 2021, Vol. 41
2021, Vol. 41文章信息
- 王瑶, 张利敏, 徐胜楠, 于鸿淼, 刘月
- WANG Yao, ZHANG Limin, XU Shengnan, YU Hongmiao, LIU Yue
- 帽儿山3种森林生态系统土壤动物与土壤呼吸及其相互关系研究
- Analysis on soil animals, soil respiration and the correlation in three forest ecosystems in Maoershan
- 生态学报. 2021, 41(1): 172-183
- Acta Ecologica Sinica. 2021, 41(1): 172-183
- http://dx.doi.org/10.5846/stxb201911252555
-
文章历史
- 收稿日期: 2019-11-25
- 网络出版日期: 2020-11-19





目前, CO2浓度升高导致的全球气候变暖等负面效应已成为当前人类生存最主要面临的生态环境问题, 土壤呼吸作为向大气中释放CO2最大的源之一, 成为社会各界关注的重点[1-2]。土壤呼吸是土壤释放CO2的过程, 主要包括4个部分, 分别为3个生物学过程(植物根系呼吸、土壤微生物呼吸和土壤动物呼吸)和1个土壤化学氧化过程[3], 其中土壤动物呼吸占土壤总呼吸的5%—10%[4]。虽然土壤动物直接释放的CO2在整个土壤呼吸中所占的比例并不高, 但是土壤动物通过土壤碎屑食物网对碳释放的间接调控作用是不容忽视的。土壤动物是生存在土壤中或者落叶下的, 对土壤有着一定影响的动物的总称[5], 它分布广泛, 种类繁多, 是生态系统中物质循环的重要的消费者[6]。土壤动物一方面可以通过其自身取食、分泌、代谢等活动直接释放CO2, 另一方面土壤动物对微生物、植物根系的取食及自身排泄作用可影响微生物生命活动及根系周转, 同时参与凋落物及有机质的分解从而对土壤呼吸产生间接影响[7]。有研究表明作为土壤碎屑食物网的较高营养级, 土壤动物占优势时增加了可利用的养分来调节土壤呼吸从而固定更多的碳[8], 因此土壤动物在调控CO2释放的生态过程中发挥着巨大的作用。由于土壤动物在土壤生态系统中具有重要地位, 而目前的大多数学者都主要采用室内培养实验[9-10]的研究方法, 且研究集中在单个或几个物种对于碳循环的影响机理, 例如有研究发现单独添加蚯蚓[9]或线虫[10]均会增加土壤CO2的排放通量, 还有研究发现蚂蚁巢穴周围CO2的排放量要高于周边土壤[11], 但是目前采用野外控制实验且对土壤动物群落的关注较少, 因此探讨有关土壤动物群落与土壤碳循环的关系问题逐渐成为研究热点[12-14]。
作为陆地生态系统的主体, 森林生态系统的土壤碳库是陆地生态系统碳库的重要组成部分[15], 约占全球土壤碳库73%[16], 而温带森林是碳循环的敏感区域[17]。因此, 本研究以帽儿山森林生态系统观测研究站为研究区, 选择3种水热状况不同的森林生态系统, 通过野外定点实验与室内分析, 探讨不同森林生态系统生长季土壤呼吸随时间的变化规律及其与土壤动物生物量和个体密度之间的相互关系。已有研究表明土壤温度、土壤含水率是影响土壤呼吸以及土壤动物群落结构的重要因素, 我们推测:由于土壤温度和土壤含水率有明显的时间变化, 因此土壤呼吸及土壤动物群落结构也应该具有显著的时间变化, 且土壤动物群落结构的变化会对土壤呼吸, 尤其是土壤异养呼吸产生显著影响。
1 材料与方法 1.1 研究区概况本研究样地位于黑龙江帽儿山森林生态系统观测研究站, 地理坐标为45°24′ N, 127°28′ E, 平均海拔约400 m。该区属典型大陆性温带季风气候, 冬季漫长而干冷, 夏季短促而湿热, 降水主要集中在7—8月份, 年平均降水量723 mm, 年平均蒸发量1093 mm, 年平均气温2.7 ℃, 年平均总日照时数1856.8 h, 无霜期约为120—140 d。地带性土壤为暗棕壤。现有的主要植被类型是经过不同程度的人为干扰而形成的典型的东北东部山区天然次生林。
