 2020, Vol. 40
2020, Vol. 40文章信息
- 何念鹏, 刘聪聪, 徐丽, 于贵瑞
- HE Nianpeng, LIU Congcong, XU Li, YU Guirui
- 生态系统性状对宏生态研究的启示与挑战
- Ecosystem traits open new insights into macroecology: opportunity and challenge
- 生态学报. 2020, 40(8): 2507-2522
- Acta Ecologica Sinica. 2020, 40(8): 2507-2522
- http://dx.doi.org/10.5846/stxb201903190515
-
文章历史
- 收稿日期: 2019-03-19
- 网络出版日期: 2019-12-31

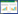




2. 中国科学院大学资源与环境学院, 北京 100049;
3. 东北师范大学草地科学研究所, 植被生态科学教育部重点实验室, 长春 130024
2. College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China;
3. Key Laboratory of Vegetation Ecology, Ministry of Education, Institute of Grassland Science, Northeast Normal University, Changchun 130024, China
生态系统是指在一定空间内生物与环境构成的统一整体, 在该系统中生物与生物、生物与环境间相互影响和相互制约, 并在一定时期内处于相对稳定的动态平衡状态[1]。生态系统是当前生态学与地学研究的重要对象, 是群落生态学和宏生态学最重要的中间环节、且与人们所需的物质和能量来源、生存环境息息相关, 因此其研究内容与范畴受到公众和政府的广泛关注。在传统研究中, 科学家非常重视生态系统组成(非生物的物质和能量、生产者、消费者、分解者等)、生态系统营养结构(食物链和食物网)和生态系统功能(物质循环、能量流动和信息传递), 并取得了令人瞩目成绩[2-3]。科学家围绕“生态系统结构与功能的关系、及其对外界干扰的响应和适应”开展了大量研究工作, 其中生物多样性与生态系统功能实验(Biodiversity and Ecosystem Functioning, BEF)是最经典研究案例[4-7]。近年来, 越来越多的研究发现影响生态系统功能的不仅是物种丰富度, 而且还有群落功能性状的多样性[8-10]。目前功能性状被广泛地应用于植物和动物个体水平适应与生产力优化机制研究, 但在群落和生态系统尺度研究中科学家却极少提及性状, 是因为在群落或生态系统性状没有意义吗?
性状定义起源于遗传学或生理学, 它是具有明显的、可遗传可测定的且有一定生理生态意义的特征参数[11-12]。性状在物种水平上被广泛应用, 却极少将其拓展到群落和生态系统尺度, 这似乎是生态学科一个很大难题[13-14]。生态系统是一个复杂系统, 复杂的自组织功能使其结构和功能达到最优化, 以更好地适应环境和抵抗外界干扰, 但如何实现这种适应性和抗性?个体水平的适应是基础, 群落水平的适应与优化机制可能也非常重要[15]。因此, 我们有必要从群落尺度和生态系统尺度探讨功能性状时空变异规律及其生态意义, 从基础概念与理论框架寻求突破以完善生态学研究体系, 更好地推动相关研究领域的发展。
1 发展“生态系统性状”的迫切需求和理论基础生态系统性状是生态系统的基本属性, 并持续地发挥着其功能, 具体表现为生态系统的环境适应性。从人类诞生之初, 就开始利用、学习和改造自然, 并逐步意识到植物性状是植物对环境长期适应与进化的结果, 并人为操控或筛选特定性状来实现农作物的高产优质。Violle等[15]在Oikos发表的综述明确指出(图 1):在器官-物种-种群-群落-生态系统水平上, 性状都具有其特定的适应或功能优化的意义, 并正式提出功能性状的概念(From trait to functional trait)。该论文在发表10年内, 引用已超过1100次, 可见其新理念深受大家认可。虽然它没有明确给出生态系统性状的定义, 却启发我们大胆地推测:“作为一个鲜活的有机系统, 生态系统是通过不断调节其结构或组成、辅以植物、动物和微生物功能性状的适应与演化, 达到生态系统功能的优化并适应特定的环境”。即在生态系统尺度, 性状对生态系统功能优化与环境适应也具有非常重要的作用。

发展生态系统尺度的性状是生态系统生态学自身发展的迫切需求。目前, 功能性状在器官、个体、种群水平的生态意义和功能已被大家所熟知和接受[13]。然而, 受性状定义的约束和人们认识的局限性, 当前绝大多数性状研究都局限于器官、个体或种群水平(图 2), 近年来科学家通过物种丰富度等简单加权方式将性状拓展到群落尺度[16-17], 但由于概念的限制, 生态系统尺度的性状研究相对较少。此外, 已开展的群落、生态系统、区域甚至全球尺度性状研究工作, 科学家们也都是采用直接算数平均法来进行尺度推导[18], 落入了“物种水平简单算数平均=群落”的陷阱;然而, 自然群落尤其是天然森林群落, 其结构和组成非常复杂, 不同区域森林结构和组成存在很大差异, 简单算术平均可能会对相关研究结论造成很大影响, 研究结论的科学性和准确性有待商榷[17, 19-21]。因此, 我们必须跳出“物种水平简单算术平均=群落”的陷阱, 使我们在群落和生态系统尺度的性状研究真正能接近自然、接近真实。这迫切需要破除传统概念的束缚, 发展在群落或生态系统尺度上的性状新概念、内涵、科学研究方法等。

|
| 图 2 植物性状研究的尺度拓展现状 Fig. 2 The current status of scaling-up for the studies of plant traits |
