 2021, Vol. 41
2021, Vol. 41文章信息
- 贾真真, 李苇洁, 田奥, 王加国
- JIA Zhenzhen, LI Weijie, TIAN Ao, WANG Jiaguo
- 贵州百里杜鹃风景区旅游干扰对杜鹃群落特征的影响
- Ecological influence of tourism disturbance on the characteristics of different communities in Baili Rhododendron forest
- 生态学报. 2021, 41(11): 4641-4649
- Acta Ecologica Sinica. 2021, 41(11): 4641-4649
- http://dx.doi.org/10.5846/stxb201907021387
-
文章历史
- 收稿日期: 2019-07-02
- 网络出版日期: 2021-04-07





自然资源作为旅游景区的基础会随着旅游活动的增加受到强烈干扰, 揭示旅游活动对旅游生态系统的影响是对旅游资源进行有效保护的必要前提[1-2]。不同旅游干扰方式及强度可直接影响植被、土壤等环境[3]进而改变群落的的结构及多样性等群落特征[4-6]。如踩踏可磨损植被, 使土壤裸露、移动和孔隙度降低等, 修建步道小径可严重破坏周围植被土壤并影响土壤水文功能和导致水土流失等[7-8]。而不同植物类型受物种组成、植被生理特征、群落结构等的影响对旅游干扰及其强度的响应也不同, 如朱珠等研究了旅游干扰对九寨沟自然保护区生态影响, 发现耐荫喜湿的乡土植物局部消失, 而喜旱耐扰动的植物种群扩大[9]。
百里杜鹃景区位于贵州省大方县与黔西县交界处, 属国家5A级旅游景区、国家生态旅游示范区, 它以地球同纬度上面积最大的原生杜鹃林带为主体, 面积125.8km2, 海拔1580—1845m。在长约50km, 宽1.2—5.3km的狭长丘陵上, 分布着以马缨杜鹃(Rhododendron delavayi)、露珠杜鹃(Rhododendron irroratum)、迷人杜鹃(Rhododendron agastum)、映山红(Rhododendron simsii)为优势种的杜鹃属植物约35个种(含亚种、变种), 分属6个亚属、4个组及9个亚组[10], 是重要的杜鹃种质资源库, 具有极高的科研和旅游价值。百里杜鹃景区以金坡花区和普底花区景观最为集中连片, 每年3—5月杜鹃花期都吸引大量游客, 旅游的季节性明显[11]。自1993年举办首届“中国贵州百里杜鹃花节”以来, 百里杜鹃景区每到花期就会受到集中性旅游活动的冲击, 致使林带边缘与游径两侧践踏现象明显, 枯落物厚度明显减少, 土壤板结[12]。而这一趋势随着景区经济效益的不断增加还会持续的对杜鹃群落结构及其物种组成等造成冲击。本研究拟通过旅游干扰程度对不同杜鹃群落的物种多样性指及群落结构的影响分析, 探讨不同旅游干扰类型及强度对杜鹃群落特征的影响, 以期为百里杜鹃景区开发利用和科学管理提供理论依据。
1 研究方法 1.1 样地选择百里杜鹃有两种典型群落, 分别为杜鹃纯林和杜鹃混交林, 本研究于2017年8月采用典型取样的方法对百里杜鹃景区内杜鹃纯林及杜鹃混交林进行调查, 调查样地在金坡乡奢香驿山顶—索马蔚—马缨林、普底乡醉九牛以及黄泥乡王家寨—嘎木等3条主要游线, 分别于主干道两侧随机选取样地, 设置大小为20m×20m的调查样地, 调查样地中的全部灌木的地径、株高、冠幅及盖度;并在每个样地中随机布设10个大小为2.5m×2.5m的草本样方, 调查样方内草本植物的种类、株高、株数、盖度等;每个样地中设置5个50cm×50cm枯落物样方测量枯落物厚度。
1.2 样地旅游干扰等级分析旅游影响系数(Tourism influencing index, 简称TII)[13-15]是反映旅游活动对植被景观干扰的一种综合程度指标, 因此用其进行旅游干扰分级。百里杜鹃景区对游人折枝行为进行了重点监管, 所以本研究中未予考虑。在本研究中, 旅游影响系数采用分级赋值方式(表 2)进行计算, 即:TII=Cm+Cr+Ct+Cw, 其中Cm为草本层现状系数, 利用草本层盖度表示, 盖度愈大, 影响程度愈小;Cr为垃圾影响系数, 垃圾数量越多, 旅游活动影响越大;Ct为踩踏影响系数, 利用枯落层厚度说明踩踏程度, 厚度越大, 影响程度越小;Cw为林木更新影响系数, 利用树木幼苗量表示, 幼苗越多, 自然更新越强, 旅游影响越小[15]。旅游影响因子赋值见表 2, 旅游影响因子权重参照文献[12], 即W(Cm)=0.0752, W(Cr)=0.2653, W(Ct) =0.5083, W(Cw)=0.1512。将所调查样地依旅游影响系数划分为轻度干扰(< 0.33)、中度干扰(0.33-0.67)与重度干扰(>0.67)三个等级。
| 序号 No. |
群落名称 Community |
地理坐标 Geographic coordinates |
地点 Site |
海拔 Altitude |
坡向 Slope direction |
坡度 Slope grade |
优势种 Dominant species |
| 1 | 杜鹃纯林 | E105°56′10.56″, N27°12′05.83″ | 金坡 | 1688 | 西北 | 13 | 露珠杜鹃 |
| 2 | 杜鹃纯林 | E105°55′33.26″, N27°11′53.93″ | 金坡 | 1638 | 南 | 5 | 露珠杜鹃 |
