 2021, Vol. 41
2021, Vol. 41文章信息
- 李泽东, 陈志成, 曹振, 车路平, 刘舒文, 张永涛
- LI Zedong, CHEN Zhicheng, CAO Zhen, CHE Luping, LIU Shuwen, ZHANG Yongtao
- 华北低山丘陵区常用树种木质部解剖特征及水其力学抗旱性
- Xylem anatomical and hydraulic drought resistance characteristics of common tree species in hilly areas of North China
- 生态学报. 2021, 41(1): 69-78
- Acta Ecologica Sinica. 2021, 41(1): 69-78
- http://dx.doi.org/10.5846/stxb201910132134
-
文章历史
- 收稿日期: 2019-10-13
- 网络出版日期: 2020-11-19





2. 山东泰山森林生态系统国家定位研究站, 泰安 271018;
3. 中国林业科学研究院林业新技术研究所, 北京 100091
2. Mountain Tai Forest Ecosystem Research Station of State Forestry Administration, Taian 271018, China;
3. Institute of New Forestry Technology, Chinese Academy of Forestry, Beijing 100091, China
全球气候变化导致的越来越频繁的干旱事件在世界范围内引起了严重的森林衰败死亡, 干旱导致树木死亡的生理学机制和树木应对干旱胁迫的适应机制成为了目前的研究热点[1]。树木对干旱胁迫的适应能力决定了其在生态系统中的分布, 其生理性状变化是适应外部环境的客观表达, 也与生存策略密切相关[2]。以往学者的研究大多集中在植物生理生化指标对干旱胁迫的响应, 包括抗旱性评价[3-6], 但共生的不同树种间水力输导系统应对干旱的适应策略方面的研究较少。
目前认为干旱导致树木死亡存在两种生理学机制, 水力失衡和碳饥饿[1, 7]。水力失衡是严重干旱胁迫下树木的水力运输功能丧失而干化死亡。依据内聚力-张力理论(Cohesion-Tension Theory), 植物主要是通过蒸腾作用的拉力将植物根部吸收的水分经木质部导管(或管胞)向上运输, 水分子依靠内聚力保证导管中的水柱不至于断裂[8]。干旱胁迫时木质部中的张力变大, 气体会通过导管(或管胞)上的纹孔进入导管产生栓塞, 干旱超过一定阈值时, 会出现气穴化, 阻断水分运输的连续性, 从而影响植物一系列生理活动甚至导致植株死亡[9]。树木木质部输水结构与其抗栓塞能力紧密相关[10-12], 通过对植物木质部输水结构的研究可以更深入全面的理解植物抗旱性。木质部中的导管或管胞提供输水功能, 纤维组织提供支撑功能, 而薄壁组织作为木质部中的活细胞提供储存功能。而且近来的微CT成像技术表明薄壁组织还对导管的栓塞修复起到重要作用[13]。根据Hagen-Poiseuille定律, 导管的输水效率与导管直径是四次方关系, 与导管密度是累加关系[14]。一般来说, 抗旱植物具有小而密的导管, 纹孔膜较厚, 导水率较低但具有更强的抗栓塞能力;非抗旱树种通常具有较大导管直径和较高的导水率, 但是抵抗栓塞能力偏低, 因此很多研究发现不同树种间木质部导管存在水分运输的高效性和安全性上的权衡关系[15-17], 但是Gleason等在全球尺度上进行meta分析, 发现只存在弱的水力安全-高效权衡[18]。碳饥饿是光合作用等非结构性碳(NSC)供应量小于呼吸作用等NSC需求量, 低于一定阈值时新陈代谢受限, 或者与水力失衡交互导致NSC无法分配或利用。NSC还为栓塞修复提供必不可少的能量和物质, 而木质部中的NSC主要储存在薄壁组织中。薄壁组织主要分为射线薄壁组织和轴向薄壁组织, 轴向薄壁组织根据是否与导管相连, 又可分为旁管薄壁组织和离管薄壁组织, 而与导管相连的旁管薄壁组织对栓塞修复起重要作用[19]。