1.2 样地布设选择帽儿山森林生态站内同一地点的水热条件不同的3种森林生态系统, 分别为红松人工林、蒙古栎林和硬阔叶林进行样地布设。3种森林生态系统植被组成丰富, 乔木树种共有9种, 下木共有9种。其中, 红松人工林乔木最多(7种), 分别为红松Pinus koraiensis Siebold et Zuccarini(优势种)、白桦Betula platyphylla Suk.、水曲柳Fraxinus mandschurica Rupr.、枫桦Betula costata Trautv.、山杨Populus davidiana Dode、胡桃楸Juglans mandshurica Maxim.和黄檗Phellodendron amurense Rupr., 下木仅有春榆Ulmus davidiana Planch var. japonica (Rehd.)Nakai和山梨Pyrus ussuriensis Maxim. 2种;蒙古栎林次之, 乔木6种, 分别为蒙古栎Quercus mongolica Fischer ex Ledebour(优势种)、紫椴Tilia amurensis Rupr.、水曲柳F. mandshurica、黄檗P. amurense、山杨P. davidiana、白桦B. platyphylla, 下木有暴马丁香Syringa reticulata (Blume) H. Hara var. amurensis (Rupr.) J. S. Pringle、色木槭Acer mono Maxim.、春榆U. propinqua、鼠李Rhamnus davurica Pall. 4种;硬阔叶林乔木最少, 仅黄檗P. amurense、水曲柳F. mandshurica、紫椴T. amurensis、胡桃楸J. mandshurica 4种, 下木最多, 分别为暴马丁香Syringa amurensis、春榆U. propinqua、榛Corylus heterophylla Fisch. ex Trautv.、稠李Padus racemosa (Lam.) Gilib.、早花忍冬Lonicera praeflorens Batalin、卫矛Euonymus alatus (Thunb.) Sieb. 6种[18-19]。3块样地的立地状况及土壤性质如表 1, 表 2。
| 林型 Forest type |
AN/(mg/kg) | AP/(mg/kg) | AK/(mg/kg) | DOC/(mg/kg) | SWC/% |
| HS | 495.79±145.39 | 16.95±8.51 | 147.22±23.96 | 208.71±72.81 | 33.06±4.04 |
| MGL | 460.04±84.60 | 17.31±9.22 | 197.59±42.31 | 187.71±51.35 | 32.98±3.51 |
| YK | 640.55±106.58 | 24.24±12.80 | 223.37±37.97 | 224.79±101.28 | 39.15±6.14 |
| AN:速效氮Available nitrogen;AP:速效磷Available phosphorus;AK:速效钾Available potassium;DOC:可溶性有机碳Dissolved organic carbon;SWC:土壤含水率Soil water content | |||||
土壤动物采集:
在3种森林生态系统中各设置3块20 m×30 m样地, 以构成3个重复。2012年5—10月, 每月对3种森林生态系统土壤动物进行取样调查。
大型土壤动物采用3点混合取样法, 取样面积为50 cm×50 cm, 取样深度为0—20 cm, 手拣法就地分拣, 固定于75%的酒精中, 带回实验室。
中小型土壤动物及土壤理化性质采用5点混合取样法, 用直径为5 cm的土钻进行采集, 取样深度为0—20 cm, 将土样装入封口袋带回实验室, 分两部分, 一部分采用Tullgren法对中小型土壤动物进行分离, 另一部分自然风干后进行土壤理化性质的测定。
土壤总呼吸(RS)测定:
2004年4月中旬已在3个森林生态系统的9块样地内均随机布设8个PVC土壤环(内径10.2cm、高8cm)。于2012年5—10月期间, 采用Li-6400便携式CO2/H2O分析系统(Li-Cor Inc., Lincoln, NE, USA)每两周测定一次土壤总呼吸速率。仅选取生长季进行测定是由于Li-6400便携式CO2/H2O分析系统在低温下难以运行。
土壤异养呼吸(RH)测定:
采用挖壕法[20]测定土壤异养呼吸。在3个森林生态系统的9块样地距边界2—3 m处随机划分4个50 cm×50 cm的样方, 挖至植物根系分布层以下55—75 cm处, 后用双层厚塑料隔离样方周围的根系, 同时除去样方内的所有活的植物体, 安置PVC土壤环(土壤环的安置方法同土壤总呼吸测定), 且与土壤总呼吸同步测定。该处理已于2004年4月初进行[16]。
1.3.2 室内分析土壤动物分类鉴定:
将分离的大型土壤动物带回实验室后立即洗净, 用滤纸吸干, 分别在分析天平上称量并记录其鲜重。然后在显微镜下依据《中国土壤动物检索图鉴》[21]对大、中小型土壤动物进行分类鉴定并计数, 通常鉴定到科或属, 对于少数土壤动物鉴定到目或纲。根据取样面积将生物量(g)和个体数(个)换算成生物量(g/m2)和个体密度(个/m2), 其中个体数为大中小型土壤动物个体数之和, 生物量仅为大型土壤动物鲜重。
土壤理化性质测定:
采用扩散法测定土壤速效氮(AN);采用电感耦合等离子体发射光谱(ICPS-7500, 日本岛津)在醋酸氨浸提液中测定土壤速效钾(AK);采用连续流动化学分析仪(SKALAR SAN++, 荷兰SKALAR)在碳酸氢钠浸提液中测定土壤速效磷(AP);采用总有机碳分析仪(TOC-VCPH, 日本岛津)分析测定土壤可溶性有机碳(DOC);取PVC环附近土壤, 采用烘干法测定土壤含水率(SWC);采用数字式瞬时温度计测定10cm的土壤温度(Ts)。
1.4 数据分析土壤呼吸数据处理方法:
由于受测定时间的限制, 无法对3种森林生态系统的RS及RH进行同步测定, 且测定的瞬时温度也存在差异, 因此采用指数模型方程计算各样地土壤动物取样时温度所对应的RS及RH。
采用Van′t Hoff指数模型拟合土壤呼吸速率与土壤温度的关系[22]。公式如下:
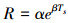
|
式中, R代表土壤总呼吸速率(RS)或土壤异养呼吸(RH)(mol CO2 m-2 s-1), Ts为10cm的土壤温度(℃);α是土壤温度为0 ℃时的土壤呼吸速率(mol CO2 m-2 s-1);β为温度反应系数。在上述模型拟合时, 需将RS及RH进行自然对数转换, 以满足模型的线性和方差齐性需求。
土壤总呼吸月增长率=(当月土壤总呼吸-上月土壤总呼吸)/上月土壤总呼吸× 100%, 土壤异养呼吸、土壤动物生物量及个体密度月增长率同上。
数据统计分析采用SPSS 22, 作图采用SigmaPlot绘图软件。采用多因素方差分析(ANOVA, LSD)检验月份和林型对3种森林生态系统土壤总呼吸、土壤异养呼吸、土壤动物生物量及土壤动物个体密度等的显著性影响。采用双变量相关分析和线性模型y=y0+ax对土壤动物与土壤呼吸的关系进行拟合分析。
2 结果与分析 2.1 3种森林生态系统土壤总呼吸(RS)及土壤异养呼吸(RH)的动态变化根据方差分析结果可得, 3种森林生态系统的RS大致表现先增强后减弱的显著的时间动态变化(P<0.05)(表 3), 其中红松人工林和硬阔叶林为8月份最高, 蒙古栎林为6月份最高, 3种森林生态系统RS最低值均出现在10月份(图 1)。不同森林生态系统的RS无显著差异(P>0.05)(表 3)。
| 因子 Factor |
土壤总呼吸RS Soil respiration |
土壤异养呼吸RH Soil heterotrophic respiration |
生物量B Biomass |
个体密度) Individual density |
|||||||||||
| df | F | P | df | F | P | df | F | P | df | F | P | ||||
| 林型Forest type | 2 | 2.498 | 0.097 | 2 | 8.416 | 0.001 | 2 | 0.227 | 0.798 | 2 | 4.214 | 0.023 | |||
| 月份Month | 5 | 8.29 | <0.001 | 5 | 8.379 | <0.001 | 5 | 6.728 | <0.05 | 5 | 9.208 | <0.05 | |||
| 林型×月份 Forest type×Month |
10 | 1.067 | 0.413 | 10 | 3.229 | 0.005 | 10 | 0.534 | 0.855 | 10 | 2.02 | 0.06 | |||
| P<0.05为显著水平, P<0.01为极显著水平 | |||||||||||||||

|
| 图 1 3种森林生态系统土壤总呼吸速率及月增长率的时间动态变化 Fig. 1 Temporal dynamic of soil total respiration rate and monthly growth rate in three forest ecosystems |
RH也呈现显著时间动态变化(P<0.05), 且不同森林生态系统的RH具有显著差异(P<0.05)(表 3)。由图 2可知3种森林生态系统的RH最高值出现的时间不同, 红松人工林9月RH最高、蒙古栎林8月RH最高, 硬阔叶林7月RH最高, 3种森林生态系统RH的最低值均出现在10月份。红松人工林和硬阔叶林的RH变化趋势相似, 均呈“增强-减弱-增强-减弱”的“M型”变化趋势, 而蒙古栎林RH呈先增强后减弱的“倒V型”变化趋势(图 2)。