此外, 发展生态系统水平性状能更好地促使当前高速发展新技术(遥感观测、通量观测、模型模拟)在生态系统生态学研究中的运用, 并为传统性状研究与宏生态学搭建一座新桥梁。生态系统生态学研究若能借用上当前高速发展的新技术, 可有力促进其自身研究手段和研究深度的快速发展。在遥感技术高速发展的现实背景下, 其观测的部分参数如叶面积指数、比叶面积、叶氮含量、光谱特征等, 本身就是或非常接近生态系统性状, 因此这些新技术和新参数可为生态系统生态学研究提供大量新数据和新思路。除此之外, 生态系统性状有助于传统性状研究的成果真正服务于宏生态学, 实现从器官水平拓展到生态系统水平的美好愿景, 拓宽传统性状研究的应用范畴, 促进性状研究自身的发展。当然, 群落水平或生态系统水平的性状参数能为遥感观测、通量观测、模型模拟提供验证、参数优化、结果比对, 也能很大程度上提高人们对宏生态研究的精度和深度(图 3)。众所周知, 当前人类社会所面临的生态环境问题, 绝大多数都是需要在生态系统尺度、流域尺度、区域尺度、甚至全球尺度来解决。因此, 发展生态系统水平的性状概念、方法和技术, 可实现性状研究、生态系统生态学研究、宏生态学研究(或宏观地学研究)的多赢, 不仅是学科发展的迫切需求, 更是社会发展的迫切需求。

|
| 图 3 传统性状与宏生态研究相联系的美好愿景和主要障碍 Fig. 3 The proposed bridge and barriers between traditional traits and macro-ecological researches |
总之, 生态学研究正在快速向宏观方向发展, 而生态系统是宏观研究的最基本尺度之一;此外, 全球变化对陆地生态系统影响的评估也多是以生态系统为主要对象。因此, 获得基于单位土地面积的生态系统性状, 有助于在空间尺度匹配(或量纲匹配)的前提下实现个体水平测定的植物性状数据与生态模型和遥感观测的有效关联, 也有助于更好地探讨区域尺度下自然生态系统结构和功能的关系及其对全球变化的响应与适应(图 3)。植物性状研究的空间壁垒和宏生态学发展的现实需求, 促使我们重新思考植物性状研究的尺度问题;需要将植物性状统一到单位土地面积尺度, 才能与宏生态学研究的主要观测手段紧密联系起来(通量观测、模型模拟和遥感观测), 推动相关学科发展。
2 生态系统性状的定义、内涵及研究案例 2.1 生态系统性状定义为了破解上述难题或疑惑, 我们团队最近发展了生态系统性状(Ecosystem traits, ESTs)的概念体系[22]。在此, 生态系统性状被定义为在群落或生态系统尺度能被单位土地面积标准化的、能体现生物(植物、动物、微生物)对环境适应、繁衍和生产力优化的任何可量度的性状(以强度或密度形式呈现)。对任何特定的生态系统而言, 生态系统性状均由一系列植物群落性状、动物群落性状、微生物群落性状、土壤性状等共同组成, 不同性状间均在群落或生态系统尺度起特定的作用或相互作用(图 4);即群落性状是生态系统性状研究的核心单元。生态系统性状的核心内涵包括:(1)任何生态系统性状均以群落为单位被转化为以单位土地面积为基数的性状, 如叶片面积经尺度拓展为叶面积指数、叶片干重经尺度拓展为叶生物量、叶片气孔密度经尺度拓展为单位土地面积的气孔个数、叶片C、N和P含量经拓展为C密度、N密度和P密度等;(2)任何生态系统性状均是可测量或可推导, 原则上均是采用严格的群落生物量(或结合叶片比叶面积)加权法推导;(3)任何生态系统性状应能从不同层面反映生物(植物群落、动物群落、微生物群落等)对环境的适应、繁衍或生产力优化, 即具有明确的生态学意义。

|
| 图 4 在群落尺度用单位土地面积标准化后生态系统性状的抽象示意图 Fig. 4 The logistic frames of ecosystem traits being standardized as land area on the levels of communities |
如何将器官水平测定的性状推导到群落, 是科研人员面临的一个巨大挑战。根据拟解决问题的不同, 前人发展了三种尺度拓展方法:(1)不考虑群落结构的直接算术平均(公式1)[23-24]。该方法在性状研究中被广泛使用, 尤其在全国、洲际或全球的大尺度性状整合分析中被广泛使用。(2)考虑群落结构后, 将不同物种相对丰富度或相对重要值作为性状推导的权重系数(公式2)[15]。(3)考虑群落结构后, 在物种和性状不匹配情景下对群落加权平均值的估算方法(公式3)[25]。上述三种方法都可以看成是对物种水平的平均, 应用于探讨群落或生态系统更高层次的过程和相互关系, 尤其是用于探讨生态学中的个体-种群间相互作用、竞争与共存等。但仔细分析, 不难发现公式1—3并未对量纲进行转换(如碳、氮、磷含量的单位仍然是g/kg; 比叶面积的单位mm2/mg;叶片气孔密度的单位number/cm2)。然而, 生态系统尺度的功能, 几乎都是基于单位土地面积(或空间)来进行测定和模拟(具体途径主要是通量观测、模型模拟、遥感观测)。因此, 公式1—3无法解决“量纲不匹配”和“尺度不匹配”的问题。
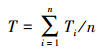
|
(1) |
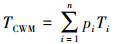
|
(2) |
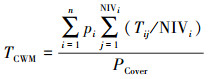
|
(3) |
式中, T或者TCWM代表群落性状值, 是一种均值的概念, CWM(Community weighted means)代表群落相对生物量(或相对丰富度, 重要值等)加权的方法, pi代表相对生物量(或相对丰富度, 重要值等), Ti代表群落中第i物种的性状值, n代表已知物种数量(公式1)或群落中物种数量(公式2)或群落中能与数据库中性状匹配的物种数量(公式3), Tij代表第i物种在已有数据库中第j个性状值, NIVi代表第i物种性状值在已有数据库中的重复数(≥1), PCover代表群落中能与数据库中性状匹配的物种累计相对生物量(或相对丰富度, 重要值等)[25]。根据生态系统性状的定义和内涵, 生态系统性状与性状、功能性状以及上述三个方程推导的传统的群落性状具有本质的差异(表 1)。
| 术语 Term |
定义 Definition |
与宏生态学匹配性 Compatibility with macroecology |
尺度 Scale |
参考文献 References |
|
| 量纲 Dimension |
空间尺度 Spatial scale |
||||
| 性状 Trait |
生物体相对稳定且可量度的所有特征参数, 包括形态、结构、生理、节律和行为方式等 | 否 | 否 | 个体或物种 | [11, 12] |
| 功能性状 Functional trait |
影响生物个体生长、繁殖和生存、或可直接或间接影响其适应度的具有生态意义参数 | 否 | 否 | 个体或物种 | [15] |