| 3 | 杜鹃纯林 | E105°55′30.21″, N27°11′49.83″ | 金坡 | 1633 | 西北 | 3 | 露珠杜鹃 |
| 4 | 杜鹃纯林 | E105°57′54.91″, N27°10′47.29″ | 金坡 | 1592 | 北 | 3 | 马缨杜鹃 |
| 5 | 杜鹃纯林 | E105°57′55.22″, N27°10′48.05″ | 金坡 | 1563 | 北 | 7 | 马缨杜鹃 |
| 6 | 杜鹃纯林 | E105°57′55.90″, N27°10′48.59″ | 金坡 | 1457 | 北 | 3 | 马缨杜鹃 |
| 7 | 杜鹃纯林 | E105°51′58.31″, N27°14′36.78″ | 普底 | 1766 | 北 | 5 | 马缨杜鹃 |
| 8 | 杜鹃纯林 | E105°51′52.94″, N27°14′16.85″ | 普底 | 1733 | 北 | 5 | 马缨杜鹃 |
| 9 | 杜鹃纯林 | E105°51′52.86″, N27°14′18.21″ | 普底 | 1723 | 北 | 5 | 马缨杜鹃 |
| 10 | 杜鹃纯林 | E106°03′44.77″, N27°15′42.68″ | 黄泥 | 1476 | 北 | 5 | 马缨杜鹃 |
| 11 | 杜鹃纯林 | E106°03′59.02″, N27°15′23.54″ | 黄泥 | 1453 | 北 | 2 | 马缨杜鹃 |
| 12 | 杜鹃纯林 | E106°03′56.47″, N27°15′31.00″ | 黄泥 | 1435 | 北 | 7 | 马缨杜鹃 |
| 13 | 杜鹃混交林 | E105°56′50.17″, N27°11′19.73″ | 金坡 | 1634 | 北 | 6 | 马缨杜鹃 |
| 14 | 杜鹃混交林 | E105°56′50.56″, N27°11′20.17″ | 金坡 | 1623 | 南 | 6 | 露珠杜鹃 |
| 15 | 杜鹃混交林 | E105°56′51.16″, N27°11′19.08″ | 金坡 | 1582 | 北 | 5 | 露珠杜鹃 |
| 16 | 杜鹃混交林 | E105°51′51.33″, N27°13′46.40″ | 普底 | 1678 | 北 | 3 | 露珠杜鹃 |
| 17 | 杜鹃混交林 | E105°51′51.30″, N27°13′52.04″ | 普底 | 1693 | 南 | 3 | 马缨杜鹃 |
| 18 | 杜鹃混交林 | E105°51′50.56″, N27°13′51.49″ | 普底 | 1692 | 北 | 3 | 马缨杜鹃 |
| 19 | 杜鹃混交林 | E106°03′42.47″, N27°15′43.79″ | 黄泥 | 1456 | 西南 | 5 | 马缨杜鹃 |
| 20 | 杜鹃混交林 | E106°03′41.50″, N27°15′47.80″ | 黄泥 | 1463 | 北 | 6 | 马缨杜鹃 |
| 21 | 杜鹃混交林 | E106°04′04.45″, N27°15′22.56″ | 黄泥 | 1393 | 北 | 3 | 马缨杜鹃 |
| 22 | 杜鹃混交林 | E106°03′44.95″, N27°15′43.21″ | 黄泥 | 1475 | 西 | 3 | 马缨杜鹃 |
| 23 | 杜鹃混交林 | E105°51′52.11″, N27°14′17.90″ | 普底 | 1727 | 北 | 3 | 马缨杜鹃 |
| 24 | 杜鹃混交林 | E105°51′51.77″, N27°14′19.33″ | 普底 | 1678 | 北 | 3 | 马缨杜鹃 |
| 25 | 杜鹃混交林 | E105°51′57.34″, N27°14′36.89″ | 普底 | 1765 | 北 | 3 | 马缨杜鹃 |
| 垃圾量 Quantity of garbage |
枯层厚 Litter thickness |
幼苗量 Seedlings |
草本层盖度 Herb layer coverage |
|||||||
| 标准(Num.) | 赋值 | 标准/cm | 赋值 | 标准(Num.) | 赋值 | 标准/% | 赋值 | |||
| 0—5 | 0.1 | 0—1.0 | 1.0 | 0 | 1.0 | 0—10 | 1.0 | |||
| 6—10 | 0.2 | 1.1—2.0 | 0.9 | 1—10 | 0.9 | 11—20 | 0.9 | |||
| 11—15 | 0.3 | 2.1—3.0 | 0.8 | 11—20 | 0.8 | 21—30 | 0.8 | |||
| 16—20 | 0.4 | 3.1—4.0 | 0.7 | 21—30 | 0.7 | 31—40 | 0.7 | |||
| 21—25 | 0.5 | 4.1—5.0 | 0.6 | 31—40 | 0.6 | 41—50 | 0.6 | |||
| 26—30 | 0.6 | 5.1—6.0 | 0.5 | 41—50 | 0.5 | 51—60 | 0.5 | |||
| 31—35 | 0.7 | 6.1—7.0 | 0.4 | 51—60 | 0.4 | 61—70 | 0.4 | |||
| 36—40 | 0.8 | 7.1—8.0 | 0.3 | 61—70 | 0.3 | 71—80 | 0.3 | |||
