 2021, Vol. 41
2021, Vol. 41文章信息
- 盘远方, 李娇凤, 姚玉萍, 姜勇, 利恒春, 王晓凤, 卢国琼, 杨晨, 黄诗雯, 蒋文平
- PAN Yuanfang, LI Jiaofeng, YAO Yuping, JIANG Yong, LI Hengchun, WANG Xiaofeng, LU Guoqiong, YANG Chen, HUANG Shiwen, JIANG Wenping
- 桂林岩溶石山青冈群落植物功能多样性和环境因子与坡向的关联研究
- Changes in plant functional diversity and environmental factors of Cyclobalanopsis glauca community in response to slope gradient in Karst hills, Guilin
- 生态学报. 2021, 41(11): 4484-4492
- Acta Ecologica Sinica. 2021, 41(11): 4484-4492
- http://dx.doi.org/10.5846/stxb201906031169
-
文章历史
- 收稿日期: 2019-06-03
- 网络出版日期: 2021-04-06





2. 广西师范大学生命科学学院, 桂林 541006
2. College of Life Sciences, Guangxi Normal University, Guilin 541006, China
生物多样性与生态系统功能的关系是目前生态学领域备受关注的热点问题之一[1]。其中, 物种多样性是生物多样性在物种水平上的表现形式[2], 也是物种丰富度和分布均匀性的一个综合反映, 它可以直接或间接的体现了群落结构、群落类型、组织水平、发展阶段、稳定程度和生境差异等[3-4]。因此, 以往学者经常用物种多样性来代替生物多样性对生物多样性和生态系统功能关系进行研究, 但是物种多样性的指标比较简单难以真实地反映物种真实特性及其对生态系统的影响[2]。生态系统的功能关系不仅需要通过物种的数量来反映, 更需要考虑其内部每个物种所具有的不同的功能特性[5]。而功能多样性则充分考虑了生境内共存物种的互补和冗余特性[6], 从而把植物不同特征与生境因子紧密连接起来[7]。同时, 可利用多个性状对不同的生态系统功能进行描述[8]。因此与物种多样性相比, 功能多样性能更准确、真实的反映物种共存机制及生态系统过程[9-10]。
功能多样性是生物多样性一个重要的组成部分[7], 它是依据测定群落内部与生态系统功能关系密切的物种性状数值的分布范围作为研究生物多样性和生态系统功能关系的重要突破口[11]。目前, 许多学者围绕功能多样性在微生物、植物和动物群落中开展了广泛研究。例如, 不同林地凋落物组合对土壤速效养分和微生物群落功能多样性的影响[12];新薛河底栖动物物种多样性与功能多样性关系的研究[13];山西五鹿山森林群落木本植物功能多样性[10]等。环境因素可以作为一个“筛”, 它能够决定哪些物种或者性状可以在群落中生存和维持[14]。众多生态环境因子的组合可以产生不同的生境类型, 而生境异质性是物种多样性得以维持的一个重要因素[15]。研究表明, 地形变化是造成生境异质性的重要原因, 如坡向的不同导致太阳辐射和降水的空间二次分配, 伴随光辐射、光照时长、大气温度、土壤水分和土壤养分等环境因子的差异, 进而影响着植物群落类型、物种组成、物种多样性和功能多样性的变化[16]。如朱云云等研究发现植物功能多样性主要受坡向影响, 且功能丰富度在阴坡、半阴坡显著高于阳坡[17]。郑丽婷等研究发现功能均匀度与坡度呈正相关, 而功能分离度、功能离散度则更多地受土壤理化性质的影响, 即与土壤总碳呈正相关, 与土壤含水量呈负相关[18]。因此, 研究坡向和环境因子对植物功能多样性的影响, 可以更好地揭示植物对局部环境采取的适应策略的不同。