|
| 图 2 3种森林生态系统土壤异养呼吸速率及月增长率的时间动态变化 Fig. 2 Temporal dynamic of soil heterotrophic respiration rate and monthly growth rate in three forest ecosystems |
RS、RH随时间的变化趋势虽然大体一致, 但变化幅度不同。5—6月及7—8月RH及RS均表现为增长, 但5—6月期间RH增幅大于RS增幅, 而7—8月RS增幅大于RH增幅;6—7月期间RH及RS均表现为下降, RS下降较RH明显;8—9月RH继续增长而RS则下降(图 1, 图 2)。
2.2 3种森林生态系统土壤动物生物量(B)、个体密度(N)的动态变化对帽儿山3种森林生态系统0—20 cm土层的土壤动物进行取样, 共获得土壤动物69类, 74145只, 其中大型土壤动物59类, 3604只, 其优势类群为正蚓科(Lumbricidae)、线蚓科(Enchytraeidae)和石蜈蚣目(Lithobiomorpha);中小型土壤动物10类, 70541只, 其优势类群为甲螨亚目(Oribatida), 等节跳科(Isotomidae), 中气门亚目(Mesostigmta)[6]。
通过双因素方差分析结果表明土壤动物生物量呈现显著的时间动态变化(P<0.05)(表 3)。由图 3可得不同月份红松人工林、蒙古栎林和硬阔叶林3种森林生态系统土壤动物生物量变化趋势相似, 均表现5—8月份变化幅度小、差异不显著, 9月份出现最高峰, 10月份下降, 而土壤动物生物量对立地条件的响应不显著(P>0.05)(表 3)。

|
| 图 3 3种森林生态系统土壤动物生物量及月增长率的时间动态变化 Fig. 3 Temporal dynamic of soil animal biomass and monthly growth rate in three forest ecosystems |
土壤动物个体密度也具有显著时间动态变化(P<0.05)(表 3), 但与生物量的变化规律不同, 其中, 5—7月份3种森林生态系统土壤动物个体密度均呈现先下降后上升趋势, 但红松人工林各月份间没有显著差异;7—8月份红松人工林和硬阔叶林呈下降趋势, 蒙古栎林呈继续上升趋势;9月份各森林生态系统土壤动物个体密度均达到峰值, 且MGL > YK > HS;10月份又均呈现变化幅度不同的下降趋势。研究表明不同森林生态系统土壤动物个体密度的变化趋势具有显著差异(P<0.05), 红松人工林土壤动物个体密度变化趋势与硬阔叶林大致相似, 与蒙古栎林差异较大, 且土壤动物个体密度整体上表现为YK > HS > MGL(图 4)。

|
| 图 4 3种森林生态系统土壤动物个体密度及月增长率的时间动态变化 Fig. 4 Temporal dynamic of soil animal individual density and monthly growth rate in three forest ecosystems |
土壤动物生物量及个体密度变化幅度也不同。土壤动物生物量及个体密度在8—9月均为显著增长且生物量增长幅度远大于个体密度增长幅度, 9—10月份均为负增长且生物量下降幅度大于个体密度下降幅度;5—6月和7—8月土壤动物生物量均为增长趋势, 而个体密度为负增长;6—7月土壤动物生物量为负增长, 个体密度为增长(图 3、图 4)。
2.3 土壤动物与土壤呼吸的相互关系用线性模型y=y0+ax对土壤总呼吸(RS)、异养呼吸(RH)及土壤动物生物量(B)、个体密度(N)进行拟合, 拟合前将数据进行自然对数转换, 以满足模型的线性和方差齐性需求。由分析可得土壤动物生物量与土壤总呼吸、异养呼吸呈负相关, 土壤动物个体密度与土壤总呼吸、异养呼吸呈负相关, 但两两之间均无显著线性回归关系。不同森林生态系统土壤呼吸与土壤动物相关性不同, 其中红松人工林中, 除土壤总呼吸速率与土壤动物个体密度呈负相关外, 土壤总呼吸速率与土壤动物生物量、土壤异养呼吸速率与土壤动物生物量及个体密度均呈正相关;而在蒙古栎林和硬阔叶林中, 土壤总呼吸速率、土壤异养呼吸速率与土壤动物生物量及个体密度两两之间均呈负相关(图 5)。

|
| 图 5 土壤动物与土壤呼吸相关性分析 Fig. 5 Analysis of correlation between soil animals and soil respiration |