| 41—45 | 0.9 | 8.1—9.0 | 0.2 | 71—80 | 0.2 | 81—90 | 0.2 | |||
| >45 | 1.0 | >9.0 | 0.1 | >80 | 0.1 | 91—100 | 0.1 | |||
(1) 重要值(Importance value. 简称Ⅳ)是评估群落物种多样性、衡量物种优势度的重要指标[16], 计算公式为:

|
式中:相对多度(%)=100×(某个物种的株数/所有物种的总株数);相对频度(%)= 100×(某个物种在统计样方中出现的次数/所有物种出现的总次数);相对优势度(%)=100×(某个乔灌木物种的胸高断面积/所有乔灌木物种的胸高断面积和)或100×(某个草本物种的盖度/所有草本物种的盖度和)。
(2) 物种多样性(Species diversity, 简称SD), 主要包括丰富度、均匀度和多样性三个方面, 它是对群落结构和功能复杂性的度量[17]。
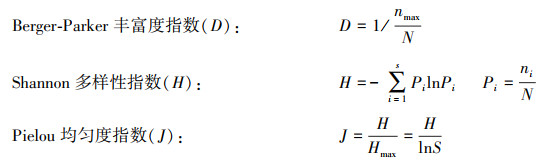
|
其中:S为出现在样地的物种数, ni为第i个物种个体数, N为所有物种个体总数, nmax为个体数量最多物种的个体数。
1.4 数据处理本研究用Canoco5.0软件对环境变量与物种多样性、植被结构的对应关系进行冗余分析(RDA), 在冗余分析中将海拔、坡度、坡向、坡位、踩踏指数、垃圾指数等定义为环境变量, 将坡位用数字等级表示, 下坡位为1, 中坡位为2, 上坡位为3;对坡向以等级制表示, 将坡向按照45°的夹角, 以正北为0°, 顺时针方向旋转分为8个坡向等级, 以数字表示各等级:1表示北坡, 2表示东北坡, 3表示西北坡, 4表示东坡, 5表示西坡, 6表示东南坡, 7表示西南坡, 8表示南坡, 数字越大, 表示坡向越向阳[18];踩踏指数与垃圾指数采用踩踏影响系数与垃圾影响系数。并通过SPSS19.0软件进行Pearson相关分析并检验结果在0.05水平的显著性, 图表绘制采用Excel 2010完成。
2 结果与分析 2.1 不同杜鹃群落物种组成在调查的所有杜鹃样地中, 共有35科57属78种木本植物, 32科45属52种草本植物, 其中12个杜鹃纯林样地有乔木14种、灌木51种、草本32种, 共97种;14个杜鹃混交林样地有乔木19种、灌木56种、草本40种, 共115种。因杜鹃群落为灌木林, 调查样地中灌木物种数量优势明显, 杜鹃混交林样地物种总数明显高于杜鹃纯林样地(见图 1)。

|
| 图 1 样地物种数量对比图 Fig. 1 Species quantity comparison of sample plots |
杜鹃纯林和杜鹃混交林均有明显优势种, 且物种组成有一定的相似性(表 3)。杜鹃纯林重要值的前三位为马缨杜鹃、箭竹和露珠杜鹃, 马缨杜鹃占群落绝对优势, 其重要值为37.15;杜鹃混交林重要值前三位为马缨杜鹃、露珠杜鹃和映山红, 马缨杜鹃重要值为31.11。
| 排名 Rank |
杜鹃纯林 Pure Rhododendron forests |
重要值(Ⅳ) Importance value |
排名 Rank |
杜鹃混交林 Mixed Rhododendron forests |
重要值(Ⅳ) Importance value |
|
| 1 | 马缨杜鹃 | 37.15 | 1 | 马缨杜鹃 | 31.11 | |
| 2 | 箭竹 | 19.93 | 2 | 露珠杜鹃 | 12.91 | |
| 3 | 露珠杜鹃 | 11.82 | 3 | 映山红 | 11.67 | |
| 4 | 映山红 | 5.08 | 4 | 迷人杜鹃 | 7.02 | |
| 5 | 小果南烛 | 3.16 | 5 | 箭竹 | 4.04 | |
| 6 | 迷人杜鹃 | 2.79 | 6 | 麻栎 | 3.11 | |
| 7 | 白栎 | 2.36 | 7 | 细齿叶柃 | 2.89 | |
| 8 | 栓皮桦 | 1.70 | 8 | 小果南烛 | 2.81 | |
| 9 | 山矾 | 1.23 | 9 | 板栗 | 2.52 | |
| 10 | 山鸡椒 | 1.20 | 10 | 白栎 | 2.47 |
由表 4可以看出, 不同杜鹃群落在不同旅游强度干扰下, Berger-Parker丰富度指数、Shannon多样性指数以及Pielou均匀度指数差异明显。在总群落中, 杜鹃纯林中度干扰时丰富度指数为4.643、多样性指数为3.041、均匀度指数为0.777, 均明显高于轻度干扰与重度干扰, 总趋势为中度干扰>轻度干扰>重度干扰;杜鹃混交林在中度干扰下丰富度指数、多样性指数、均匀度指数分别为4.688、2.992、0.743, 亦高于轻度干扰和重度干扰, 其趋势则是中度干扰>重度干扰>轻度干扰。
| 群落类型 Community |
丰富度指数DBerger-Parker index | 多样性指数HShannon index | 均匀度指数JPielou index | |||||||||
| Ⅰ | Ⅱ | Ⅲ | Ⅰ | Ⅱ | Ⅲ | Ⅰ | Ⅱ | Ⅲ | ||||
| 杜鹃纯林 | AL | 1.500 | 1.772 | 1.098 | 0.824 | 1.333 | 0.392 | 0.750 | 0.520 | 0.283 | ||
| Pure Rhododendron forests | SL | 6.333 | 5.279 | 1.165 | 2.502 | 2.749 | 0.786 | 0.948 | 0.816 | 0.241 | ||