桂林位于广西壮族自治区东北部, 是典型的岩溶石山地貌, 岩溶发育强烈、景观复杂、生态脆弱, 植被遭到破坏极易退化且难以恢复。在桂林岩溶石山地区, 由于自然干扰和人为活动频繁等原因, 现存完整的原生植被非常少, 大多以灌丛或藤刺灌丛为主。以石漠化为典型特征的生态退化成为制约该地区经济发展的重大生态环境问题之一, 植被恢复成为桂林岩溶石山地区石漠化治理和生态重建的重要任务[19]。而青冈(Cyclobalanopsis glauca)为壳斗科(Fagaceae)青冈属(Cyclobalanopsis)常绿大乔木, 是我国南方亚热带岩溶石山地区常绿落叶阔叶混交林的主要成分之一, 具有较强的石生性、耐旱性, 是岩溶生态系统的顶级群落的建群种, 对维持岩溶生态系统稳定起着至关重要的作用[20-21]。近年来, 基于植物功能多样性和功能性状特征的研究方法在生态领域中受到重视, 但大多的研究主要集中在植物功能性状与环境因子相关性等方面, 而关于地形、环境因子对群落植物功能多样性指数影响的研究鲜见报道。因此, 本文以桂林岩溶石山不同坡向上的青冈群落为研究对象, 同时选取2个与群落系统功能存在密切关系的功能性状(比叶面积、木材密度)计算功能多样性指数。通过分析青冈群落植物功能多样性指数和环境因子(岩石裸露率、土壤全磷和土壤有效磷含量)在不同坡向上的变化规律, 探讨不同坡向环境因子对青冈群落功能多样性的筛选效应, 阐释岩溶石山青冈群落的构建机制, 以期为岩溶石山植被恢复重建及生态系统服务功能评价提供科学依据。
1 材料与方法 1.1 研究区概况研究区位于广西壮族自治区桂林市(24°40′N—25°40′N, 110°9′E—110°42′E), 桂林为典型的岩溶石山地貌, 海拔范围在100—500 m。该地区地貌特征明显, 石峰、石林等生态景观类型多样。该地区属中亚热带湿润季风气候, 气候温和, 雨量充沛, 年平均降雨量1 949.5 mm, 降雨量年分配不均, 雨季主要集中在4—7月份, 年均蒸发量为1 690—1 905 mm, 年平均气温19 ℃, 全年无霜期309 d, 全年以偏北风为主, 风速为2.2—2.7 m/s。研究区主要以石灰岩、白云岩等碳酸盐类岩石为主的岩溶石山, 具有典型的岩溶石山特征(岩石裸露率高、环境容纳量低、成土速度慢、土壤层浅薄、土壤钙含量高、土壤蓄水能力差)。受其特殊的生境条件制约, 其适生的植物均具有较强的耐旱性、石生性、嗜钙性等特点。研究区主要优势种为青冈(Cyclobalanopsis glauca), 而伴生种则有红背山麻杆(Alchornea trewioides)、山麻杆(Alchornea davidii)、菜豆树(Radermachera sinica)、粗糠柴(Mallotus philippensis)、岩樟(Cinnamomum saxatile)、胡颓子(Elaeagnus pungens)等。
1.2 样地设置与群落学调查在对桂林岩溶石山植被类型、分布情况等做系统踏查的基础上, 结合青冈群落的类型、结构以及生境等特点, 于2017年7—9月选取位于桂林市郊的演坡山和张家村以及阳朔县白沙镇和高田镇碑头村4处受人为活动最少、原生性较强且发育完好的青冈群落为研究对象。分别在阴坡(碑头)、阳坡(张家村、演坡山和白沙镇)各设置7个20 m×20 m的样方, 共14个样方, 各样方的基本概况如表 1所示。先将每个20 m×20 m的样方划分为4个10 m×10 m的小样方。调查时, 记录每个小样方内胸径≥1 cm的所有木本植物的种名、树高、基径、胸径、空间坐标与存活状况。利用精度为0.01的可触式智能双星导航仪(Touch35)获取海拔、坡向、坡度、经纬度等数据, 同时记录每个10 m×10 m小样方内的岩石裸露率和调落物厚度等环境特征。
| 样方编号 Sample number |
采样点 Sampling locations |
经纬度 Latitude and longitude |
海拔/m Altitude |
岩石裸露率/% Rock exposure rate |
坡向 Slope aspect |
| P1 | 碑头 | 24°44′42.93″ N 110°25′19.87″ E | 173 | 56 | 阴坡 |
| P2 | 碑头 | 24°44′43.44″ N 110°25′18.84″ E | 188 | 50 | 阴坡 |
| P3 | 碑头 | 24°44′43.67″ N 110°25′19.56″ E | 187 | 42 | 阴坡 |
| P4 | 碑头 | 24°44′43.40″ N 110°25′19.71″ E | 198 | 42 | 阴坡 |
| P5 | 碑头 | 24°44′42.78″ N 110°25′19.87″ E | 198 | 36 | 阴坡 |
| P6 | 碑头 | 24°44′42.46″ N 110°25′22.30″ E | 198 | 48 | 阴坡 |
| P7 | 碑头 | 24°44′42.42″ N 110°25′22.32″ E | 208 | 53 | 阴坡 |
| P8 | 芦笛岩张家村 | 25°18′19.44″ N 110°15′48.35″ E | 179 | 72 | 阳坡 |
| P9 | 演坡山 | 25°19′10.08″ N 110°15′19.06″ E | 234 | 51 | 阳坡 |