通过双变量相关分析发现土壤呼吸与土壤动物相关性也随时间变化具有明显差异。在生长季初期和末期, 土壤呼吸与土壤动物相关性大致呈负相关:5月除土壤异养呼吸与土壤动物个体密度呈正相关外, 土壤总呼吸与土壤动物生物量和个体密度、土壤异养呼吸与土壤动物生物量均呈负相关, 6月、10月表现为土壤总呼吸、异养呼吸与土壤动物生物量、个体密度均为负相关;而在生长季中期7月、8月时, 土壤总呼吸、异养呼吸与土壤动物个体密度呈正相关, 与土壤动物生物量呈负相关, 9月时土壤总呼吸、异养呼吸与土壤动物生物量、个体密度均为正相关(图 6)。

|
| 图 6 土壤动物与土壤呼吸相关性的时间动态变化 Fig. 6 Temporal dynamic changes of the correlation between soil animals and soil respiration |
一般认为土壤温度和水分均为引起土壤呼吸时间动态变化的主要影响因子[23], 我们的研究发现土壤总呼吸及土壤异养呼吸均呈现在生长季的初期和末期较低而中期较高的显著性时间动态变化趋势, 这与大多数的研究结果相一致。李思思等[24]在对青海高寒区5种森林生态系统土壤总呼吸季节变化研究中表明土壤所处地区的气候条件(特别是温度和水分)会直接或间接地影响土壤总呼吸速率。帽儿山地区属温带大陆性季风气候, 生长季中期时气温较高, 降水较多, 土壤温度及水分状况适宜, 有利于土壤微生物、土壤动物及植物根系的呼吸, 同时生长季中期植物根系分泌大量易分解的碳水化合物到土壤中[25], 加快土壤中的化学氧化反应, 共同促进土壤总呼吸及异养呼吸增强;而是生长季前后期土壤温度、水分相对较低, 可利用碳源较少, 改变了土壤生物群落结构, 降低生物及根系生物量, 抑制土壤氧化酶活性, 从而削弱了土壤总呼吸及异养呼吸速率。
从增长率角度研究发现, 土壤总呼吸及土壤异养呼吸速率在5—6月份和7—8月份均为增长趋势而8—9月份土壤异养呼吸速率增长而土壤总呼吸速率下降。由于5—6月份是春季, 地上部分枝叶开始萌芽生长, 植物要以最大的地下部分的根系量来满足地上部分的养分供应, 因此细根生物量最大, 此时植物根系对土壤呼吸起主导作用[26], 7—8月份是植物生长最旺盛的季节, 此时植物根系对土壤呼吸作用也较为显著, 到了8—9月份秋季即生长季的后期, 地上部分生长减慢, 地下部分细根现存量降低[26], 土壤总呼吸呈负增长。
不同森林生态系统由于受到多种土壤环境因子的影响, 其土壤异养呼吸也具有显著差异。本研究表明立地状况不同的3种森林生态系统土壤异养呼吸差异显著, 表现为红松人工林最低, 硬阔叶林最高, 即阔叶林>针叶林, 国庆喜等[27]在修正的IBIS模拟的土壤呼吸的动态特征研究中也得出了相同的结论。Raich等指出由于针叶林土壤碳循环速度相对阔叶林土壤要低, 所以阔叶林的异养呼吸速率要高于针叶林[28]。这可能是由于不同森林生态系统凋落物质量、土壤养分水分等环境因素差异造成的[29], 硬阔叶林中土壤养分、水分大, 凋落物较多, 土壤微生物呼吸的底物增加, 导致硬阔叶林中土壤异养呼吸速率较高。
3.2 3种森林生态系统土壤动物生物量(B)和个体密度(N)的动态变化前人的研究表明土壤动物生物量及个体密度受土壤、气候[30-31]、植被类型[32]等多种因子影响, 一年中土壤动物会随着温度及降水的变化发生明显的季节性动态变化。张荣祖等在对长白山北坡森林生态系统土壤动物初步调查中发现在寒温带和中温带地区土壤动物的数量通常在7—9月最多, 这与其温度和降水量的变化基本上是一致的[33]。帽儿山地区属寒温带, 研究发现帽儿山3种森林生态系统土壤动物个体密度及生物量均具有季节性动态变化, 9月份土壤动物个体密度及生物量增幅均最大, 与李娜等[6]对大型土壤动物个体密度的研究结果有所不同, 可能是因为中小型土壤动物个体密度在秋季达到最高[34]而引起的差异, 中小型土壤动物迁移能力较弱, 秋季降水较少, 土壤中含水量适宜中小型土壤动物生长繁殖。
不同森林生态系统土壤动物个体密度差异与土壤性质及植被条件有关[33], 其中土壤环境因子是引起土壤动物群落结构发生变化的重要影响因素。本研究结果表明不同森林生态系统土壤动物个体密度为YK > HS > MGL, 但HS与YK相差不大。有研究表明土壤中营养物质的不同会导致土壤动物群落结构有差异[34], 速效氮、速效磷、速效钾、可溶性有机碳等作为土壤动物重要的营养源, 可为土壤动物提供其生存所必需的能量。表 2可得硬阔叶林中速效氮、速效磷、速效钾、可溶性有机碳含量均为3种森林生态系统中最高, 为土壤动物提供了大量营养物质, 因此导致土壤动物个体密度较大。