| HL | 1.275 | 3.735 | 3.280 | 0.926 | 2.096 | 2.020 | 0.402 | 0.740 | 0.766 | |||
| TC | 1.505 | 4.613 | 1.494 | 1.604 | 3.041 | 1.551 | 0.492 | 0.777 | 0.415 | |||
| 杜鹃混交林 | AL | 2.515 | 1.581 | 2.154 | 1.941 | 1.189 | 1.502 | 0.736 | 0.611 | 0.684 | ||
| Mixed Rhododendron | SL | 3.469 | 4.219 | 2.663 | 2.544 | 2.669 | 1.702 | 0.668 | 0.759 | 0.614 | ||
| forests | HL | 2.057 | 2.862 | 3.790 | 1.890 | 1.744 | 2.589 | 0.621 | 0.573 | 0.864 | ||
| TC | 3.519 | 4.688 | 3.985 | 2.537 | 2.992 | 2.878 | 0.703 | 0.743 | 0.694 | |||
| 其中:Ⅰ, 轻度干扰Mild interference;Ⅱ, 中度干扰Moderate interference;Ⅲ, 重度干扰Severe interference;AL, 乔木层Arbor layer;SL, 灌木层Shrub layer;HL, 草本层Herb layer;TC, 总群落Total community | ||||||||||||
从乔本层来看, 杜鹃纯林中度干扰时丰富度指数和多样性指数最高, 分别为1.772和1.333, 而均匀度指数却是轻度干扰时最高为0.750;杜鹃混交林则是轻度干扰时丰富度指数、多样性指数、均匀度指数最高, 分别为2.515、1.941、0.736。在灌木层中, 杜鹃纯林丰富度指数和均匀度指数均随干扰程度增加而降低, 丰富度指数由6.333降至5.279再到1.165, 均匀度指数由0.948降至0.816再到0.241, 降幅较大。而在草本层中, 杜鹃纯林中度干扰时丰富度指数和多样性指数最高, 杜鹃混交林的丰富度指数和多样性指数均随干扰程度的增加而增加, 重度干扰时丰富度指数最高, 分别为3.790和2.589。
2.3 环境变量与杜鹃群落多样性的RDA分析对杜鹃群落样地物种多样性和结构数据进行除趋势对应分析(Detrended Correspondence Analysis, DCA),结果中第一轴的lengths of gradient分别为0.61和1.33, 均小于3, 说明数据适于进行RDA分析。通过RDA分析(见图 2), 6个环境因子对物种多样性的解释率为39.9%, 第一轴解释了39.5%的变量信息, 杜鹃群落物种多样性指数(D、H、J)与垃圾指数、踩踏指数相关性呈正相关且关系密切, 与坡位(Pos)、海拔(ELE)呈负相关;杜鹃群落物种数(N)与踩踏指数关系最为密切;而6个环境因子对杜鹃群落植被结构的解释率仅为32.84%, 第一轴解释率为18.76%, 第二轴解释率为10.87%, 其中垃圾指数(Gar)与杜鹃群落草本层盖度呈明显的负相关, 踩踏指数(Tra)与草本均高呈明显正相关, 与其他结构指标亦有明显相关性。

|
| 图 2 杜鹃群落环境变量与物种多样性、植被结构的冗余分析(RDA) Fig. 2 Redundancy analysis (RDA) ordination of environmental variables and species diversity and vegetation structure of Rhododendron communities Asp, 坡向Slope Direction;Slo, 坡度Slope Grade;Pos, 坡位Slope position;ELE, 海拔Altitude;Gar, 垃圾指数(垃圾影响系数) Garbage index;Tra, 踩踏指数(踩踏影响系数) Trample index, N:物种数量Species number;H-A, 乔木层均高Average height of Arbor layer;C-A, 乔木层盖度Coverage of Arbor layer;H-S, 灌木层均高Average height of Shrub layer;C-S, 灌木层盖度Coverage of Shrub layer;H-H, 草本层均高Average height of Herb layer;C-H, 草本层盖度Coverage of Herb layer |
将各样地杜鹃纯林、混交林的垃圾指数、踩踏指数与群落多样性指数及物种数做相关分析, 结果如由表 5所示。在杜鹃纯林中, 踩踏指数与各多样性指标均无显著相关性(P>0.05);垃圾指数与多样性指数、物种数呈显著负相关(P < 0.05)。而在混交林中, 垃圾指数及踩踏指数均与群落多样性指数及物种数无显著相关性。
| 丰富度D Berger-Parker |
多样性H Shannon |
均匀度J Pielou |
物种数N Species number |
||
| 杜鹃纯林 | 垃圾指数 | -0.444 | -0.586* | -0.399 | -0.614* |