| P10 | 演坡山 | 25°19′9.35″ N 110°15′19.26″ E | 237 | 54 | 阳坡 |
| P11 | 演坡山 | 25°19′08.67″ N 110°15′18.82″ E | 237 | 76 | 阳坡 |
| P12 | 演坡山 | 25°19′08.32″ N 110°15′17.85″ E | 240 | 67 | 阳坡 |
| P13 | 白沙镇 | 24°49′53.67″ N 110°22′48.27″ E | 268 | 58 | 阳坡 |
| P14 | 白沙镇 | 24°49′52.10″ N 110°22′44.17″ E | 244 | 45 | 阳坡 |
土壤采样是以10 m×10 m小样方为基本单位, 采用“梅花五点法”[22], 在每个10 m×10 m小样方的4个顶点处和中心位置, 清除表层调落物和石块后, 用直径10 cm的土钻钻取0—20 cm土层土样混合均匀带回实验室用于土壤养分的测定。将野外采回的土样放在室内通风处自然风干, 风干后剔除土样中掺杂的根系、石块、钙核及动植物残体等杂物后倒入研钵中研细, 过不同孔径(0.15 mm、0.25 mm、1.4 mm)的土壤筛, 对预处理后的土样进行土壤全磷、土壤有效磷等指标的测定。其中, 土壤全磷含量(TP, g/kg)采用NaOH碱熔-钼锑抗比色法测定[23];土壤有效磷含量(AP, mg/kg)采用双酸浸提紫外可见分光光度比色法测定[23]。
1.4 植物功能性状取样与测定植物功能性状测定主要包括样方内胸径≥1 cm的木本植物植株个体的比叶面积和木材密度等指标。每个植株个体选取接近冠层3片完全展开且健康成熟的阳叶及3根长10 cm左右直径为1—2 cm的非当年生枝条放入密封袋内, 置于阴暗环境下并迅速带回实验室进行叶和茎功能性状测定。叶性状的测定, 利用叶面积仪(Yaxin-1241, 北京雅欣理仪科技有限公司)扫描每片叶片的叶面积, 然后将叶片样品放入有对应编号的信封内后置于80 ℃烘箱中烘干至恒重后, 称量并记录其叶片干重。对于木材密度的测定, 本文利用测定的枝条密度代替木材密度, 将枝条削去表面的树皮后, 使用电子天平测定其鲜重, 接着用量筒排水法测定枝条的体积, 然后将其放入有对应编号的信封内并置于80 ℃烘箱中烘干至恒重后, 测定枝条干重。本次采集了6528个植株个体, 并测定了19584个叶片性状和19584个枝条性状。
1.5 数据处理功能多样性指数计算是利用比叶面积和木材密度这两个与群落生态系统功能关系存在密切相关的性状为基础进行多度加权计算。功能多样性指数计算公式如下[24-26]:
功能丰富度(FRci)

|
(1) |
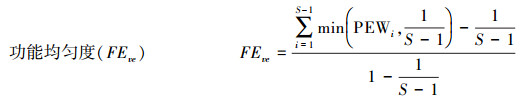
|
(2) |

|
(3) |
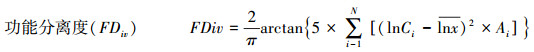
|
(4) |
式中, FRci为群落i中性状c的功能丰富度, SFci为群落中物种所占据的生态位空间, Rc为所有群落中性状c占据生态为空间。S为物种丰富度, PEWi为物种i的局部加权均匀度。aj是j物种的相对多度, zj是物种j到加权质心的距离。FDiv为包含多项功能特征的功能分离度指数, Ci为第i项功能特征的数值, lnx为物种特征值的自然对数, Ai为第i项功能特征的丰度比例, N为群落中的物种数。
首先, 不同坡向(阴坡、阳坡)上植物功能多样性指数与环境因子的差异性分析采用Wilcoxon加符秩检验。其次, 探究不同坡向上植物功能多样性指数与环境因子的相关性采用简单线性回归模型分析, 即以植物功能多样性指数作为因变量, 以环境因子作为自变量进行回归分析。最后, 根据R2和对模型判断P的统计检验得出不同坡向上对植物功能多样性指数影响显著的环境因子, 并做相关关系趋势图(即散点图)。所有统计分析和绘图均在R 3.5.2软件(The R Development Core Team, 2018)中完成。
2 结果与分析 2.1 不同坡向植物功能多样性的比较如图 1所示, 功能丰富度、功能均匀度、功能离散度和功能分离度对阴坡、阳坡生境均产生显著的响应(P<0.05)。且功能丰富度、功能均匀度、功能离散度和功能分离度在不同坡向上均表现出相同的变化趋势, 即表现为阴坡显著大于阳坡。

|