3.3 土壤动物与土壤呼吸的相关性分析土壤动物是影响土壤呼吸的重要因子, 其组成和群落结构的变化应对土壤呼吸, 尤其是土壤异养呼吸产生显著影响。而本研究结果表明土壤总呼吸、土壤异养呼吸与土壤动物生物量及个体密度均呈负相关, 即随着土壤动物生物量及个体密度的增加, 土壤总呼吸、土壤异养呼吸速率均降低, 但两两回归关系并不显著, 与假设有异。可能是由于土壤动物生物量及个体密度的增加导致微生物及根系生物量减少, 最终导致土壤总呼吸及异养呼吸降低。
大部分学者研究表明土壤动物可促进土壤呼吸的增加, 例如热带雨林中蚂蚁和白蚁巢穴CO2排放量高于周围散土区[35]。但也有研究表明土壤动物也可通过影响微生物及植物根系对土壤呼吸产生抑制作用。一方面土壤动物会对土壤微生物的生命活动产生一定的抑制作用, 从而进一步抑制土壤呼吸。部分土壤动物对于土壤微生物具有选择性捕食的作用而导致微生物生物量降低, 最终抑制土壤微生物呼吸作用[13], 例如, 蚯蚓、线虫、跳虫等。Eisenhauer等[36]在蚯蚓入侵北美北部阔叶林的研究中发现蚯蚓可使土壤微生物量显著降低。土壤中食微线虫可通过捕食微生物来抑制土壤呼吸及异养呼吸。Rohan等[37]研究发现不同种弹尾目跳虫对微生物呼吸均有显著抑制作用。另一方面土壤动物可通过降低根系呼吸来抑制土壤呼吸。部分土壤动物以植物根系为食, 直接影响根系生物量及周转速率, 加速根的死亡, 从而影响根系呼吸。植食性土壤动物主要包括昆虫类土壤动物和植物寄生线虫。Ueckert[38]研究表明在农田生态系统中植食性瓢虫可使地下根系生物量降低43%, 而Stevens等[39]研究发现在施用杀虫剂处理后可明显增加细根生物量。
前人研究表明土壤动物呼吸仅占土壤呼吸的5%—10%, 且具有空间异质性[4], 而综合土壤动物对土壤微生物及植物根系的影响, 可表明土壤动物主要是通过直接影响土壤微生物及植物根系对土壤呼吸产生较大的间接的影响。而目前对于土壤动物、土壤微生物及植物根系三者之间的关系了解较少, 后续还应进一步探讨土壤动物与土壤微生物及植物根系的相互关系, 建立三者之间的关系模型, 进一步明确土壤动物与土壤呼吸响应机制。同时, 对不同生态系统土壤呼吸数据进行比较分析时, 也要考虑实验方法带来的影响。首先, 目前大多数土壤动物和土壤呼吸相关性研究以较容易控制的室内培养实验为主, 但室内培养实验的条件与野外控制实验的自然环境相差较大[14], 尤其在土壤温度及水分条件方面, 同时室内培养实验也忽略了植物在土壤动物与土壤呼吸相互关系研究中的重要影响, 因此在未来的研究中需要考虑土壤中多种因子影响以提高室内培养实验的准确性, 同时还要将室内培养实验与野外控制实验相结合[40], 以便于更准确的揭示土壤动物与土壤呼吸相互关系。其次, 本研究所使用的壕沟法对于壕沟内外的土壤动物群落结构可能会产生一定影响, 进而影响土壤动物与土壤呼吸的相互关系[41], 目前还未有较好的解决方法, 未来还应继续在技术方法上进行创新, 有助于更准确建立土壤动物与碳循环之间的联系。
温度和水分是控制土壤CO2排放的主要因子。有研究表明, 在增温条件下土壤动物对CO2排放具有较强的促进作用[42]。而本研究在土壤动物与土壤呼吸相关性的时间动态变化分析中发现生长季初期和末期, 土壤动物生物量和个体密度与土壤呼吸呈负相关, 表明此时土壤动物对土壤呼吸具有抑制作用, 这可能是由于生长季初期和末期土壤温度和水分均较低所导致。
4 结论由于不同月份土壤温度及水分差异大导致土壤总呼吸、异养呼吸及土壤动物生物量、个体密度及其相关性均有显著性时间动态变化, 且不同森林生态系统土壤异养呼吸及土壤动物个体密度有显著差异, 均在硬阔叶林中最高。相关性分析表明, 土壤动物可通过抑制微生物生命活动和降低根系呼吸从而对土壤总呼吸及异养呼吸产生负反馈作用, 三者是不可分割的整体, 与温度、水分等环境因子共同调控着土壤呼吸。因此继续挖掘土壤动物、微生物及植物根系三者之间的响应机制, 探讨温度和水分的变化对土壤动物和土壤呼吸相关性的影响, 对进一步探讨土壤动物在土壤碳循环中的功能和作用并揭示土壤碳平衡的变化具有重要的意义。
| [1] |
IPCC. Climate Change 2014:I:Impacts, Adaptation, and Vulnerability. Part A:Global and Sectoral Aspects. Contribution of Working Group Ⅱ to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. United Kingdom: Cambridge University Press, 2014.