| 群落性状 Community trait |
“传统”群落性状是群落内物种特定性状通过算术平均值或根据物种丰富度、重要值等推导的加权平均值(未进行量纲或空间尺度转换) | 否 | 否 | 群落 | [25] |
| 群落性状是指在群落尺度上充分考虑群落结构、物种贡献和面积、并能被单位土地面积标准化的、能体现生物对环境适应、繁衍和生产力优化的任何可量度的性状(以强度或密度形式呈现, 完成了量纲和空间尺度转换) | 是 | 是 | 群落 | [22] | |
| 生态系统性状 Ecosystem trait |
在群落或生态系统尺度, 充分考虑群落结构、物种贡献和面积、并能被单位土地面积标准化的、能体现生物对环境适应、繁衍和生产力优化的任何可量度的性状(以强度或密度形式呈现);生态系统性状是由一系列植物、动物、微生物等群落性状组成, 其基本研究的核心单元是群落性状 | 是 | 是 | 生态系统 | [22] |
由此可见, 如何将器官水平测定的性状科学地推导到群落或生态系统水平、并与自然群落或生态系统功能相匹配仍然是一个巨大挑战。为了破解该重大技术难题, 何念鹏团队在借用群落结构数据、每个物种比叶面积数据和异速生长方程数据等, 发展了新的生态系统性状推导方法, 并能将其标准化为单位土地面积上的群落性状, 公式4和公式5分别为计算质量标准化和叶片面积标准化的性状[22, 26], 这两个公式既考虑了复杂的群落结构、又实现了生态系统性状向单位土地面积转换的目的;因此, 他们不仅可以用来探讨传统生态系统生态学中植物、动物、微生物间、生物与非生物要素间的相互关系或相互作用, 还可与生态系统功能相联系更好地探讨生态系统水平性状与功能的关系及其影响机制(图 5)。
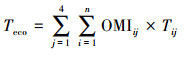
|
(4) |
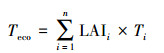
|
(5) |

|
| 图 5 生态系统性状构建传统性状与宏生态的桥梁与主要途径 Fig. 5 Ecosystem traits can bridge traditional traits to macroecology researches through a series of scaling-up and scaling-matching C:碳, Carbon; N:氮, Nitrogen; P:磷, Phosphorus; GPP:总初级生产力, Gross primary productivity; NPP:净初级生产力, Net primary productivity;ET:生态系统蒸散, Ecosystem transpiration; WUE:水分利用效率, Water use efficiency |
公式中, 性状分为质量标准化的性状(如单位叶片质量上的N含量)和面积标准化的性状(气孔密度, 单位叶片面积上的气孔数量)。公式4用于推导质量标准化的性状, 公式5用于推导叶片面积标准化的性状。Teco代表性状T的生态系统水平性状值, 是一种累加的概念。n代表群落中的物种数量, OMIij代表群落中第i物种第j器官(根-茎-叶-枝)质量指数, LAIi代表群落中第i物种的叶面积指数, 其算法是根据i物种的胸径、树高和一元或二元生长方程推导出该物种各器官的生物量, 并根据比叶面积SLA(Specific leaf area)将叶片生物量转为叶片面积, 并将器官生物量或者叶片面积标准化在单位土地面积上。Ti或Tij代表第i个物种的性状值或第i个物种在j器官上的性状值。在公式4中, 若性状在根-茎-叶-枝器官上不连续, 则可忽略该器官的性状值的计算或归为0值, 也可分别计算某一层次或亚层次(微层次)的生态系统性状。若性状在植物-动物-微生物-土壤中是连续的(如C含量), 可以根据实际研究目进行推导。
生态系统性状从方法学上来说是一个尺度转换或推绎问题, 也就是如何把器官、个体水平的性状转换到单位土地面积上群落水平(动物群落性状、植物群落性状、微生物群落性状等);因此, 要进一步发展生态系统性状研究的方法学框架, 尺度转换是关键突破点。新概念体系下的生态性状与传统群落水平的性状既有本质区别、也存在内在联系。主要区别在于, 第一, 概念上生态系统性状是一种基于单位面积的累加概念, 而传统群落水平的性状基于测定数据是平均值概念, 二者拟回答的科学问题不同。第二, 生态系统性状需要更系统的数据, 如准确匹配的群落结构-物种组成-性状值-生物量方程, 而传统群落水平的性状可以通过丰富度、重要值等进行估计。二者的内在联系在于, 若传统群落水平的性状也能达到精准的数据要求, 则生态系统性状与传统群落水平的性状可以通过叶面积(或器官质量)指数等重要参数进行转换。生态系统性状不是局限于单位土地面积上的性状, 也可以是由两个或多个生态系统性状派生出的新性状, 如N:P(单位土地面积上的N含量和P含量的比值), 比叶面积SLA(单位土地面积上的叶片面积和叶片生物量的比值), 这种派生的生态系统性状与传统群落水平的性状在数值上相同。相比于传统性状, 生态系统性状可能在解释生态系统功能上更具有优势, 以生态系统生产力(Gross primary productivity, GPP)为例, GPP是以单位土地面积进行量化, 生态系统性状也是以单位土地面积进行量化, 从而克服了二者在量纲上的不匹配性。
2.3 生态系统性状研究的案例 2.3.1 数据源要求根据生态系统性状的定义和内涵, 需要将传统的器官水平测定的性状科学地推导至基于单位土地面积的群落或者生态系统水平。因此, 它对数据源的系统性要求非常高。以植物性状为例, 为了实现器官水平测定的叶绿素含量、氮和磷含量的尺度外推, 理论上需要详细的群落结构数据、每个物种的比叶面积数据、每个物种的叶绿素、氮和磷含量数据;当然, 不同的参数推导过程中对数据的系统要求会有所差异。然而, 现有的大尺度性状数据库, 都是采用收集公开数据方法构建, 其主体数据集中为植物叶片易于测定的性状(叶片大小、厚度、比叶面积、碳、氮含量等)和植物个体大小等[27], 叶片常规形态性状-气孔性状-解剖结构性状-叶绿素含量-元素含量等配套数据几乎没有。如全球著名植物性状数据库(Plant Trait database, TRY)收录了148000个物种的叶片性状数据, 具有600多万条性状记录(https://www.try-db.org);但TRY却非常缺乏叶-枝-根-干的配套性状数据以及群落结构、土壤微生物、土壤质地等配套数据。由于上述局限性, 使基于TRY数据库的所有分析均在个体水平开展(包括与TRY相似的收集数据的相关研究), 难以满足生态系统性状推导的需求[22, 28]。