| 图 1 不同坡向功能多样性指数的比较 Fig. 1 Compare different slope aspect of functional diversity index FRic:功能丰富度Functional richness;FEve:功能均匀度Functional evenness;FDis:功能离散度Functional dispersion;FDiv:功能分离度Functional divergence; a和b表示阴坡与阳坡差异性显著 |
如图 2所示, 岩石裸露率、土壤全磷含量、土壤有效磷含量在阴坡、阳坡上均表现为显著差异性(P<0.05);其中岩石裸露率和土壤有效磷含量表现为相同的变化趋势, 即阴坡小于阳坡;而土壤全磷含量则为阴坡大于阳坡。

|
| 图 2 不同坡向环境因子的比较 Fig. 2 Comparison of different slope aspect environmental factors RER:岩石裸露率Rock exposure rate;TP:土壤全磷Soil total phosphorus;AP:土壤有效磷Soil available phosphorus |
多元逐步回归分析结果表明, 在阴坡上, 功能离散度、功能分离度与岩石裸露率呈显著负相关(P<0.05, R2=0.606, R2=0.501);而功能离散度、功能分离度与岩石裸露率在阳坡上差异性不显著(P>0.05)(图 3)。在阳坡上, 功能均匀度与土壤全磷含量呈显著正相关(P<0.05, R2=0.626);功能分离度与土壤有效磷含量呈显著正相关(P<0.05, R2=0.574), 但功能均匀度与土壤全磷含量、功能分离度与土壤有效磷含量在阴坡上相关性不显著(P>0.05)(图 3)。而功能丰富度、功能均匀度与岩石裸露率在阴坡、阳坡相关性不显著(P>0.05)(图 3)。功能丰富度、功能离散度和功能分离度与土壤全磷含量在阴坡、阳坡相关性不显著(P>0.05);功能丰富度、功能均匀度和功能离散度与土壤有效磷含量在阴坡、阳坡相关性也不显著(P>0.05)(图 3)。

|
| 图 3 不同坡向功能多样性指数与环境因子的回归分析 Fig. 3 Regression analysis of different slope aspect between functional diversity index and environmental factors 实线表示阴坡;虚线表示阳坡 |
功能丰富度指数是指对多少生态位空间被现有物种所占据性状空间的面积或体积的测量[27]。所以在某种程度上, 功能丰富度指数与物种丰富度指数具有正相关性, 即功能多样性随着物种丰富度的升高而升高[28-29]。本研究结果显示功能丰富度阳坡显著低于阴坡。而功能丰富度指数可作为潜在生态位空间的指示使用, 即当物种的性状呈随机分布时, 其物种越丰富, 那么物种所占据的性状空间生态位也就越大[24]。低的功能丰富度表示群落内的物种种类不够丰富, 资源没有完全被充分利用即生态位空间还有剩余, 我们研究结果发现功能丰富度阴坡显著大于阳坡, 暗示植物功能丰富度对坡向这个环境因子具有明显的响应, 这说明了生长在阳坡上的植物生长发育受到了坡向的制约, 从而使植物没能充分利用生态空间[26]。功能均匀度指数是指物种性状平均值在己占据性状空间中是否分布均匀的度量[25]。当性状空间内的物种及其多度都均匀时, 功能均匀度为最高。反之, 则代表群落中物种和其丰富度在性状空间内呈分散集群状态[24], 即物种或性状存在间隙分布, 这会导致生产力和稳定性减小, 增加入侵机会[8, 24]。本研究结果显示, 功能均匀度阴坡显著大于阳坡, 意味着阴坡植物的功能性状在生态位空间的分布更加均匀。一般阳坡生境, 群落内光辐射强度大, 群落内主要以喜阳的物种为主, 且种类和分布格局均没有阴坡物种丰富和分布均匀, 同时这些物种具有相近的性状(如较低的比叶面积)来适应光因子的环境筛选作用[18], 可能会产生很高的功能冗余。功能离散度指数就是指群落资源差异程度、竞争程度及群落中极端物种的优势度的一个综合衡量[24], 它是通过相对多度来计算各物种间的距离, 但是仅对加权后的功能性状的平均绝对偏差进行度量, 所以功能离散度指数不受物种丰富度的影响。具有高功能离散度的群落会增加生态系统功能, 更有效的资源利用, 低功能离散度则暗示着这些区域生态位分化程度较低, 资源利用不充分, 资源竞争剧烈[24]。本研究结果显示阳坡的功能离散度显著低于阴坡, 这说明阴坡的生态位分化程度高于阳坡且资源利用较充分, 资源竞争强度较弱, 而阳坡植物的生存则要面临更强的资源竞争。功能分离度体现了群落中物种间生态位的互补程度[25]。生态系统中的功能分离度高说明物种生态位重叠效应弱, 生态位分化程度高, 由此产生较低的资源竞争。在我们研究中发现阴坡的植物功能分离度显著高于阳坡。这说明同阳坡比, 阴坡生境资源丰富, 物种种类丰富, 而较高的物种丰富度也增加了功能性状的数值范围, 从而提高了阴坡的功能离散度[9]。