华北低山丘陵区土壤贫瘠, 干旱缺水, 水土流失严重, 具有众多的未造林或立地条件差的石质丘陵山地, 严重制约着这些未开发山地的生产力[20]。而植被恢复是华北低山丘陵地区生态环境恢复的重要措施[21], 所以采用植物措施对干旱瘠薄山地进行绿化, 对改善当地生态环境、固碳增汇、有着重要的作用。基于植物木质部解剖及非结构性碳的测定, 探讨华北低山丘陵区常用树种水力学抗旱性特征尚未见报道, 研究不同树种水力结构的差异及其在结构上表现出的适应性策略, 不仅有利于抗旱植物的筛选, 也将更好地揭示干旱条件下植物生存、演化、及区域群落结构动态过程, 为该区植物保护和植被恢复提供依据。在本研究中, 根据以往学者的研究[22-25]以及实际造林经验选出了抗旱树种(低山丘陵区造林树种)和非抗旱树种(城市绿化树种)两类, 其中抗旱树种包括臭椿(Ailanthus altissima)、黄栌(Cotinus coggygria)、麻栎(Quercus acutissima)、榆树(Ulmus pumila)和胡桃(Juglans regia), 非抗旱树种包括白玉兰(Magnolia denudata)、鹅掌楸(Liriodendron chinensis)、紫荆(Cercis chinensis)和二球悬铃木(Platanus acerifolia)。测定了以上10个树种木质部横截面解剖的大量性状指标及NSC浓度, 主要研究以下内容:1)抗旱树种的木质部导管的解剖特征相较于非抗旱树种有怎样的差异;2)这种差异表现出两类树种具有怎样不同的应对干旱的策略。
1 材料与方法 1.1 试验地概况研究地点位于山东农业大学植物园内, 位于泰安市南部(E 117°11′, N 36°16′), 属于温带季风大陆性气候, 四季分明。植被属暖温带落叶阔叶林区域, 植物区系属于华北植物区系, 包括森林、灌丛、灌草丛、草甸、等生态系统;多年平均气温13℃, 极端最高气温40℃, 极端最低气温-20.7℃, 7月气温最高, 平均26.4℃, 1月最低, 平均-2.6℃;年平均降水量697mm, 最大年降水量1083.00mm, 最小年降水量434.0mm, 降水分布不均, 易发生春旱;年日照时数2355h;无霜期198d;土壤以棕壤、褐土为主, 肥力中等。
1.2 试验材料处理方法试验于2018年6月进行, 在山东农业大学植物园内采集不同树种的枝条样品。每个树种选取3株胸径相近的树作为3个重复, 树种信息见表 1。用高枝剪剪下树冠中上部、长势良好且向阳的1—2年生枝条, 枝条直径0.8—1.2cm, 取下后立即带回实验室处理。将枝条上所有叶片取下用于总叶面积的测定, 余下枝条的一部分用于木质部切片的制作, 另一部分剥皮去髓, 放入烘箱105℃杀青, 80℃烘干48h, 之后用粉碎机粉碎并过100目筛用于木质部NSC浓度的测定。
| 树种 Species |
代码 Code |
科 Family |
散孔材/环孔材 Diffuse porous wood/ Ring porous wood |
抗旱性 Drought resistance |
| 臭椿Ailanthus altissima | Aa | 苦木科Simaroubaceae | 环孔材 | 抗旱树种 |
| 黄栌Cotinus coggygria | Cc | 漆树科Anacardiaceae | 环孔材 | 抗旱树种 |
| 麻栎Quercus acutissima | Qa | 壳斗科Fagaceae | 环孔材 | 抗旱树种 |
| 榆树Ulmus pumila | Up | 榆科Ulmaceae | 散孔材 | 抗旱树种 |
| 胡桃Juglans regia | Jr | 胡桃科Juglandaceae | 环孔材 | 抗旱树种 |
| 白玉兰Magnolia denudata | Md | 木兰科Magnoliaceae | 散孔材 | 非抗旱树种 |
| 鹅掌楸Liriodendron chinensis | Lc | 木兰科Magnoliaceae | 散孔材 | 非抗旱树种 |
| 紫荆Cercis chinensis | Ch | 豆科Fabaceae | 散孔材 | 非抗旱树种 |
| 灯台树Bothrocaryum controversum | Bc | 山茱萸科Cornaceae | 散孔材 | 非抗旱树种 |
| 二球悬铃木Platanus acerifolia | Pt | 悬铃木科Platanaceae | 散孔材 | 非抗旱树种 |
| R:环孔材;D:散孔材;S:抗旱树种;W:非抗旱树种 | ||||
用LI-3100C叶面积仪测定叶面积;枝条边材面积用扫描仪测定, 边材面积与总叶面积的比值为Huber值。NSC是可溶性糖和淀粉的总和[26], 采用改良的蒽酮-硫酸法测定[27]。
木质部切片制作:将3cm长枝条固定在滑动切片机上进行切片, 切片厚度在18—22μm, 完成后在番红-阿尔新蓝溶液中染色60—90s, 在不同浓度的乙醇溶液中脱水, 在载玻片上固定之后在光学显微镜下观察, 在10×物镜下观察当年和上一年的木质部解剖, 选区3—5个视野拍照。导管直径、导管密度、薄壁组织面积等均使用Image-J软件测量完成, 木质部解剖图像见图 1。

|
| 图 1 树种木质部解剖图像 Fig. 1 Tree xylem anatomy image 图中红色为纤维组织, 蓝色为薄壁组织; 麻栎的最大导管直径、平均导管直径和导管壁厚度均最大, 臭椿、榆树、胡桃的轴向薄壁组织较多;白玉兰、鹅掌楸和紫荆的平均导管直径较小, 导管密度较大 |
