲轮虫进行有性生殖, 产生休眠卵, 以适应不利的环境条件。已有研究表明, 在1.0×106、2.0×106和4.0×106个/mL的斜生栅藻密度下, 2.0—10.0 mg/L的利福平均显著提高萼花臂尾轮虫的后代混交率, 其中1.0×106和4.0×106个/mL的斜生栅藻密度下后代混交率提高幅度较大[16]。在1.0×106、2.0×106和4.0×106个/mL的斜生栅藻密度下, 13.5— 40.5 mg/L、40.5 mg/L和40.5 mg/L的盐酸四环素显著地提高萼花臂尾轮虫的后代混交率, 其中1.0×106和4.0×106个/mL的斜生栅藻密度下后代混交率提高幅度也较大[26]。与上述研究结果不同的是, 三个斜生栅藻密度下, 盐酸土霉素浓度对萼花臂尾轮虫的后代混交率却无显著性影响[15]。与Jiang等[15]的研究结果相一致, 本研究结果表明, 各食物密度下, 青霉素钠浓度对萼花臂尾轮虫的后代混交率也无显著性影响。看来, 轮虫有性生殖的发生幅度与抗生素的种类和浓度等有关。
3.5 轮虫各生命表统计学参数对抗生素污染的敏感性综合现有的研究结果来看, 萼花臂尾轮虫各生命表统计学参数对抗生素污染的敏感性因抗生素种类和食物密度的不同而存在着差异。在1.0×106、2.0×106和4.0×106个细胞/mL等三个斜生栅藻密度下, 生命期望和后代混交率对利福平污染均具有较高的敏感性[16], 净生殖率对盐酸土霉素污染具有较高的敏感性[15]。在1.0×106和2.0×106个细胞/mL等两个斜生栅藻密度下, 世代时间对盐酸四环素具有较高的敏感性[26];与此相似的是, 本研究结果表明, 在1.0×106和2.0×106个细胞/mL等两个斜生栅藻密度下, 世代时间和净生殖率对青霉素钠均具有较高的敏感性。
| [1] |
Carvalho T I, Santos L. Antibiotics in the aquatic environments: a review of the European scenario. Environment International, 2016, 94: 736-757. DOI:10.1016/j.envint.2016.06.025 |
| [2] |
Sarmah A K, Meyer M T, Boxall A B A. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environment. Chemosphere, 2006, 65(5): 725-759. DOI:10.1016/j.chemosphere.2006.03.026 |
| [3] |
Kümmerer K. Antibiotics in the aquatic environment-A review-Part Ⅰ. Chemosphere, 2009, 75(4): 417-434. DOI:10.1016/j.chemosphere.2008.11.086 |
| [4] |
Jiang L, Hu X L, Yin D Q, Zhang H C, Yu Z Y. Occurrence, distribution and seasonal variation of antibiotics in the Huangpu River, Shanghai, China. Chemosphere, 2011, 82(6): 822-828. DOI:10.1016/j.chemosphere.2010.11.028 |
| [5] |
Tong L, Huang S B, Wang Y X, Liu H, Li M J. Occurrence of antibiotics in the aquatic environment of Jianghan Plain, central China. Science of the Total Environment, 2014, 497-498: 180-187. DOI:10.1016/j.scitotenv.2014.07.068 |
| [6] |
Davies J, Davies D. Origins and evolution of antibiotic resistance. Microbiology and Molecular Biology Reviews, 2010, 74(3): 417-433. DOI:10.1128/MMBR.00016-10 |
| [7] |
Wollenberger L, Halling-Sørensen B, Kusk K O. Acute and chronic toxicity of veterinary antibiotics to Daphnia magna. Chemosphere, 2000, 40(7): 723-730. DOI:10.1016/S0045-6535(99)00443-9 |
| [8] |
Isidori M, Lavorgna M, Nardelli A, Pascarella L, Parrella A. Toxic and genotoxic evaluation of six antibiotics on non-target organisms. Science of the Total Environment, 2005, 346(1/3): 87-98. |
| [9] |
Bownik A, Ślaska B, Bochra J, Gumieniak K, Gałek K. Procaine penicillin alters swimming behaviour and physiological parameters of Daphnia magna. Environmental Science and Pollution Research, 2019, 26(2): 18662-18673. |
| [10] |
Snell T W, Janssen C R. Rotifers in ecotoxicology: a review. Hydrobiologia, 1995, 313-314(1): 231-247. DOI:10.1007/BF00025956 |
| [11] |
Dahms H U, Hagiwara A, Lee J S. Ecotoxicology, ecophysiology, and mechanistic studies with rotifers. Aquatic Toxicology, 2011, 101(1): 1-12. DOI:10.1016/j.aquatox.2010.09.006 |
| [12] |
王金秋, 周叶平, 林隽姬, 王天宇. 两种抗生素对褶皱臂尾轮虫种群增长的影响. 复旦学报: 自然科学版, 2008, 47(3): 347-353. |
| [13] |
张雄. 蓝藻和抗生素对萼花臂尾轮虫种群生态学的影响研究[D]. 武汉: 中南民族大学, 2012.