提出了“系统性+符合自然”的新思路, 并建议在新思路下建设与TRY截然不同的新型数据库。新型数据库能解决先前数据库(含TRY和类似简单测定或收集型数据库)无法解决的科学问题(1)所测定的指标体系能更接近复杂的天然森林生态系统和草地生态系统;(2)能实现从物种水平(或器官水平)尺度上推至群落水平和生态系统水平, 并探讨自然群落水平以及生态系统水平性状与功能的定量关系。为了尽量降低人为干扰, 以典型森林生态系统的系统性性状调查为例, 应将实验样地设置于自然保护区内或野外生态站的长期监测样地内, 在每种森林生态系统的典型地段, 分别设置4个30 m×40 m乔木调查样方、每个乔木样方内分别设置2个5 m×5 m灌木调查样方和4个1 m×1 m草本调查样方, 并按中国生态系统研究网络(China Ecosystem Research Network, CERN)调查规范, 采用每木检尺详细调查乔、灌、草的群落结构特征。样品的采集包括植物和土壤样品等。
植物样品采集:①乔木样品采集:选择样地内(或周边)长势良好的乔木植株, 采用吊塔+人工爬树+高枝剪相结合模式获得顶层向阳树枝;随后手工摘取无病虫害的、完全展开的健康叶片, 每个样地分别形成叶或细枝混合样品。在叶片样品采集的同时, 采用生长锥采集树芯样品;同时, 沿侧根挖掘出完成的细根根系(从1级到5级根)。②灌木样品采集:对每个灌木物种, 在每个样地选择1—3株健康植株, 利用高枝剪或枝剪截取冠层顶端枝条, 分别获得叶和枝条的混合样品;同时采用挖掘法获得灌木根系。③草本样品采集:在每个样地内, 每个草本物种选取5—30株, 采集地上部分(叶、茎)和根系。此外, 在草本样方内收集地表凋落物混合样品。土壤样品采集:利用6 cm直径的土钻, 分0—10 cm、10—30 cm、30—50 cm、50—100 cm四层进行土壤取样。在每个样地取30—40钻土壤样品, 获得每层土壤样品>10 kg;随后, 大约4 kg鲜土壤快速冷藏于-20 ℃冰柜, 其他样品遮荫风干。所测定指标包括叶-枝-干-根性状(如叶片性状包括常规形态、气孔、叶片结构、化学元素、叶绿素含量、非结构性碳水化合物、叶片热值等), 土壤微生物组成和酶活性、土壤有机质含量及其组分等[19];基于上述标准, 在中国45个典型森林生态系统开展了系统性的调查与测试工作。

|
| 图 6 中国陆地生态系统性状数据库的野外调查样点空间分布 Fig. 6 The spatial distribution of field investigation for Functional Trait Database in China (China_Traits) |
类似地, 在内蒙古高原、黄土高原和西藏高原典型草地分布区, 设置了30个典型草地生态系统, 开展了类似于森林生态系统性状的详细调查采样工作。在草地调查中, 还增加了土壤小型动物(线虫、跳虫、蚯蚓等)、蝗虫等调查工作。以此为基础, 初步建成了中国生态系统性状数据库(Functional Trait Database in China, China_Traits)。目前, 该数据库包含40多个典型森林生态系统4700多种植物叶-枝-干-根的性状数据、30多个典型草地生态系统800多种植物叶-茎(干)-根的性状数据、配套的群落结构、气候、土壤、土壤微生物、生态系统总初级生产力、生态系统净初级生产力等参数。China_Traits具有系统性和匹配性的优点, 并具有从器官-物种-群落-生态系统-区域的纵深分析能力, 与著名的全球植物性状数据库(Plant Trait database, TRY)形成了鲜明的对比(图 7)。从更深入揭示自然生态系统而言, 人们需要具有开展多维度性状变化及其影响因素的基础数据;因此, China_Traits数据库具有TRY数据库无法比拟的优势, 其“系统性+符合自然”理念将是未来相关数据库建设的重要原则。

|
| 图 7 中国生态系统性状数据库的特点及其潜在创新性的应用范畴 Fig. 7 Comparison between Functional Trait Database in China (China_Traits) and Plant Trait Database (TRY) in view of their data composition and application TRY:植物性状数据库, Plant Trait Database |
研究人员利用中国东部南北样带的9个典型森林生态系统的详细调查数据, 利用公式4和5的方法, 完成一系列植物性状参数从器官水平到群落水平的推导, 获得了相应的生态系统性状[22]。已完成推导的具体参数包括叶片常规形态特征、叶绿素含量、叶片非结构性碳水化合物、叶片气孔特征、叶片解剖结构特征、植物叶-枝-干-根的碳氮磷含量等, 并从器官-物种-功能群-群落-生态系统角度探讨这些性状的纬度变异规律和主要影响因素[16-17, 19-20, 29-30]。除了叶绿素含量、叶片非结构性碳水化合物、叶片气孔特征、叶片解剖结构特征大尺度空间变异特征属全球首次报道外, 还系统性地将性状研究从传统的器官尺度推导到了群落或生态系统尺度, 为生态系统性状研究提供了可复制的案例与方法学依据, 开拓了植物群落性状研究的新领域。
基于实测数据所推导的生态系统性状, 在天然森林生态系统中建立了(多个)植物群落性状与生产力的定量关系, 为生态系统性状与功能研究提供了有力的研究范例, 也奠定了重要理论基础。长期以来, 由于缺乏系统性的性状调查数据, 传统的“功能性状-功能关系”研究都局限在植物个体水平, 或通过控制实验进行, 其结论未得到大尺度的天然森林群落数据的验证;此外以“个体性状=群落性状”的方式进行尺度拓展难以适用于复杂的天然森林群落[25, 28]。因此, 如何建立功能性状与生产力的关系成为功能生态学领域研究的世界性难题[31]。利用详细的调查数据和“群落结构+异速生长方程+比面积法”方法, 突破了从器官-功能群-群落推导的技术难题, 并发现植物群落水平的气孔密度能解释水分利用效率51%的空间变异[20];类似地, 从比叶面积、叶片解剖结构、叶绿素含量角度分别建立了其与GPP的定量关系[19, 30], 为天然群落植物性状与功能定量关系的研究提供了可借鉴范例。虽然多个植物功能性状与GPP都显著相关, 但它们单独解释度却都不高, 因此有关“植物叶片多个性状协同是植物生产力优化重要机制”的假设是合理的, 但不能盲目地夸大单一性状的重要性。