3.2 环境因子在不同坡向上的变化规律岩溶石山典型的生境特点是峰丛地貌, 地形破碎, 土层浅薄且不连续, 具有明显的坡向变化。而坡向作为重要的地形因子, 是影响土壤养分、水分和太阳辐射重新再分配的间接环境因子之一。不同的坡向会导致所接受的太阳辐射量差异性大, 进而导致了土壤水分、温度、湿度、有机质、氮磷含量等在不同坡向上分布不均[30-31]。本研究表明, 环境因子(岩石裸露率、土壤全磷含量和土壤有效磷含量)在阴坡、阳坡上均存在显著差异。其中, 阳坡的岩石裸露率显著大于阴坡, 这与刘旻霞等研究结果一致[32], 阳坡接受到的太阳辐射总量较阴坡大, 导致土壤表层温度升高、土壤水分蒸发量大, 土壤含水量减少, 阳坡土壤风化严重, 加上岩溶石山本身石漠化严重, 土层浅薄且不连续, 水分容易漏失, 导致阳坡岩石裸露率较阴坡高。土壤养分作为土壤肥力的重要标志, 可直接影响到植物的生长和生物量的形成。而土壤磷素是植物生长发育所必需的重要元素之一, 磷素在土壤中的含量受土壤母质、成土作用以及外界因素的影响[33]。本研究结果显示, 土壤全磷含量阴坡高于阳坡, 土壤有效磷含量为阳坡显著高于阴坡(图 2)。这与盘远方对桂林岩溶石山灌丛植物叶功能性状和土壤因子对坡向的响应研究中得出土壤TP含量在阴坡显著大于阳坡的结论具有一致性, 表明阴坡具有相对较高的土壤营养资源[34]。但与土壤AP含量在阳坡小于阴坡的研究结论相悖。这可能与阳坡接受太阳辐射较多, 土温较高, 有利于微生物活动, 因而本研究中阳坡的土壤有效磷含量较高[17]。
3.3 不同坡向上植物功能多样性指数与环境因子的回归分析地形因子的作用在于影响物质和能量的再分配, 地形是多种环境因子的复合, 水分, 温度和土壤养分的变化集中体现在地形梯度的变化上[35]。其中, 坡向通过影响地表接受的太阳辐射量的不同导致其所在地段的温度、湿度、水分条件必然也会有一定的差异。这种环境条件的差异必然会影响植物功能性状的表型可塑性。那么植物的生长发育不仅取决于自身的生理特性, 也受环境条件的影响。对于功能多样性而言, 是以物种性状特征为基础, 通过多度加全计算得出数值的高低来反映群落结构不同方面的功能特征[36]。功能多样性指标包含功能丰富度、功能均匀度、功能离散度、功能分离度四个指数, 这些指数可表现功能多样性不同方面的特征[24]。本研究结果显示功能均匀度指数与土壤全磷在阳坡上呈显著的正相关(图 3)。全磷是土壤中主要的限制因子, 也就是说, 土壤磷的含量越丰富, 物种或性状的分布间隙越小越均匀。同时功能性状在多维生态位空间上分布的更具有规律性, 物种对土壤养分等资源利用的更充分。功能离散度与岩石裸露率在阴坡上呈显著的负相关(图 3)。功能离散度描述物种功能和性状空间物种簇(若干物种聚集在一起)所处位置的差异程度, 可作为群落资源差异程度, 乃至竞争程度的指标。在本研究中发现阳坡岩石裸露率显著高于阴坡, 暗示阳坡的土壤水分流失程度高, 土壤养分缺乏, 资源贫瘠。相对于阳坡, 阴坡的生境岩石裸露率低, 水分和土壤养分资源较丰富, 因而决定其植被组成较阳坡丰富, 以致各物种性状叠加后整体群落的功能性状更加丰富, 意味着群落中的各部分资源被有效利用, 生态位分化程度高, 竞争能力和抗干扰能力强。因此, 在阴坡岩石裸露率与功能离散度指数呈负相关。功能分离度与斑块大小成显著负相关, 小斑块有更高的分离度[27]。本研究发现在阴坡上功能分离度与岩石裸露率呈显著的负相关(图 3), 印证了阴坡具有更高的功能分离度。此外, 在阳坡上, 功能分离度与土壤有效磷呈显著的正相关(图 3)。有效磷是指土壤中可被植物吸收利用的磷的总称, 其含量影响植物的生长状况, 土壤中有效磷含量越高, 则磷在植物体内参与的光合作用、呼吸作用、能量储存和传递等生理代谢能力越强, 促进阳坡植物根系的形成和生长, 以便扎根至深层土壤来吸收水源, 提高了植物适应外界干旱环境条件的能力[17]。由于阳坡土壤裸露、风化严重等较恶劣的环境条件会影响生态系统的生物量及可获取的资源数量, 因而植物需要调整其生理性状来适应其生存环境以最大化地获取资源, 降低生态位重叠程度和竞争强度。因此, 功能分离度与土壤有效磷呈显著的正相关。