根据导管横切面积与直径的关系计算出每个导管的直径D:
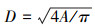
|
根据Hagen-Poiseuille方程计算潜在最大导水率Kp[28]:
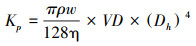
|
Kp单位是kg m-1 MPa-1 s-1, 其中ρw是纯水密度, 取998.2 kg/m3, η是粘滞系数, 取10-9 MPa·s, VD是导管密度。
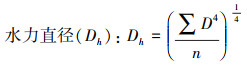
|
其中, D是导管直径, n是导管数量。

|
其中, t是相邻两个导管的导管壁厚度, b是较大导管的导管直径。
1.4 数据分析对树种间的性状进行单因素方差统计分析及Ducan法多重比较, 用独立样本t检验((Independent-Samples t-Test))分析抗旱树种和非抗旱树种间的木质部性状差异, 用Pearson相关分析抗旱树种、非抗旱树种及所有树种性状间的相关性。利用Origin 2018软件作图。
2 结果与分析 2.1 10个树种的木质部性状变异所研究的10个树种的16个木质部性状均有较大变异性。其中抗旱树种与木质部水力相关性状中潜在最大导水率的变异最大;薄壁组织性状中离管薄壁组织比例的变异最大;非结构性碳中可溶性糖的变异最大;非抗旱树种与木质部水力相关性状中Huber值变异最大;薄壁组织性状中离管薄壁组织比例的变异最大;非结构性碳中可溶性糖的变异最大。组成木质部的三大组织中导管占木质部横截面的比例为7.68%—25.64%, 平均值为17.93%±5.22%;薄壁组织为22.10%—38.55%, 平均值为34.45%±4.93%;纤维组织为43.23%—64.04%, 平均值为50.46%±6.80%。
2.2 抗旱树种与非抗旱树种非结构性碳(NSC)浓度的差异不同树种的木质部NSC浓度变异较大(变异系数为40.30%)。麻栎(Qa)的淀粉浓度在10个树种中最高, 达到(295.95±0.75)μg/mg, 是最小的白玉兰(Md)的5倍((58.20±0.91)μg/mg)。麻栎的可溶性糖浓度也高于其他所有树种((107.33±2.26)μg/mg)。抗旱树种的淀粉含量显著高于非抗旱树种(P < 0.01), 可溶性糖浓度无显著差异(P>0.05), 但抗旱树种的总NSC浓度显著高于非抗旱树种(P < 0.01)。

|
| 图 2 不同树种之间木质部非结构性碳差异 Fig. 2 Xylem Non-structural carbohydrate differences between different tree species |
麻栎(Qa)的平均导管直径和导管壁厚度都是最大, 分别为(119.71±21.171)μm, (4.57±0.713)μm;榆树(Up)作为抗旱树种中唯一的散孔材, 相较于其他环孔材抗旱树种, 其导管密度最大((168.76±26.552)μm), 而平均导管直径最小((25.59±5.249)μm)。总体比较, 抗旱树种的导管壁厚度、最大导管直径、旁管薄壁组织比例、轴向薄壁组织比例均显著大于非抗旱树种(P < 0.01);抗旱树种的平均导管直径、导管密度及其他性状与非抗旱树种并无显著差异(P>0.05)。
2.4 不同树种木质部性状的协同性分析10个树种的导管壁厚度与平均导管直径、最大导管直径和潜在最大导水率均显著相关, 5个抗旱树种间也存在这种关系, 但非抗旱树种不存在。5个抗旱树种和所有10个树种的最大导管直径与潜在最大导水率显著相关, 但非抗旱树种间不相关。抗旱树种总薄壁组织比例与轴向薄壁组织面积比例存在显著的正相关关系, 表明抗旱树种的薄壁组织主要由轴向薄壁组织主导, 5个非抗旱树种和总的10个树种间未表现出现此种关系;5个抗旱树种的导管连接度与Huber值存在显著的负相关关系, 而非抗旱树种无此种关系。
3 讨论Morris等对全球范围内2332种木本被子植物的薄壁组织比例、轴向薄壁组织比例、射线薄壁组织比例和平均导管直径进行分析, 发现总的薄壁组织变化范围为6.88%—64.20%, 轴向薄壁组织比例变化范围为0%—44.10%, 射线组织比例变化范围为5.23%—42.47%;导管平均直径变化范围为10%—435.12%[19]。本研究的10个树种的上述木质部性状均在此范围内(表 2、表 3)。
| 分类 Classification |
特征 Characteristics |
单位 Unit |
平均值 Mean |
变异系数 Coefficient of variation/% |
最大值 Max |
最小值 Min |
| 水力性状 | 潜在最大导水率 | Kg m-1 MPa-1s-1 | 66.51 | 80.35 | 162.64 | 15.06 |