2.3.3 生态系统性状在全国尺度的研究案例宏生态学主要技术手段主要是在流域或区域尺度开展, 能否提供流域或区域尺度与之匹配的基于地面测定的生态系统性状, 将是拓展性状研究领域或应用的重要基础。因此, 除了在样地尺度完成生态系统性状的推导并建立群落性状与生产力、水分利用效率定量关系外, 也以氮为例开展了全国尺度的生态系统性状推导与研究(生态系统性状之氮)。
生态系统的氮含量及其分配, 是陆地生态系统重要的研究内容, 也是生态系统性状的重要参数。利用前期收集的16000多个森林和草地地上生物量数据、15000多个森林和草地地下生物量数据, 结合China_Trait数据库中5000多个物种叶-枝-干-根氮含量数据, 以及群落结构基础上推导的15000多个样地的叶-枝-干-根性状参数, 在此基础上利用随机森林模型+土地覆被数据生产了中国陆地生态系统叶、枝、干、根、0—20 cm土壤和20—100 cm土壤的氮密度性状数据(基于单位面积)。有了这些数据, 就可能给研究提供新的分析角度, 例如探讨其储量和空间变异格局, 生态系统内氮分配规律、氮含量与GPP和NPP间的关系、如何将生态系统性状之氮整合进入生态系统过程模型等。随着类似的生态系统性状参数或应用案例的增多, 不仅有助于我们深入地开展生态系统生态学的研究, 也有助于架起传统性状与宏生态学的桥梁, 更好地解决当前面临的生态环境问题。
3 生态系统性状对宏生态研究的启示与挑战生态系统性状概念体系的提出与发展, 能够促进生态系统生态学和宏生态学(宏观地学)的快速发展, 具体体现在以下几个方面:
3.1 发展以性状为基础的生态系统生态学新研究框架生态系统结构通常是指生态系统内植物、动物、微生物、土壤等组成要素的种类、大小、水平位置、空间位置、或者是营养级位置、食物网位置;生态系统功能是指生态系统总初级生产力、次级生产力、净初级生产力、养分利用效率、水分利用效率、固碳能力、水土保持能力等。由于生态系统性状概念的缺失, 以往对性状的研究大多数均是在器官或个体水平进行, 因此人们常不将其与结构和功能并行, 从而产生了“结构-过程-功能”的经典框架(图 8)。科研人员围绕生态系统结构-功能关系及其对外界干扰的适应与响应等开展了大量研究工作, 并试图建立生态系统结构和功能的定量关系并用于指导生产实践。最经典研究案例为生物多样性与BEF, 据不完全统计已经发表了近1.8万篇相关论文, 但依然未从理论上和实验上获得完全一致的研究结论[4];科研人员只能用抽样效应与物种性状变异等来解释这些差异。生态系统性状概念体系的提出, 将构建以生态系统性状为基础的生态系统生态学新研究框架, 为解决当前生态系统生态学研究中许多难题提供新的探索途径。

|
| 图 8 构建以性状为基础的生态系统生态学新研究框架 Fig. 8 Development of new theoretical frames for trait-based ecosystem ecology |
此外, 作为一个复杂系统, 生态系统具有复杂的自组织能力使其结构和功能达到最优化, 更好地适应环境或抵抗外界干扰, 甚至具有可再生能力;主要通过“结构-性状-功能”三维结构来共同实现的。如前面所讨论的一样, 生态系统结构本身难以直接实现功能或体现对环境的适应, 但结构可以通过调节物种组成(物种具有其特定的性状变异和适应范围), 进而通过调节群落整体的性状特征来影响生态系统功能和适应性。Violle等[15]已经明确提出了从器官-生态系统整体体系中, 性状都在其适应环境和生产力优化方面扮演着重要的角色;随着生态系统性状概念体系、推导方法和可利用数据源的日益增加, 必将促使生态系统生态学新研究框架的发展。
3.2 丰富生态系统生态学的研究思路和技术途径生态系统性状概念体系的提出和发展, 为在生态系统尺度深入探讨植物、动物、土壤微生物性状的内部关系、协同或趋异规律提供了新思路, 也可更好地探讨植物-动物-土壤微生物-土壤和气候等的相互作用关系、并从性状角度揭示植物群落、动物群落和土壤微生物群落的构建与维持机制。理论上讲, 生态系统生态学研究应基于群落和生态系统尺度的数据来开展相关研究, 而不是器官、物种或种群尺度;然而, 受测试技术与理念的限制, 从生态系统尺度开展植物群落-动物群落-土壤微生物群落-土壤和气候相互关系的研究却十分欠缺。绝大多数研究报道都是局限于特定的种类(植物、动物、土壤微生物)与其他生物或环境要素的影响, 严格意义上说不是真正的生态系统生态学研究范畴(种群生态或群落生态学)。生态系统性状概念体系及其新研究框架, 将在相对统一的空间尺度和量纲上获得植物群落性状、动物群落性状、土壤微生物群落性状和土壤属性等, 为深入研究生态系统尺度生物-生物、生物-非生物性状间的关系奠定坚实的理论基础(图 4)。
另一方面, 生态系统性状是一系列基于单位土地面积群落性状的组合, 其可以很好地解决长期以来(植物、动物)性状数据与宏观尺度观测技术空间尺度不匹配的问题(遥感观测、通量观测和模型模拟、大数据整合等), 同时也为充分利用各种高新技术发展, 推动生态系统生态学自身的发展打开了一扇门(图 8)。随着宏观尺度的高新技术快速发展(遥感、雷达和通量观测), 将可能会产生更多或可用于解释生态系统结构、性状和功能的参数, 如叶面积指数、比叶面积、荧光参数、群落结构参数等, 未来必将成为相关领域新的生长点。因此, 生态系统性状为构建地面测试参数与高新技术桥梁奠定了坚实的基础, 将极大地推动生态系统生态学自身的发展;当然, 其发展方向和发展程度很大程度依赖于新技术的发展速度及其与生态系统生态学理论研究的紧密程度。
3.3 推动宏生态研究与应用范畴从技术手段来说, 宏生态学主要依赖于当前高速发展的遥感观测、通量观测、模型模拟和大数据整合的高新技术, 随着这些技术的发展人们可以获得越来越多的参数, 尤其是日益精细的各种光谱参数和高精度雷达透视技术, 将是未来宏生态学发展和应用的利器。然而, 无论这些高新观测技术如何发展, 都需要生态系统的地面实测数据的支撑、验证和检验;先前由于地面性状测试数据与它们在空间尺度上的不匹配, 导致许多遥感产品、通量观测数据和模型模拟结果难以被验证, 难以提升精度。随着生态系统性状概念体系和推导方法的提出与发展, 传统地面测定的大量生态参数将可能会被转化成为单位土地面积上标准化的生态系统性状数据, 用于验证宏生态学的主要技术, 提高它们观测或预测精度, 更好地解决各类生态环境问题(图 9)。