4 结论坡向影响地表接受的太阳辐射量, 而太阳辐射是决定土壤营养循环的主要因子。由于不同坡向光、热、水、土等自然因素的差异, 植被的空间分布格局也不同。本研究结果显示不同坡向环境因子与群落植物功能多样性指数存在显著的差异并且群落植物功能多样性指数受到坡向、岩石裸露率、土壤全磷含量、土壤有效磷含量等共同作用。植物的生长发育会受到其生境的影响, 尽管不同坡向上的植物生长发育会受到土壤养分资源和岩石裸露率的限制, 但植物功能性状会采取不同的生存策略来适应其生境。本文揭示了青冈群落植物功能性状多样性和环境因子对不同坡向的响应规律, 旨在探讨岩溶地区强烈的环境异质性的驱使下优势种群青冈的生理生态适应对策。岩溶石山是我国生态环境脆弱区, 森林植被的恢复是其石漠化治理和生态环境建设的首要任务, 而封山育林是岩溶地区森林植被自然恢复的重要途径之一。在自然封育过程中对于郁闭度较高的天然林进行适度的人为干扰, 通过间伐、砍灌、清理林下活物等抚育措施, 建立小面积林窗, 使青冈更新层能够接受更多的光照, 以便为更新层个体的发育创造有利生境, 提高乔木幼苗存活率及物种多样性。另一方面, 应积极宣传, 提高民众对青冈种群的科学认知, 加强对青冈种群的保护, 阻止人为砍伐导致青冈种群数量和分布面积的锐减, 从而保护青冈种群的持续稳定发展。
| [1] |
李瑞新, 丁勇, 马文静, 牛建明, 张庆. 植物功能多样性及其与生态系统功能关系研究进展. 生态环境学报, 2016, 25(6): 1069-1075. |
| [2] |
Mason N W H, Richardson S J, Peltzer D A, De Bello F, Wardle D A, Allen R B. Changes in coexistence mechanisms along a long-term soil chronosequence revealed by functional trait diversity. Journal of Ecology, 2012, 100(3): 678-689. DOI:10.1111/j.1365-2745.2012.01965.x |
| [3] |
田平, 程小琴, 韩海荣, 周文嵩. 环境因子对山西太岳山典型森林类型物种多样性及其功能多样性的影响. 西北植物学报, 2017, 37(5): 992-1003. |
| [4] |
彭羽, 米凯, 卿凤婷, 冯金朝, 薛达元. 影响植被退化生态因子的多尺度分析——以和林县为例. 应用基础与工程科学学报, 2015, 23(S1): 11-19. |
| [5] |
Biswas S R, Mallik A U, Braithwaite N T, Biswas P L. Effects of disturbance type and microhabitat on species and functional diversity relationship in stream-bank plant communities. Forest Ecology and Management, 2019, 432: 812-822. DOI:10.1016/j.foreco.2018.10.021 |
| [6] |
Niu K C, Choler P, De Bello F, Mirotchnick N, Du G Z, Sun S C. Fertilization decreases species diversity but increases functional diversity: A three-year experiment in a Tibetan alpine meadow. Agriculture, Ecosystems & Environment, 2014, 182: 106-112. |
| [7] |
Zhang J T, Zhang B, Qian Z Y. Functional diversity of Cercidiphyllum japonicum, communities in the Shennongjia Reserve, central China. Journal of Forestry Research, 2015, 26(1): 171-177. DOI:10.1007/s11676-014-0010-z |
| [8] |
Chen D M, Cheng J H, Chu P F, Mi J, Hu S J, Xie Y C, Tuvshintogtokh I, Bai Y F. Effect of diversity on biomass across grasslands on the Mongolian Plateau: Contrasting effects between plants and soil nematodes. Journal of Biogeography, 2016, 43(5): 955-966. DOI:10.1111/jbi.12683 |