| Pure Rhododendron forests | 踩踏指数 | 0.422 | 0.393 | 0.248 | 0.331 |
| 杜鹃混交林 | 垃圾指数 | -0.019 | -0.117 | 0.242 | -0.313 |
| Mixed Rhododendron forests | 踩踏指数 | -0.290 | -0.444 | -0.150 | -0.436 |
| *表示在0.05水平上显著相关, **表示在0.01水平上显著相关 | |||||
将各样地杜鹃纯林及混交林的垃圾指数、踩踏指数与杜鹃群落生长结构做相关分析, 结果如表 6所示。在杜鹃纯林中, 垃圾指数与草本均高呈极显著负相关(P < 0.01), 与其他群落生长结构均无显著相关性, 踩踏指数与草本均高呈显著正相关, 与灌木均高、灌木盖度呈极显著负相关。在杜鹃混交林中, 垃圾指数与各生长指标均无显著相关性, 踩踏指数与草本均高呈显著正相关。
| 乔木均高 H-A |
乔木盖度 C-A |
灌木均高 H-S |
灌木盖度 C-S |
草本均高 H-H |
草本盖度 C-H |
||
| 杜鹃纯林 | 垃圾指数 | 0.063 | 0.135 | 0.299 | 0.136 | -0.742** | -0.559 |
| Pure Rhododendron forests | 踩踏指数 | -0.136 | -0.106 | -0.738** | -0.798** | 0.610* | 0.189 |
| 杜鹃混交林 | 垃圾指数 | 0.335 | -0.096 | 0.269 | -0.171 | 0.012 | -0.200 |
| Mixed Rhododendron forests | 踩踏指数 | -0.145 | -0.067 | 0.013 | 0.207 | 0.667* | -0.147 |
| H-A: 乔木层均高Average height of Arbor layer;C-A: 乔木层盖度Coverage of Arbor layer;H-S: 灌木层均高Average height of Shrub layer;C-S: 灌木层盖度Coverage of Shrub layer;H-H: 草本层均高Average height of Herb layer;C-H: 草本层盖度Coverage of Herb layer | |||||||
将与干扰指数显著相关的多样性指标、生长结构因子筛选出来, 建立多样性指数、生长结构随干扰指数的回归方程, 如图 3所示。由图可知, 杜鹃纯林Shannon多样性指数、物种数、草本均高与垃圾指数成线性关系, 草本均高、灌木均高、灌木盖度与踩踏指数亦呈线性关系。Shannon多样性指数、物种数、草本均高均随垃圾指数的增加而减少, 垃圾指数每增加一个单元(0.1), Shannon多样性指数降低0.29, 物种数量减少2.5个, 草本均高降低0.11m。当杜鹃群落无垃圾干扰时, Shannon多样性指数为2.644, 物种数为23个, 草本均高为0.7m。草本均高随踩踏指数的增加而增加, 踩踏指数每增加一个单位(0.1), 草本均高增加0.05m;灌木均高和灌木盖度则随踩踏指数的增加而降低, 踩踏指数每增加一个单位, 灌木均高减少0.45m, 灌木盖度减少0.92。在杜鹃混交林样地中, 草本均高与踩踏指数呈正相关, 踩踏指数每增加一个单位, 草本均高增加0.09m。

|
| 图 3 杜鹃纯林多样性指数、生长结构对不同干扰的响应 Fig. 3 The response of diversity index and growth structure of pure Rhododendron forests to different disturbances |
杜鹃纯林与杜鹃混交林的主要优势物种存在一定的相似性, 重要值前十的木本植物中均包含马缨杜鹃、露珠杜鹃、映山红、迷人杜鹃、小果南烛等杜鹃花科植物。马缨杜鹃为百里杜鹃的主要建群种[19-20], 因此在两种杜鹃群落中, 马缨杜鹃分别以37.15和31.11的最高重要值占据优势地位。箭竹为其伴生种, 这在西南其他地区很是常见[21-22], 箭竹成集群分布, 株数多, 受人为干扰弱, 因此在重度干扰下的杜鹃群落中能占据更大的生态位。此外, 在杜鹃纯林和杜鹃混交林重要值前十的木本植物中还有相同的天然次生先锋树种—白桦, 其扩展能力强, 适生范围宽, 在杜鹃纯林和杜鹃混交林中的重要值均在2—3之间, 相对稳定。
3.2 旅游干扰对杜鹃群落物种多样性影响通过冗余分析(RDA)可知, 环境变量解释杜鹃群落多样性、结构的贡献率仅为39.9%和32.84%, 表明坡度、坡向、坡位、海拔以及踩踏指数、垃圾指数并非杜鹃群落多样性影响的至关因素, 但垃圾指数、踩踏指数确与杜鹃群落多样性及结构存在一定的相关性。Pearson相关分析表明单一垃圾指数和踩踏指数与杜鹃混交林物种多样性、群落生长结构相关性不强, 而对杜鹃纯林影响更为显著。从不同干扰强度来看, 杜鹃纯林和杜鹃混交林中度干扰时Berger-Parker丰富度指数和Shannon多样性指数最高, 这与吴甘霖[23]等对黄山松群落物种多样性研究中发现的中等干扰能一定程度上增加群落的物种多样性的结论相似, 与中度干扰假说(intermediate disturbance hypothesis)相符合, 即中等程度的干扰频率能维持高的物种多样性[24], 不同的是杜鹃纯林和杜鹃混交林多样性指数应对旅游干扰强度的变化趋势不同, 在杜鹃纯林中中度干扰>轻度干扰>重度干扰, 而在杜鹃混交林中中度干扰>重度干扰>轻度干扰。