|
| [14] |
El-Bassat R A, Touliabah H E, Harisa G I. Toxicity of four pharmaceuticals from different classes to isolated plankton species. African Journal of Aquatic Science, 2012, 37(1): 71-80. DOI:10.2989/16085914.2012.666376 |
| [15] |
Jiang S, Xi Y L, Zhu H, Zhang B X, Yu J H. Combined effects of oxytetracycline concentration and algal food level on the life-table demography of Brachionus calyciflorus (Rotifera). Annales de Limnologie-International Journal of Limnology, 2018, 54: 28. DOI:10.1051/limn/2018020 |
| [16] |
翟盼, 温新利, 陈治文, 赵政, 李海洋, 席贻龙. 抗生素利福平对萼花臂尾轮虫生命表参数的影响. 中国环境科学, 2016, 36(6): 1886-1894. DOI:10.3969/j.issn.1000-6923.2016.06.043 |
| [17] |
Araujo A, Mcnair J N. Individual- and population-level effects of antibiotics on the rotifers, Brachionus calyciflorus and B. plicatilis. Hydrobiologia, 2007, 593(1): 185-199. DOI:10.1007/s10750-007-9044-y |
| [18] |
González-Pérez B K, Sarma S S S, Nandini S. Effects of selected pharmaceuticals (ibuprofen and amoxicillin) on the demography of Brachionus calyciflorus and Brachionus havanaensis (Rotifera). The Egyptian Journal of Aquatic Research, 2016, 42(3): 341-347. DOI:10.1016/j.ejar.2016.09.003 |
| [19] |
Rao T R, Sarma S S S. Demographic parameters of Brachionus patulus Muller (Rotifera) exposed to sublethal DDT concentrations at low and high food levels. Hydrobiologia, 1986, 139(3): 193-200. DOI:10.1007/BF00028292 |
| [20] |
Sarma S S S, Nandini S, Flores J L G. Effect of methyl parathion on the population growth of the rotifer Brachionus patulus (O. F. Müller) under different algal food (Chlorella vulgaris) densities. Ecotoxicology and Environmental Safety, 2001, 48(2): 190-195. DOI:10.1006/eesa.2000.2014 |
| [21] |
Gama-Flores J L, Sarma S S S, Nandini S. Acute and chronic toxicity of the pesticide methyl parathion to the rotifer Brachionus angularis (Rotifera) at different algal (Chlorella vulgaris) food densities. Aquatic Ecology, 2004, 38(1): 27-36. DOI:10.1023/B:AECO.0000020986.92471.32 |
| [22] |
石娟, 席贻龙, 杨琳璐, 汪圣广, 陈枫华, 苏田娟. 不同藻密度下Cd2+浓度对萼花臂尾轮虫生命表统计学参数的影响. 应用生态学报, 2010, 21(6): 1614-1620. |
| [23] |
Xu X P, Xi Y L, Chu Z X, Xiang X L. Effects of DDT and dicofol on population growth of Brachionus calyciflorus under different algal (Scenedesmus obliquus) densities. Journal of Environmental Biology, 2014, 35(5): 907-916. |
| [24] |
赵清清, 席贻龙, 李志超, 潘玲. 不同藻密度下Zn2+浓度对萼花臂尾轮虫实验种群增长参数的影响. 生态学报, 2015, 35(12): 4026-4033. |
| [25] |
黄克强, 席贻龙, 朱韩, 张丙行, 潘玲. 不同藻密度下布洛芬浓度对萼花臂尾轮虫生命表统计学参数的影响. 生态学报, 2019, 39(3): 1105-1113. |
| [26] |
项贤领, 朱晔璘, 徐秋磊, 朱凌云, 席贻龙. 盐酸四环素浓度和食物密度对萼花臂尾轮虫生活史特征的综合影响. 生态学报, 2017, 37(22): 7718-7728. |
| [27] |
US EPA. Methods for measuring the acute toxicity of effluents to freshwater and marine organisms. 3rd ed. Cincinnati, Ohio: U. S. Environmental Protection Agency, 1985.
|
| [28] |
章宗涉, 黄祥飞. 淡水浮游生物研究方法. 北京: 科学出版社, 1991.
|
| [29] |
Chen H, Li X J, Zhu S C. Occurrence and distribution of selected pharmaceuticals and personal care products in aquatic environments: a comparative study of regions in China with different urbanization levels. Environmental Science and Pollution Research, 2012, 19(6): 2381-2389. DOI:10.1007/s11356-012-0750-2 |
| [30] |
朱韩, 席贻龙, 徐秋磊, 黄克强, 李加男, 江姗. 不同藻密度下诺氟沙星对萼花臂尾轮虫生命表统计学参数的影响. 生态毒理学报, 2019, 14(3): 163-173. |
| [31] |
Huang L, Xi Y L, Zha C W, Zhao L L. Effect of aldrin on life history characteristics of rotifer Brachionus calyciflorus Pallas. Bulletin of Environmental Contamination and Toxicology, 2007, 79(5): 524-528. DOI:10.1007/s00128-007-9271-y |
| [32] |
Huang L, Xi Y-L, Zha C W, Zhao L L, Wen X L. Effects of dieldrin and 17β-estradiol on life history characteristics of freshwater rotifer Brachionus calyciflorus Pallas. Journal of Freshwater Ecology, 2012, 27(3): 381-392. DOI:10.1080/02705060.2012.668499 |
| [33] |
Zha C W, Xi Y L, Huang L, Zhao L L. Effect of sublethal exposure to chlordecone on life history characteristics of freshwater rotifer Brachionus calyciflorus Pallas. Bulletin of Environmental Contamination and Toxicology, 2007, 78(1): 79-83. DOI:10.1007/s00128-007-9003-3 |
| [34] |
Pan L, Xi Y L, Li Z C, Zhao Q Q, Hu Z J. Effects of mercury on the life table demography of the rotifer Brachionus calyciflorus under different algal food (Scenedesmus obliquus) densities. Acta Ecologica Sinica, 2016, 36(3): 218-223. DOI:10.1016/j.chnaes.2016.04.009 |
| [35] |
Calabrese E J. Hormesis: why it is important to toxicology and toxicologists. Environmental Toxicology and Chemistry, 2008, 27(7): 1451-1474. DOI:10.1897/07-541.1 |