| [9] |
Mouchet M A, Villéger S, Mason N W H, Mouillot D. Functional diversity measures: an overview of their redundancy and their ability to discriminate community assembly rules. Functional Ecology, 2010, 24(4): 867-876. DOI:10.1111/j.1365-2435.2010.01695.x |
| [10] |
薛倩妮, 闫明, 毕润成. 山西五鹿山森林群落木本植物功能多样性. 生态学报, 2015, 35(21): 7023-7032. |
| [11] |
Ali A, Yan E R, Chang S X, Cheng J Y, Liu X Y. Community-weighted mean of leaf traits and divergence of wood traits predict aboveground biomass in secondary subtropical forests. Science of the Total Environment, 2017, 574: 654-662. DOI:10.1016/j.scitotenv.2016.09.022 |
| [12] |
宋蒙亚, 李忠佩, 刘明, 刘满强, 江春玉. 不同林地凋落物组合对土壤速效养分和微生物群落功能多样性的影响. 生态学杂志, 2014, 33(9): 2454-2461. |
| [13] |
陈静, 蒋万祥, 贺诗水, 王洪凯, 卓丽玲, 陈青, 王红妹. 新薛河底栖动物物种多样性与功能多样性研究. 生态学报, 2018, 38(9): 3328-3336. |
| [14] |
Dunck B, Algarte V M, Cianciaruso M V, Rodrigues L. Functional diversity and trait-environment relationships of periphytic algae in subtropical floodplain lakes. Ecological Indicators, 2016, 67: 257-266. DOI:10.1016/j.ecolind.2016.02.060 |
| [15] |
赵振勇, 王让会, 尹传华, 张慧芝, 王雷. 天山南麓山前平原植物群落物种多样性及空间分异研究. 西北植物学报, 2007, 27(4): 4784-4790. |
| [16] |
段贝贝, 赵成章, 徐婷, 郑慧玲, 冯威, 韩玲. 兰州北山不同坡向刺槐叶脉密度与气孔性状的关联性分析. 植物生态学报, 2016, 40(12): 1289-1297. DOI:10.17521/cjpe.2016.0215 |
| [17] |
朱云云, 王孝安, 王贤, 邓美皎. 坡向因子对黄土高原草地群落功能多样性的影响. 生态学报, 2016, 36(21): 6823-6833. |
| [18] |
郑丽婷, 苏田, 刘翔宇, 尹芳, 郭超, 妥彬, 阎恩荣. 庙岛群岛典型植物群落物种、功能、结构多样性及其对环境因子的响应. 应用生态学报, 2018, 29(2): 343-351. |
| [19] |
Gazol A, Camarero J J. Functional diversity enhances silver fir growth resilience to an extreme drought. Journal of Ecology, 2016, 104: 1063-1075. DOI:10.1111/1365-2745.12575 |
| [20] |
胡刚, 梁士楚, 张忠华, 谢强. 桂林岩溶石山青冈栎群落的数量分析. 生态学杂志, 2007, 26(8): 1177-1181. |
| [21] |
胡满, 曾思齐, 龙时胜, 仇建友, 俞军剑. 青冈栎次生林种群结构及动态特征. 中南林业科技大学学报, 2017, 37(11): 110-114. |
| [22] |
卜文圣, 臧润国, 丁易, 张俊艳, 阮云泽. 海南岛热带低地雨林群落水平植物功能性状与环境因子相关性随演替阶段的变化. 生物多样性, 2013, 21(3): 278-287. |
| [23] |
鲍士旦. 土壤农化分析 (第三版). 北京: 中国农业出版社, 2000: 76-79.