| Hydraulic traits | 导管密度 | mm-2 | 75.81 | 71.43 | 168.76 | 15.80 |
| 导管平均直径 | μm | 60.12 | 57.66 | 119.71 | 25.59 | |
| Huber值 | mm2/cm2 | 0.01 | 40.73 | 0.02 | 0.01 | |
| 导管壁厚度 | μm | 3.13 | 25.91 | 4.57 | 2.55 | |
| 最大导管直径 | μm | 147.06 | 22.24 | 199.56 | 111.97 | |
| 导管连接度 | μm/μm | 0.12 | 20.61 | 0.16 | 0.09 | |
| 导管面积比例 | % | 17.37 | 20.47 | 20.94 | 12.25 | |
| 薄壁组织 | 离管薄壁组织比例 | % | 10.95 | 39.60 | 16.63 | 6.55 |
| Parenchyma | 旁管薄壁组织比例 | % | 7.10 | 37.60 | 11.00 | 3.04 |
| 轴向薄壁组织比例 | % | 18.05 | 17.83 | 21.80 | 13.84 | |
| 射线薄壁组织比例 | % | 14.28 | 16.44 | 18.71 | 12.66 | |
| 薄壁组织比例 | % | 32.33 | 14.26 | 38.55 | 26.50 | |
| 非结构性碳 | 可溶性糖浓度 | μg/mg | 0.70 | 43.38 | 1.07 | 0.32 |
| Non-structural carbohydrate | 淀粉浓度 | μg/mg | 2.18 | 23.93 | 2.96 | 1.45 |
| 总NSC浓度 | μg/mg | 2.88 | 22.57 | 4.03 | 2.28 |
| 分类 Classification |
特征 Characteristics |
单位 Unit |
平均值 Mean |
变异系数 CV/% |
最大值 Max |
最小值 Min |
| 水力性状 | Huber值 | mm2/cm2 | 0.02 | 58.05 | 0.03 | 0.01 |
| Hydraulic traits | 潜在最大导水率 | Kg m-1 MPa-1 s-1 | 15.18 | 45.54 | 23.14 | 6.63 |
| 导管密度 | mm-2 | 131.79 | 34.97 | 186.47 | 72.97 | |
| 导管面积比例 | % | 18.49 | 34.70 | 25.64 | 7.65 | |
| 导管面积比例 | % | 0.12 | 20.81 | 0.15 | 0.08 | |
| 最大导管直径 | μm | 72.22 | 18.29 | 85.36 | 51.91 | |
| 导管平均直径 | μm | 39.85 | 16.14 | 46.87 | 31.93 | |
| 导管壁厚度 | μm | 1.55 | 9.33 | 1.81 | 1.43 | |
| 薄壁组织 | 离管薄壁组织比例 | % | 5.56 | 67.46 | 9.80 | 0.86 |
| Parenchyma | 轴向薄壁组织比例 | % | 8.08 | 45.27 | 12.43 | 3.55 |
| 旁管薄壁组织比例 | % | 2.52 | 20.60 | 3.02 | 1.57 | |
| 射线薄壁组织比例 | % | 22.49 | 19.30 | 29.88 | 18.55 | |
| 薄壁组织比例 | % | 30.57 | 16.59 | 34.34 | 22.10 | |
| 非结构性碳 | 淀粉浓度 | μg/mg | 1.13 | 29.06 | 1.52 | 0.58 |
| Non-structural carbohydrate | 可溶性糖浓度 | μg/mg | 0.39 | 27.46 | 0.53 | 0.24 |
| 总NSC浓度 | μg/mg | 1.52 | 22.25 | 1.77 | 0.90 |
植物木质部空穴化引起的植物输水功能障碍可影响植物的水分平衡和气孔运动, 导致植物死亡[29], 因此植物的抗旱性在一定程度上可以由其抵抗栓塞发生的能力或栓塞后的修复能力决定[30]。抗旱性较强的植物一般具有较大的导管密度和较小的导管直径, 但本研究中抗旱树种的平均导管直径、Kp和导管密度与非抗旱树种相比并无显著差异(P>0.05), 抗旱树种的导管壁厚度和轴向薄壁组织均显著大于非抗旱树种(图 3, P < 0.01), 表明仅依靠导管直径和导管密度判断植物抗栓塞能力并不客观, 还应与导管纹膜孔特征和薄壁组织等木质部水力性状结合。本研究中抗旱树种最大导管直径与Kp存在显著正相关关系(R2=0.848), 而非抗旱树种的这种关系却并不明显(R2=0.430), 表明抗旱树种直径较大的导管具有更高的导水率。