当前, 全球变化的生态效应多是在从态系统水平进行评估的。在利用N:P变化来探讨养分限制性或未来氮磷沉降不对称的生态效应时, 如果能用群落水平的N:P(群落内不同物种长期适应与权衡的整体表现形式)数据, 其评估结果应该可以更准确。此外, 也能为生态模型提供更多和更准确的关键参数, 将显著提高其模拟精度[13, 32]。以C:N为例, 由于群落水平数据难以获得, 大多数生态模型均使用个别物种数据或少数物种算数平均值来替代, 一定程度影响了模型的拟合精度, 但是可以提供更精确的群落水平C:N, 降低关键输入参数的不确定性, 可有效地提高相关模型的预测精度。生态系统性状的提出能促使更多植被性状纳入生态模型, 为开发新一代模型奠定坚实基础。
4 生态系统性状发展与应用面临的主要困难与挑战 4.1 生态系统生态学具有多学科交叉特色, 各学科来源名词能否统一?生态系统生态学是一个特定空间范围内的以生态系统为研究对象的科学, 因此其具有非常明确的学科交叉特性(图 10)。由于各个学科在起源、研究对象等方面存在不同, 使得描述生态系统内相似特征的名词多种多样。例如, 性状来源于遗传学和生理学, 但主要用于植物叶、枝、干、根器官;而其他学科却广泛使用特征、属性、参数等名词来描述生态系统不同组分。这些被广泛使用的名词, 在各自学科上都具有合理性和可溯源性;然而, 在生态系统尺度, 没有相对统一的名词将一定程度阻碍生态系统生态学的发展。理论上, 由于他们的描述目的以及英文起源都非常相近(强调在生态系统研究时的统一, 并不排斥或抹杀各学科自身的特点或名词使用权), 因此, 从生态系统尺度统一这些名词并不困难。总之, 生态系统生态学研究过程, 使用统一名词是可行的、必要的, 是其学科自身发展的迫切需求;鉴于生态系统整体也具有适应性、抗性和可再生能力, 综合上述讨论建议统一采用生态系统性状。

|
| 图 10 生态系统生态学具有交叉学科特点使其关键概念难以统一 Fig. 10 Ecosystem ecology with interdisciplinary feature make it difficult to unify some key concepts |
尺度拓展过程中的非线性问题, 是生态学所有分支学科共同面临的巨大挑战。从生态系统性状的基本定义, 可清楚地看出其具有将传统性状或参数推导到单位土地面积的过程。因此, 生态系统性状概念的进一步发展和应用必然要面临两个巨大的挑战:(1)哪些性状可以推导至生态系统性状, 哪些性状不能?(2)如何建立生态系统性状与功能的定量联系?在复杂的生态系统中, 几乎每个性状都存在冗余、互补或竞争等复杂情况, 使其推导过程、生态效应等均呈现出非线性问题[33]。更为重要的是:在复杂自然生态系统中, 任何适应机制与功能优化机制都是通过多种性状的协同来完成的(甚至拮抗效应), 过分强调单一性状的重要性或绝对贡献可能会得出错误的结论(图 11)。例如近期研究表明:植物群落的叶绿素含量仅能解释其GPP或NPP的30%[30], 气孔导度和特性仅能解释水分利用率(Water Use Efficiency, WUE)利用效率的44%[20]。虽然有如此多的困难与挑战, 但生态系统性状开启了性状“从器官到生态系统、从理论发展到区域应用”的大门。

|
| 图 11 生态系统性状推导、性状与功能关系间的非线性问题将是巨大挑战 Fig. 11 The nonlinear problems in the processes of traits scaling-up require to be resolved and the relationship between trait and functioning at different scales are big challenges |
通过上述大量讨论, 可以清楚地看出生态系统性状, 尤其是动物和植物性状, 需要大量的、系统性的测定数据为支撑;因此, 如何快速拓展可用于推导生态系统性状的数据源是一个重要瓶颈。目前著名的TRY数据库, 因其缺乏群落结构数据且每个地点物种数有限, 难以实现大面积植物群落性状的推导;因此, 建立植物群落性状与物种数量的关系将成为未来研究的重要途径[28]。然而, 需要看到现实调查工作的另一面;在实际调查过程, 许多科学家在特定地点或多个地点, 都开展了系统的群落结构与性状调查, 但受传统概念的影响与个人理解的差异, 这类系统性性状数据或群落数据很少被并公布并积累下来, 造成巨大损失。随着“系统性+符合自然”数据库理论的建立、生态系统性状概念体系的发展与应用, 相信类似的系统性调查数据将会更多、更好地保存下来, 从而形成生态系统性状的重要数据源[22]。此外, 中国和世界各地的野外生态台站长期实验样地、BEF大样地等, 将具有开展系统性性状调查数据的所有必要条件, 将成为这方面的主力军。总之, 随着生态系统性状概念、推导方法和应用的发展, 当拥有多个(几百或上千个)地点详细调查数据的新型性状数据库时, 必将在性状多维度研究、生态系统生态学自身研究、与宏生态相结合的区域应用等方面取得重要突破。除此之外, 生态系统性状概念和理论框架发展下去的生命力在于有一些特征性的和新的指标, 并能发展出新指标的计算方法。随着技术的进步和人们认识的提高, 生态系统性状的内涵会更加丰富, 如生态系统的碳利用效率已经引起了人们的重视, Chen等[34]探究了中国50个生态系统的碳利用效率的分布格局及其影响机制, He等[35]利用生态模型在全球尺度上绘制了碳利用效率的栅格图等。在此呼吁不同学科的科学家不断丰富生态系统性状的内涵, 尤其是从动物群落性状和微生物群落性状角度的深入探讨, 为宏生态学的研究提供坚实的理论和数据基础。
5 结语生态系统性状新概念和理论体系的提出与发展, 是以前期基础理论积累为基础, 是学科自身发展的体现, 且具有明确的时代需求。虽然当前其在方法学和数据源等方面仍不完善, 但它却将传统性状研究拓展到了群落和生态系统水平, 构建了一个将传统性状与宏生态研究(或宏观地学研究)的桥梁, 给人们展现了一个实现“性状研究、生态系统生态学研究、宏观生态研究”多赢的途径和方法。本文是对先前《Trends in Ecology and Evolution》论文深入思考后的解读与补充, 希望能通过广泛的讨论, 尤其是希望能从动物群落性状和微生物群落性状等角度加强交流, 逐渐完善生态系统性状的基本理论、推动“以性状为基础的生态系统生态学”新研究框架的发展, 并切实推动性状研究在区域生态环境问题上的应用。