根据Tilman[25]关于群落多样性与稳定性的试验和理论研究指出:较高的生物多样性可以增加植物群落的生产力和生态系统的稳定性, 群落结构越复杂, 群落越稳定, 生态服务能力和自然更新能力越强, 抵御不良环境和外部干扰能力越强。很明显在两种杜鹃群落中, 杜鹃混交林物种更为丰富, 对于旅游干扰在物种多样性上表现更加稳定, 生物多样性较高, 受影响程度小, 抗干扰能力强。对于杜鹃纯林群落, 旅游干扰按轻、中、重排序时, Berger-Parker丰富度指数分别为1.505、4.613、1.494, Shannon多样性指数则是1.604、3.041、1.551, 可见物种多样性受干扰程度影响波动明显。
一些研究认为旅游干扰对乔木层的盖度没有影响, 草本层和灌木层影响较为明显[26-27]。但通过百里杜鹃样地乔木层分析, 重度干扰杜鹃样地平均盖度56%远低于中度干扰样地平均盖度85%, 原因可能在于杜鹃群落为低矮小乔木, 在乔木层郁闭度良好的杜鹃林分, 更易于吸引游客在林下休憩赏花, 从而导致土壤紧实, 又因杜鹃种子多以母株为中心扩散传播, 直接影响杜鹃种子发芽与更新[28], 进而影响杜鹃林乔木层物种多样性的变化。特别是杜鹃纯林乔木层物种多样性受旅游强度干扰负面影响大, Berger-Parker丰富度指数从中度干扰的1.772降至重度干扰的1.098, 多样性指数更从中度干扰的1.333降至重度干扰的0.392。
杜鹃纯林Shannon多样性指数、物种数以及草本均高都随垃圾指数增加而减少, 原因可能在于旅游者丢弃的塑料袋等固体废弃物, 能够改变土壤结构, 使杜鹃群落所喜酸性土壤PH值升高碱性增强, 进而导致杜鹃群落多样性指标降低[29-31]。土壤理化性质的改变也可使得土壤微生物的生长环境随之变化, 植物根系生长困难进而影响地上部分的生长。如三江源地区的研究表明, 退化草甸的土壤微生物及放线菌组成趋于简单、真菌种类数较天然草甸明显降低[32-33]。
杜鹃纯林踩踏指数与灌木均高、盖度呈现极显著负相关, 与草本均高呈显著正相关。杜鹃群落中踩踏干扰的介入既使上层土壤水含量等土壤理化性质均有所下降, 也打破了低矮灌木及草本植物原有的竞争格局, 使得光照、水分等条件重新分配, 一些耐踩踏的植物充分利用格局重组的机会占据有利位置, 通过调整再生产策略适应干扰的介入[33]。随踩踏干扰程度的增加, 杜鹃纯林中草本植物的优势种发生了很大变化, 在干扰程度较弱的样地以蕨类为主, 在干扰程度较强的样地则以椴树科扁担杆属植物堇菜、禾本科荩草属植物为主。相似的结论在很多研究中心均有报道, 如在香格里拉高寒草甸的研究表明随着踩踏程度的增加, 物种丰富度没有明显的变化, 但优势物种和主要伴生种的组合发生了很大变化, 在踩踏干扰较弱的样地以禾本科牧草为主要优势种, 在中高强度的样地中以禾本科和豆科植物为主要优势种[34]。
4 结论百里杜鹃景区不同杜鹃群落对于不同旅游干扰有着不同的响应, 从整体上说杜鹃混交林物种丰富, 抗干扰能力强于杜鹃纯林。根据调查样地分析, 旅游干扰对杜鹃群落存在一定的中度干扰效应, 即杜鹃纯林与杜鹃混交林同样在中度干扰时具有最高的Berger-Parker丰富度指数和Shannon多样性指数, 杜鹃群落物种多样性的增加在一定干扰程度内是生态系统自我调节的一种表现, 一旦超过其自我调节能力的阈值将很难恢复。正因为不同杜鹃群落对于不同旅游干扰有着不同的响应, 为了保持杜鹃群落的稳定性, 针对不同杜鹃群落应执行相应的管理制度。(1)对以露珠杜鹃为优势种的杜鹃纯林, 可在干扰严重地段设立木质围栏以便规范游客行为, 对于密度较大的露珠杜鹃纯林应适当调节, 开林窗, 以助露珠杜鹃形成自然的冠幅和林下更新。(2)杜鹃纯林应控制重度干扰, 在杜鹃花季适当分流, 合理配置旅游项目, 避免景点超负荷承载。(3)要实现动态监测, 加强旅游信息化建设, 建立网络监测系统, 减轻区域旅游环境承载压力。
| [1] |
Van der Duim R, Caalders J. Biodiversity and tourism: impacts and interventions. Annals of Tourism Research, 2002, 29(3): 743-761. DOI:10.1016/S0160-7383(01)00087-1 |
| [2] |
Kelly C L, Pickering C M, Buckley R C. Impacts of tourism on threatened plant taxa and communities in Australia. Ecological Management & Restoration, 2003, 4(1): 37-44. |
| [3] |
Pickering C M, Rossi S, Barros A. Assessing the impacts of mountain biking and hiking on subalpine grassland in Australia using an experimental protocol. Journal of Environmental Management, 2011, 92(12): 3049-3057. DOI:10.1016/j.jenvman.2011.07.016 |
| [4] |