根据Hagen-Poiseuille定律, 当植物木质部导管直径较大时, 水分运输效率越高, 导水率越高[31]。本研究中树种的导管壁厚度与Kp、最大导管直径、平均导管直径均呈显著正相关关系(R2分别为0.554, 0.781, 0.727), 这一点在抗旱树种上的体现更为突出(图 4), 即本研究抗旱树种的导管直径较大时, 其导管有较厚的导管壁, 导管内径增加的同时需要有较强的机械支撑力才能保证较大口径的导管不易破裂, 导管壁厚度在一定程度上可以体现其机械支撑力[32], 厚的导管壁也影响导管纹孔形态, 尤其是纹孔膜厚度[10], 所以厚的导管壁保证了水分运输的安全性。本研究中的抗旱树种具有较高导水率的同时在一定程度上兼顾了安全性, 因此并未体现出水分运输高效性与安全性的权衡。

|
| 图 3 不同树种之间导管解剖特征差异 Fig. 3 Differences in vessel anatomy between different tree species |

|
| 图 4 不同树种木质部性状的协同性分析 Fig. 4 Synergistic analysis of xylem traits in different tree species 实线表明线性关系显著;虚线表明线性关系不显著;粗线表示所有树种指标间的关系; *:P < 0.05;**:P < 0.01 |
NSC为植物生长繁殖和新陈代谢提供能量, 并被认为对抵抗干扰和干扰后的恢复有重要作用[33], 主要是可溶性糖和淀粉。可溶性糖是植物在长期的干旱环境中重要的渗透调节物质, 是植物细胞中浓度较高的一大类物质[34], 有研究表明干旱胁迫会使淀粉也转化为可溶性糖[35-36]。本研究中抗旱树种的NSC浓度为(2.882±0.689)μg/mg, 显著高于非抗旱树种的(1.528±0.358)μg/mg, 表明具有更高的环境适应性以及较高的碳供应水平。Toshihiro等发现阔叶植物木质部栓塞修复能力要比针叶树植物强, 其原因就是阔叶植物木质部中有较多薄壁组织和NSC[37]。目前研究发现植物抗栓塞能力与植物茎干的水容存在相关关系, 木质部的导管通过吸收旁管薄壁组织中的水分或者离子, 改变导管内溶液的渗透势使水分向导管运输, 从而缓和导管内张力, 达到防栓塞的目的[38], 同时薄壁组织中含有大量水通道蛋白可以参与到木质部栓塞修复中[39]。本研究中的抗旱树种相较于非抗旱树种具有更多薄壁组织, 表明抗旱树种具备更强的栓塞修复能力。也有研究表明, 导管连接度越大, 植物木质部预防张力下导管破裂的能力越大[40]。Huber值表示维持单位叶面积水分供给所需的枝条边材投入[41], 一定程度反映植物枝条的输水效率。本研究抗旱树种的导管连接度与Huber值存在显著的负相关关系(R2=0.609), 导管连接度是导管壁厚度与导管直径的比值, 两类树种间导管直径无显著差异但抗旱树种的导管壁厚度显著大于非抗旱树种(图 3), 抗旱树种导管连接度与Huber值的这种关系或许表明导管壁更厚的树种容易维持更多的叶面积用来光合固碳。总体来说本研究的抗旱树种即通过厚的导管壁维持最大导管直径和水力效率, 同时又保证了抗栓塞的水力安全性, 并且抗旱树种具有更多的旁管薄壁组织和NSC也保证了足够的能量和资源进行栓塞修复。
麻栎作为干旱瘠薄山地绿化造林的先锋树种[42], 有最大的平均导管直径和最厚的导管壁, 反映了麻栎在水分运输高效性和安全性上有较好的协同关系。
4 结论本研究抗旱树种相较于非抗旱树种, 木质部NSC浓度更高、导管壁更厚、最大导管直径更大、旁管薄壁组织和轴向薄壁组织比例更大。抗旱树种直径较大的导管和较厚的导管壁既保证了较高的水分运输效率又具有一定的抗栓塞能力, 同时抗旱树种更多的旁管薄壁组织和NSC又为更强的贮水能力和栓塞修复能力提供了条件。本研究木质部导管解剖特征可为华北干旱瘠薄山区绿化造林树种的选择提供参考。
| [1] |
McDowell N, Pockman W T, Allen C D, Breshears D D, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams D G, Yepez E A. Mechanisms of plant survival and mortality during drought:why do some plants survive while others succumb to drought?. New Phytologist, 2008, 178(4): 719-739. DOI:10.1111/j.1469-8137.2008.02436.x |
| [2] |
王凯, 雷虹, 刘建华. 春季辽宁西北部主要绿化树种根叶抗旱生理性状评价. 应用生态学报, 2016, 27(6): 1853-1860. |
| [3] |