| [1] |
Tansley AG. The use and abuse of vegetational concepts and terms. Ecology, 1935, 16(3): 284-307. |
| [2] |
Odum HT. Trophic structure and productivity of Silver Springs, Florida. Ecological Monographs, 1957, 27(1): 55-112. DOI:10.2307/1948571 |
| [3] |
Lindeman R L. The trophic-dynamic aspect of ecology. Ecology, 1942, 23(4): 399-417. DOI:10.2307/1930126 |
| [4] |
Huang Y Y, Chen Y X, Castro-Izaguirre N, Baruffol M, Brezzi M, Lang A N, Li Y, Härdtle W, von Oheimb G, Yang X F, Liu X J, Pei K Q, Both S, Yang B, Eichenberg D, Assmann T, Bauhus J, Behrens T, Buscot F, Chen XY, Chesters D, Ding BY, Durka W, Erfmeier A, Fang J Y, Fischer M, Guo LD, Guo D L, Gutknecht JLM, He JS, He CL, Hector A, Hönig L, Hu RY, Klein AM, Kühn P, Liang Y, Li S, Michalski S, Scherer-Lorenzen M, Schmidt K, Scholten T, Schuldt A, Shi X Z, Tan MZ, Tang Z Y, Trogisch S, Wang Z W, Welk E, Wirth C, Wubet T, Xiang W H, Yu M J, Yu XD, Zhang J Y, Zhang S R, Zhang N L, Zhou HZ, Zhu CD, Zhu L, Bruelheide H, Ma K P, Niklaus PA, Schmid B. Impacts of species richness on productivity in a large-scale subtropical forest experiment. Science, 2018, 362(6410): 80-83. DOI:10.1126/science.aat6405 |
| [5] |
Gravel D, Bell T, Barbera C, Bouvier T, Pommier T, Venail P, Mouquet N. Experimental niche evolution alters the strength of the diversity-productivity relationship. Nature, 2011, 469(7328): 89-92. DOI:10.1038/nature09592 |
| [6] |
O'Connor MI, Gonzalez A, Byrnes JEK, Cardinale BJ, Duffy JE, Gamfeldt L, Griffin JN, Hooper D, Hungate BA, Paquette A, Thompson PL, Dee LE, Dolan KL. A general biodiversity-function relationship is mediated by trophic level. Oikos, 2017, 126(1): 18-31. DOI:10.1111/oik.03652 |
| [7] |
Snelgrove PVR, Thrush SF, Wall DH, Norkko A. Real world biodiversity-ecosystem functioning:a seafloor perspective. Trends in Ecology & Evolution, 2014, 29(7): 398-405. |
| [8] |
Díaz S, Lavorel S, de Bello F, Quétier F, Grigulis K, Robson TM. Incorporating plant functional diversity effects in ecosystem service assessments. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(52): 20684-20689. DOI:10.1073/pnas.0704716104 |
| [9] |
Naeem S, Wright JP. Disentangling biodiversity effects on ecosystem functioning:deriving solutions to a seemingly insurmountable problem. Ecology Letters, 2003, 6(6): 567-579. DOI:10.1046/j.1461-0248.2003.00471.x |
| [10] |
Lavorel S, Garnier E. Predicting changes in community composition and ecosystem functioning from plant traits:revisiting the Holy Grail. Functional Ecology, 2002, 16(5): 545-556. DOI:10.1046/j.1365-2435.2002.00664.x |
| [11] |
Cornelissen JHC, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich DE, Reich PB, ter Steege H, Morgan HD, van der Heijden MGA, Pausas JG, Poorter H. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany, 2003, 51(4): 335-380. DOI:10.1071/BT02124 |