Tzatzanis M, Wrbka T, Sauberer N. Landscape and vegetation responses to human impact in sandy coasts of Western Crete, Greece. Journal for Nature Conservation, 2003, 11(3): 187-195. DOI:10.1078/1617-1381-00047 |
| [5] |
Cole D N. Vegetational changes associated with recreational use and fire suppression in the Eagle Cap Wilderness, Oregon: Some management implications. Biological Conservation, 1981, 20(4): 247-270. DOI:10.1016/0006-3207(81)90013-6 |
| [6] |
Törn A, Tolvanen A, Norokorpi Y, Tervo R, Siikamäki P. Comparing the impacts of hiking, skiing and horse riding on trail and vegetation in different types of forest. Journal of Environmental Management, 2009, 90(3): 1427-1434. DOI:10.1016/j.jenvman.2008.08.014 |
| [7] |
Leung Y F, Marion J L. Recreation impacts and management in wilderness: a state-of-knowledge review//McCool S F, ed. Wilderness Science in a Time of Change Conference. Ogden, UT: USDA Forest Service, 2000: 23-48.
|
| [8] |
Monz C A, Cole D N, Leung Y F, Marion J L. Sustaining visitor use in protected areas: future opportunities in recreation ecology research based on the USA experience. Environmental Management, 2010, 45(3): 551-562. DOI:10.1007/s00267-009-9406-5 |
| [9] |
朱珠, 包维楷, 庞学勇, 闫晓丽, 李玉武. 旅游干扰对九寨沟冷杉林下植物种类组成及多样性的影响. 生物多样性, 2006, 14(4): 284-291. DOI:10.3321/j.issn:1005-0094.2006.04.002 |
| [10] |
陈翔, 黄家勇, 谢华, 陈训, 高贵龙. 百里杜鹃自然保护区杜鹃属植物资源研究. 贵州科学, 2010, 28(4): 26-34, 39-39. DOI:10.3969/j.issn.1003-6563.2010.04.005 |
| [11] |
贾真真, 李苇洁, 罗时琴, 李安定. 百里杜鹃风景名胜区季节性旅游影响分析. 安徽农业科学, 2012, 40(32): 15744-15745. DOI:10.3969/j.issn.0517-6611.2012.32.066 |
| [12] |
贾真真, 李苇洁, 李安定, 罗时琴. 百里杜鹃景区旅游对游径两侧植被的影响. 贵州科学, 2012, 30(5): 74-77. DOI:10.3969/j.issn.1003-6563.2012.05.017 |
| [13] |
王晓俊. 美国风景资源管理系统及其方法. 自然资源学报, 1993, 8(4): 371-380. DOI:10.3321/j.issn:1000-3037.1993.04.011 |
| [14] |
李贞, 保继刚, 覃朝锋. 旅游开发对丹霞山植被的影响研究. 地理学报, 1998, 53(6): 554-561. DOI:10.3321/j.issn:0375-5444.1998.06.011 |
| [15] |
程占红, 张金屯, 上官铁梁. 芦芽山自然保护区旅游开发与植被环境关系——旅游影响系数及指标分析. 生态学报, 2003, 23(4): 703-711. DOI:10.3321/j.issn:1000-0933.2003.04.011 |
| [16] |
方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. |
| [17] |
郝占庆, 于德永, 杨晓明, 丁之慧. 长白山北坡植物群落α多样性及其随海拔梯度的变化. 应用生态学报, 2002, 13(7): 785-789. DOI:10.3321/j.issn:1001-9332.2002.07.005 |
| [18] |