吴芹, 张光灿, 裴斌, 方立东. 3个树种对不同程度土壤干旱的生理生化响应. 生态学报, 2013, 33(12): 3648-3656. |
| [4] |
赵文达, 杨晓鹏, 孙铭, 张成林, 伍文丹, 马啸, 张新全. 不同扁穗雀麦种质苗期抗旱性鉴定与评价. 草业科学, 2018, 35(11): 2664-2671. DOI:10.11829/j.issn.1001-0629.2018-0013 |
| [5] |
张海娜, 鲁向晖, 金志农, 李阳, 王瑞峰, 李宗勋, 刘利昆. 高温条件下稀土尾砂干旱对4种植物生理特性的影响. 生态学报, 2019, 39(7): 2426-2434. |
| [6] |
蔡建国, 章毅, 孙欧文, 杨倩倩. 绣球抗旱性综合评价及指标体系构建. 应用生态学报, 2018, 29(10): 3175-3182. |
| [7] |
陈志成, 刘畅, 刘晓静, 万贤崇. 光强和树体大小对锐齿栎树木水、碳平衡的影响. 林业科学, 2017, 53(9): 18-25. |
| [8] |
Dixon H H, Joly J. On the ascent of sap. Philosophical Transactions of the Royal Society:Biological Sciences, 1895, 186: 563-576. |
| [9] |
Anderegg W R L, Klein T, Bartlett M, Sack L, Pellegrini A F A, Choat B, Jansen S. Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought-induced tree mortality across the globe. Proceedings of the National Academy of Sciences of the United States of America, 2016, 113(18): 5024-5038. DOI:10.1073/pnas.1525678113 |
| [10] |
Wheeler J K, Sperry J S, Hacke U G, Hoang N. Inter-vessel pitting and cavitation in woody Rosaceae and other vesselled plants:a basis for a safety versus efficiency trade-off in xylem transport. Plant, Cell & Environment, 2005, 28(6): 800-812. |
| [11] |
Sperry J S, Hacke U G, Pittermann J. Size and function in conifer tracheids and angiosperm vessels. American Journal of Botany, 2006, 93(10): 1490-1500. DOI:10.3732/ajb.93.10.1490 |
| [12] |
Cai J, Tyree M T. The impact of vessel size on vulnerability curves:data and models for within-species variability in saplings of aspen, Populus tremuloides Michx. Plant, Cell & Environment, 2010, 33(7): 1059-1069. |
| [13] |
Holbrook N M, Ahrens E T, Burns M J, Zwieniecki M A. In vivo observation of cavitation and embolism repair using magnetic resonance imaging. Plant Physiology, 2001, 126(1): 27-31. DOI:10.1104/pp.126.1.27 |
| [14] |
Berlyn G P. Plant structures:xylem structure and the ascent of sap. Science, 1983, 222(4623): 500-501. |
| [15] |
Larter M, Pfautsch S, Domec J C, Trueba S, Nagalingum N, Delzon S. Aridity drove the evolution of extreme embolism resistance and the radiation of conifer genus Callitris. New Phytologist, 2017, 215(1): 97-112. DOI:10.1111/nph.14545 |