|
| [24] |
Mason N W H, Mouillot D, Lee W G, Wilson J B. Functional richness, functional evenness and functional divergence: the primary components of functional diversity. Oikos, 2005, 111(1): 112-118. DOI:10.1111/j.0030-1299.2005.13886.x |
| [25] |
Villéger S, Mason N W H, Mouillot D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology, 2008, 89(8): 2290-2301. DOI:10.1890/07-1206.1 |
| [26] |
Laliberté E. Legendre PA distance-based framework for measuring functional diversity from multiple traits. Ecology, 2010, 91(1): 299-305. DOI:10.1890/08-2244.1 |
| [27] |
路兴慧, 臧润国, 丁易, 黄继红, 杨秀森, 周亚东. 抚育措施对热带次生林群落植物功能性状和功能多样性的影响. 生物多样性, 2015, 23(1): 79-88. |
| [28] |
Bu W S, Zang R G, Ding Y. Functional diversity increases with species diversity along successional gradient in a secondary tropical lowland rainforest. Tropical Ecology, 2014, 55(3): 393-401. |
| [29] |
Ruiz-Benito P, Gómez-Aparicio L, Paquette A, Messier C, Kattge J, Zavala M A. Diversity increases carbon storage and tree productivity in Spanish forests. Global Ecology and Biogeography, 2014, 23(3): 311-322. DOI:10.1111/geb.12126 |
| [30] |
刘旻霞, 马建祖. 甘南高寒草甸植物功能性状和土壤因子对坡向的响应. 应用生态学报, 2012, 23(12): 3295-3300. |
| [31] |
方精云, 沈泽昊, 崔海亭. 试论山地的生态特征及山地生态学的研究内容. 生物多样性, 2004, 12(1): 10-19. DOI:10.3321/j.issn:1005-0094.2004.01.003 |
| [32] |
刘旻霞, 王刚. 高寒草甸植物群落多样性及土壤因子对坡向的响应. 生态学杂志, 2013, 32(2): 259-265. |
| [33] |
赵少华, 宇万太, 张璐, 沈善敏, 马强. 土壤有机磷研究进展. 应用生态学报, 2004, 15(11): 2189-2194. DOI:10.3321/j.issn:1001-9332.2004.11.041 |
| [34] |
盘远方, 陈兴彬, 姜勇, 梁士楚, 陆志任, 黄宇欣, 倪鸣源, 覃彩丽, 刘润红. 桂林岩溶石山灌丛植物叶功能性状和土壤因子对坡向的响应. 生态学报, 2018, 38(5): 1581-1589. |
| [35] |
董世魁, 汤琳, 张相锋, 刘世梁, 刘全儒, 苏旭坤, 张勇, 武晓宇, 赵珍珍, 李钰, 沙威. 高寒草地植物物种多样性与功能多样性的关系. 生态学报, 2017, 37(05): 1472-1483. |
| [36] |
许格希, 史作民, 唐敬超, 刘顺, 马凡强, 许涵, 刘世荣, 李意德. 海南尖峰岭热带山地雨林林冠层树种功能多样性特征. 应用生态学报, 2016, 27(11): 3444-3454. |