Zhang J, Xu B, Li M. Vegetation patterns and species diversity along elevational and disturbance gradients in the Baihua Mountain Reserve, Beijing, China. Mountain Research and Development, 2013, 33: 170-178. DOI:10.1659/MRD-JOURNAL-D-11-00042.1 |
| [19] |
吴倩楠, 董建文, 郑宇, 傅伟聪, 李慧, 朱志鹏, 陈梓茹, 丁国昌. 百里杜鹃国家森林公园优势种生态位研究. 南京林业大学学报: 自然科学版, 2017, 41(2): 175-180. |
| [20] |
李久林, 廖凤林. 百里杜鹃林马缨杜鹃种群结构和动态研究. 贵州科学, 1997, 15(1): 64-69. |
| [21] |
杨阳, 马立辉, 王海洋. 濒危植物树枫杜鹃(Rhododendron changii)种群结构及伴生群落特征. 生态学杂志, 2019, 38(5): 1352-1362. |
| [22] |
黄梅, 戴晓勇, 杨成华, 邓伦秀, 袁丛军, 李鹤, 龙海燕, 冯育才. 阔柄杜鹃典型植物群落的结构及其多样性特征分析. 西部林业科学, 2020, 49(2): 113-120, 136-136. |
| [23] |
吴甘霖, 黄敏毅, 段仁燕, 赵凯. 不同强度旅游干扰对黄山松群落物种多样性的影响. 生态学报, 2006, 26(12): 3924-3930. DOI:10.3321/j.issn:1000-0933.2006.12.002 |
| [24] |
Connell J H, Slatyer R O. Mechanisms of succession in natural communities and their role in community stability and organization. The American Naturalist, 1977, 111(982): 1119-1144. DOI:10.1086/283241 |
| [25] |
Tilman D, Downing J A. Biodiversity and stability in grasslands. Nature, 1994, 367(6461): 363-365. DOI:10.1038/367363a0 |
| [26] |
管东生, 丁键, 王林. 旅游和环境污染对广州城市公园森林植物和土壤的影响. 中国环境科学, 2000, 20(3): 277-280. DOI:10.3321/j.issn:1000-6923.2000.03.020 |
| [27] |
Weaver T, Dale D. Trampling effects of hikers, motorcycles and horses in meadows and forests. Journal of Applied Ecology, 1978, 15(2): 451-457. DOI:10.2307/2402604 |
| [28] |
李苇洁, 陈训. 百里杜鹃森林公园马缨杜鹃种群结构与更新初步研究. 贵州科学, 2005, 23(3): 46-49. DOI:10.3969/j.issn.1003-6563.2005.03.011 |
| [29] |
Bashir S, Goswami S. Tourism induced challenges in municipal solid waste management in hill towns: case of Pahalgam. Procedia Environmental Sciences, 2016, 35: 77-89. DOI:10.1016/j.proenv.2016.07.048 |
| [30] |
安登第, 何毅, 韩爱萍, 王晶. 不同利用方式对亚高山草甸草地土壤性质和微生物的影响. 草业科学, 2003, 20(6): 1-3. DOI:10.3969/j.issn.1001-0629.2003.06.001 |
| [31] |
王根绪, 沈永平, 钱鞠, 王军德. 高寒草地植被覆盖变化对土壤水分循环影响研究. 冰川冻土, 2003, 25(6): 653-659. DOI:10.3969/j.issn.1000-0240.2003.06.010 |
| [32] |
林超峰, 陈占全, 薛泉宏, 来航线, 陈来生, 张登山. 青海三江源区植被退化对土壤养分和微生物区系的影响. 应用与环境生物学报, 2007, 13(6): 788-793. DOI:10.3321/j.issn:1006-687x.2007.06.008 |
| [33] |
Müllerová J, Vítková M, Vítek O. The impacts of road and walking trails upon adjacent vegetation: effects of road building materials on species composition in a nutrient poor environment. Science of the Total Environment, 2011, 409(19): 3839-3849. DOI:10.1016/j.scitotenv.2011.06.056 |
| [34] |
张勇. 旅游踩踏对香格里拉高寒草甸植物群落的短期影响[D]. 昆明: 云南大学, 2013.
|