| [16] |
Cosme L H M, Schietti J, Costa F R C, Oliveira R S. The importance of hydraulic architecture to the distribution patterns of trees in a central Amazonian forest. New Phytologist, 2017, 215(1): 113-125. DOI:10.1111/nph.14508 |
| [17] |
郝广友.水分条件迥异环境中同属或同种不同生活型热带木本植物的水分和光合生理生态比较研究[D].北京: 中国科学院研究生院, 2009.
|
| [18] |
Gleason S M, Westoby M, Jansen S, Choat B, Hacke U G, Pratt R B, Bhaskar R, Brodribb T J, Bucci S J, Cao K F, Cochard H, Delzon S, Domec J C, Fan Z X, Feild T S, Jacobsen A L, Johnson D M, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, McCulloh K A, Mencuccini M, Mitchell P J, Morris H, Nardini A, Pittermann J, Plavcová L, Schreiber S G, Sperry J S, Wright I J, Zanne A E. Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world's woody plant species. New Phytologist, 2016, 209(1): 123-136. DOI:10.1111/nph.13646 |
| [19] |
Morris H, Gillingham M A F, Plavcová L, Gleason S M, Olson M E, Coomes D A, Fichtler E, Klepsch M M, Martínez-Cabrera H I, McGlinn D J, Wheeler E A, Zheng J M, Ziemińska K, Jansen S. Vessel diameter is related to amount and spatial arrangement of axial parenchyma in woody angiosperms. Plant, Cell & Environment, 2018, 41(1): 245-260. |
| [20] |
杨吉华, 张永涛, 孙明高, 石浩来, 刘春生. 石灰岩丘陵土壤旱作保水技术的研究. 水土保持学报, 2000, 14(3): 62-66. DOI:10.3321/j.issn:1009-2242.2000.03.013 |
| [21] |
杨程. 华北石质山区植被恢复途径的探讨. 长春大学学报, 2008, 18(3): 95-99. DOI:10.3969/j.issn.1009-3907-B.2008.03.025 |
| [22] |
陈志成. 8个树种对干旱胁迫的生理响应及抗旱性评价[D].泰安: 山东农业大学, 2013.
|
| [23] |
叶权平, 张文辉, 于世川, 薛文艳. 桥山林区麻栎群落主要乔木种群的种间联结性. 生态学报, 2018, 38(9): 3165-3174. |
| [24] |
韩东, 王浩舟, 郑邦友, 王锋. 基于无人机和决策树算法的榆树疏林草原植被类型划分和覆盖度生长季动态估计. 生态学报, 2018, 38(18): 6655-6663. |
| [25] |
徐嘉娟, 李火根. 鹅掌楸LcPAT8基因的克隆及功能初步分析. 林业科学, 2017, 53(9): 45-54. |
| [26] |
Hoch G, Popp M, Körner C. Altitudinal increase of mobile carbon pools in Pinus cembra suggests sink limitation of growth at the Swiss treeline. Oikos, 2002, 98(3): 361-374. DOI:10.1034/j.1600-0706.2002.980301.x |
| [27] |
王学奎. 植物生理生化实验原理和技术 ((第二版)). 高等教育出版社, 2006: 75-77.