|
| [2] |
Kagawa S, Suh S, Hubacek K, Wiedmann T, Nansai K, Minx J. CO2 emission clusters within global supply chain networks:implications for climate change mitigation. Global Environmental Change, 2015, 35: 486-496. DOI:10.1016/j.gloenvcha.2015.04.003 |
| [3] |
王泽西, 陈倩妹, 黄尤优, 邓慧妮, 谌贤, 唐实玉, 张健, 刘洋. 川西亚高山森林土壤呼吸和微生物生物量碳氮对施氮的响应. 生态学报, 2019, 39(19): 7197-7207. |
| [4] |
王兵, 姜艳, 郭浩, 赵广东, 白秀兰. 土壤呼吸及其三个生物学过程研究. 土壤通报, 2011, 42(2): 483-490. |
| [5] |
战丽莉.农田黑土中小型土壤动物多样性特征及其影响因素[D].长春: 中国科学院研究生院(东北地理与农业生态研究所), 2013.
|
| [6] |
李娜, 张雪萍, 张利敏. 三种温带森林大型土壤动物群落结构的时空动态. 生态学报, 2013, 33(19): 6236-6245. |
| [7] |
Sørensen L I, Holmstrup M, Maraldo K, Christensen S, Christensen B. Soil fauna communities and microbial respiration in high Arctic tundra soils at Zackenberg, Northeast Greenland. Polar Biology, 2006, 29(3): 189-195. DOI:10.1007/s00300-005-0038-9 |
| [8] |
Fu S L, Coleman D C, Hendrix P F, Crossley Jr D A. Responses of trophic groups of soil nematodes to residue application under conventional tillage and no-till regimes. Soil Biology and Biochemistry, 2000, 32(11/12): 1731-1741. |
| [9] |
姜炳棋, 马旖旎, 单军, 郭红岩, 季荣. 威廉腔环蚓(Metaphire guillelmi)对14C-土壤有机质转化的研究. 生态环境学报, 2011, 20(6/7): 1011-1017. |
| [10] |
Luo T X, Li H X, Wang T, Hu F. Influence of nematodes and earthworms on the emissions of soil trace gases (CO2, N2O). Acta Ecological Sonica, 2008, 28(3): 993-999. DOI:10.1016/S1872-2032(08)60033-5 |
| [11] |
Jílková V, Picek T, Frouz J. Seasonal changes in methane and carbon dioxide flux in wood ant (Formica aquilonia) nests and the surrounding forest soil. Pedobiologia, 2015, 58(1): 7-12. DOI:10.1016/j.pedobi.2014.12.001 |
| [12] |
王可洪, 武海涛, 吴东辉. 土壤动物对土壤呼吸的影响研究进展. 土壤通报, 2013, 44(6): 1526-1530. |
| [13] |
徐璇, 王维枫, 阮宏华. 土壤动物对森林凋落物分解的影响:机制和模拟. 生态学杂志, 2019, 38(9): 2858-2865. |
| [14] |
聂立凯, 于政达, 孔范龙, 李悦, 郗敏. 土壤动物对土壤碳循环的影响研究进展. 生态学杂志, 2019, 38(3): 882-890. |
| [15] |
刘领, 王艳芳, 悦飞雪, 李冬, 赵威. 基于森林清查资料的河南省森林植被碳储量动态变化. 生态学报, 2019, 39(3): 864-873. |
| [16] |
牛莉平, 许嘉巍, 靳英华, 尹海霞, 王嫒林, 刘羽霞, 张英洁. 温带森林不同演替阶段下的土壤CO2排放通量昼间变化. 生态学报, 2017, 37(8): 2673-2683. |
| [17] |
张慧东, 尤文忠, 魏文俊, 周梅. 暖温带-中温带过渡区4种典型森林土壤呼吸的温度敏感性. 生态环境学报, 2015, 24(11): 1757-1764. |
| [18] |
杨金艳, 王传宽. 东北东部森林生态系统土壤碳贮量和碳通量. 生态学报, 2005, 25(11): 2875-2882. DOI:10.3321/j.issn:1000-0933.2005.11.012 |
| [19] |
杨金艳, 王传宽. 土壤水热条件对东北森林土壤表面CO2通量的影响. 植物生态学报, 2006, 30(2): 286-294. DOI:10.3321/j.issn:1005-264X.2006.02.011 |
| [20] |
Bond-Lamberty B, Wang C K, Gower S T. A global relationship between the heterotrophic and autotrophic components of soil respiration?. Global Change Biology, 2004, 10(10): 1756-1766. DOI:10.1111/j.1365-2486.2004.00816.x |
| [21] |
尹文英. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
|
| [22] |
Luo Y Q, Wan S Q, Hui D F, Wallace L L. Acclimatization of soil respiration to warming in a tall grass prairie. Nature, 2001, 413(6856): 622-625. DOI:10.1038/35098065 |
| [23] |
刘顺, 杨洪国, 罗达, 史作民, 刘千里, 张利. 川西亚高山不同森林类型土壤呼吸和总硝化速率的季节动态. 生态学报, 2019, 39(2): 550-560. |
| [24] |
李思思, 贺康宁, 田赟, 左巍, 王玮璐, 唐达, 张潭, 李倩. 青海高寒区5种典型林分土壤呼吸季节变化及其影响因素. 北京林业大学学报, 2016, 38(10): 95-103. |
| [25] |
杨智杰, 郑裕雄, 陈仕东, 刘小飞, 熊德成, 林伟盛, 胥超, 杨玉盛, 史思红. 应用小波多尺度分析亚热带森林土壤异养呼吸特征. 生态学报, 2018, 38(14): 5078-5086. |
| [26] |
朱胜英, 周彪, 毛子军, 王秀伟, 孙元发. 帽儿山林区6种林分细根生物量的时空动态. 林业科学, 2006, 42(6): 13-19. |
| [27] |
国庆喜, 张海燕, 王兴昌, 王传宽. 东北典型森林土壤呼吸的模拟——IBIS模型的局域化应用. 生态学报, 2010, 30(9): 2295-2303. |
| [28] |
Raich J W, Tufekciogul A. Vegetation and soil respiration:correlations and controls. Biogeochemistry, 2000, 48(1): 71-90. DOI:10.1023/A:1006112000616 |
| [29] |
陈灿, 江灿, 范海兰, 林勇明, 吴承祯. 凋落物去除/保留对杉木人工林林窗和林内土壤呼吸的影响. 生态学报, 2017, 37(1): 102-109. |
| [30] |
李红月, 殷秀琴, 马辰, 郭玉梅. 长白山地丘陵区不同土地利用方式土壤动物群落生态分布特征. 土壤学报, 2017, 54(4): 1018-1028. |
| [31] |
杨效东, 刘宏茂, 郑征, 沙丽清. 砂仁种植对季节雨林土壤节肢动物群落结构影响的初步研究. 生态学杂志, 2003, 22(4): 10-15. DOI:10.3321/j.issn:1000-4890.2003.04.003 |
| [32] |
寇新昌, 殷秀琴. 长白山地不同次生林土壤动物群落多样性特征及其分布格局. 山地学报, 2017, 35(4): 429-436. |
| [33] |
张荣祖, 杨明宪, 陈鹏, 张庭伟. 长白山北坡森林生态系统土壤动物初步调查. 森林生态系统研究, 1980(1): 133-152. |
| [34] |
韩慧莹, 殷秀琴, 寇新昌. 长白山地低山区土壤动物群落特征及其对环境因子变化的响应. 生态学报, 2017, 37(7): 2197-2205. |
| [35] |
Ohashi M, Maekawa Y, Hashimoto Y, Takematsu Y, Hasin S, Yamane S. CO2 emission from subterranean nests of ants and termites in a tropical rain forest in Sarawak, Malaysia. Applied Soil Ecology, 2017, 117-118: 147-155. DOI:10.1016/j.apsoil.2017.04.016 |
| [36] |
Eisenhauer N, Schlaghamerský J, Reich P B, Frelich L E. The wave towards a new steady state:Effects of earthworm invasion on soil microbial functions. Biological Invasions, 2011, 13(10): 2191-2196. DOI:10.1007/s10530-011-0053-4 |
| [37] |
Cragg R G, Bardgett R D. How changes in soil faunal diversity and composition within a trophic group influence decomposition processes. Soil Biology and Biochemistry, 2001, 33(15): 2073-2081. DOI:10.1016/S0038-0717(01)00138-9 |
| [38] |
Ueckert D N. Impact of a white grub (Phyllophaga crinita) on a shortgrass community and evaluation of selected rehabilitation practices. Journal of Range Management, 1979, 32(6): 445-448. DOI:10.2307/3898556 |
| [39] |
Stevens G N, Jones R H. Patterns in soil fertility and root herbivory interact to influence fine-root dynamics. Ecology, 2006, 87(3): 616-624. DOI:10.1890/05-0809 |
| [40] |
卢明珠, 吕宪国, 管强, 武海涛. 蚯蚓对土壤温室气体排放的影响及机制研究进展. 土壤学报, 2015, 52(6): 1209-1225. |
| [41] |
范志平, 王红, 邓东周, 孙学凯, 高俊刚, 曾德慧. 土壤异养呼吸的测定及其温度敏感性影响因子. 生态学杂志, 2008, 27(7): 1221-1226. |
| [42] |
卢明珠.三江平原湿地蚯蚓对土壤温室气体排放的影响[D].长春: 中国科学院研究生院(东北地理与农业生态研究所), 2016.
|