|
| [28] |
Poorter L, McDonald I, Alarcón A, Fichtler E, Licona J C, Peña-Claros M, Sterck F, Villegas Z, Sass-Klaassen U. The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species. New Phytologist, 2010, 185(2): 481-492. DOI:10.1111/j.1469-8137.2009.03092.x |
| [29] |
Nardini A, Gullo M A L, Salleo S. Refilling embolized xylem conduits:is it a matter of phloem unloading?. Plant Science, 2011, 180(4): 604-611. DOI:10.1016/j.plantsci.2010.12.011 |
| [30] |
Schoonmaker A L, Hacke U G, Landhäusser S M, Lieffers V J, Tyree M T. Hydraulic acclimation to shading in boreal conifers of varying shade tolerance. Plant, Cell & Environment, 2010, 33(3): 382-393. |
| [31] |
Stuart S A, Choat B, Martin K C, Holbrook N M, Ball M C. The role of freezing in setting the latitudinal limits of mangrove forests. New Phytologist, 2007, 173(3): 576-583. DOI:10.1111/j.1469-8137.2006.01938.x |
| [32] |
徐茜, 陈亚宁. 胡杨茎木质部解剖结构与水力特性对干旱胁迫处理的响应. 中国生态农业学报, 2012, 20(8): 1059-1065. |
| [33] |
Poorter L, Kitajima K. Carbohydrate storage and light requirements of tropical moist and dry forest tree species. Ecology, 2007, 88(4): 1000-1011. DOI:10.1890/06-0984 |
| [34] |
Zeppel M J B, Harrison S P, Adams H D, Kelley D I, Li G Q, Tissue D T, Dawson T E, Fensham R, Medlyn B E, Palmer A, West A G, McDowell N G. Drought and resprouting plants. New Phytologist, 2015, 206(2): 583-589. DOI:10.1111/nph.13205 |
| [35] |
Mitchell P J, O'Grady A P, Tissue D T, White D A, Ottenschlaeger M L, Pinkard E A. Drought response strategies define the relative contributions of hydraulic dysfunction and carbohydrate depletion during tree mortality. New Phytologist, 2013, 197(3): 862-872. DOI:10.1111/nph.12064 |
| [36] |
Hoch G, Körner C. The carbon charging of pines at the climatic treeline:a global comparison. Oecologia, 2003, 135(1): 10-21. DOI:10.1007/s00442-002-1154-7 |
| [37] |
Umebayashi T, Morita T, Utsumi Y, Kusumoto D, Yasuda Y, Haishi T, Fukuda K. Spatial distribution of xylem embolisms in the stems of Pinus thunbergii at the threshold of fatal drought stress. Tree Physiology, 2016, 36(10): 1210-1218. DOI:10.1093/treephys/tpw050 |
| [38] |
Nardini A, Salleo S, Jansen S. More than just a vulnerable pipeline:xylem physiology in the light of ion-mediated regulation of plant water transport. Journal of Experimental Botany, 2011, 62(14): 4701-4719. DOI:10.1093/jxb/err208 |
| [39] |
冷华妮.植物栓塞修复机制与质膜内在水通道蛋白基因的克隆、表达和转基因研究[D].北京: 中国林业科学研究院, 2012.
|
| [40] |
李荣.耐旱树种木质部结构与耐旱性关系研究[D].杨凌: 西北农林科技大学, 2016.
|
| [41] |
赵延涛, 许洺山, 张志浩, 周刘丽, Arshad A, 张晴晴, 宋彦君, 阎恩荣. 浙江天童常绿阔叶林不同演替阶段木本植物的水力结构特征. 植物生态学报, 2016, 40(2): 116-126. |
| [42] |
张永涛, 杨吉华, 慕宗昭. 山东退化山地立地分类体系构建及造林模型研究与应用. 北京: 电子工业出版社, 2016: 10-18.
|