| [12] |
Moretti M, Dias ATC, de Bello F, Altermatt F, Chown SL, Azcarate FM, Bell JR, Fournier B, Hedde M, Hortal J, Ibanez S, Öckinger E, Sousa JP, Ellers J, Berg MP. Handbook of protocols for standardized measurement of terrestrial invertebrate functional traits. Functional Ecology, 2017, 31(3): 558-567. DOI:10.1111/1365-2435.12776 |
| [13] |
Lu X J, Wang Y P, Wright I J, Reich P B, Shi Z, Dai Y J. Incorporation of plant traits in a land surface model helps explain the global biogeographical distribution of major forest functional types. Global Ecology and Biogeography, 2017, 26(3): 304-317. DOI:10.1111/geb.12535 |
| [14] |
Kunstler G, Falster D, Coomes DA, Hui F, Kooyman RM, Laughlin DC, Poorter L, Vanderwel M, Vieilledent G, Wright SJ, Aiba M, Baraloto C, Caspersen J, Cornelissen JHC, Gourlet-Fleury S, Hanewinkel M, Herault B, Kattge J, Kurokawa H, Onoda Y, Peñuelas J, Poorter H, Uriarte M, Richardson S, Ruiz-Benito P, Sun IF, Ståhl G, Swenson NG, Thompson J, Westerlund B, Wirth C, Zavala MA, Zeng HC, Zimmerman JK, Zimmermann NE, Westoby M. Plant functional traits have globally consistent effects on competition. Nature, 2016, 529(7585): 204-207. DOI:10.1038/nature16476 |
| [15] |
Violle C, Navas ML, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E. Let the concept of trait be functional!. Oikos, 2007, 116(5): 882-892. DOI:10.1111/j.0030-1299.2007.15559.x |
| [16] |
Wang RL, Yu GR, He NP, Wang QF, Zhao N, Xu ZW. Latitudinal variation of leaf morphological traits from species to communities along a forest transect in eastern China. Journal of Geographical Sciences, 2016, 26(1): 15-26. DOI:10.1007/s11442-016-1251-x |
| [17] |
Zhang JH, Zhao N, Liu CC, Yang H, Li ML, Yu GR, Wilcox K, Yu Q, He NP. C:N:P stoichiometry in China's forests:from organs to ecosystems. Functional Ecology, 2018, 32(1): 50-60. |
| [18] |
Wieczynski DJ, Boyle B, Buzzard V, Duran SM, Henderson AN, Hulshof CM, Kerkhoff AJ, McCarthy MC, Michaletz ST, Swenson NG, Asner GP, Bentley LP, Enquist BJ, Savage VM. Climate shapes and shifts functional biodiversity in forests worldwide. Proceedings of the National Academy of Sciencesof the United States of America, 2019, 116(2): 587-592. DOI:10.1073/pnas.1813723116 |
| [19] |
He NP, Liu CC, Tian M, Li ML, Yang H, Yu GR, Guo DL, Smith MD, Yu Q, Hou JH. Variation in leaf anatomical traits from tropical to cold-temperate forests and linkage to ecosystem functions. Functional Ecology, 2018, 32(1): 10-19. |
| [20] |
Liu CC, He NP, Zhang JH, Li Y, Wang QF, Sack L, Yu GR. Variation of stomatal traits from cold temperate to tropical forests and association with water use efficiency. Functional Ecology, 2018, 32(1): 20-28. |
| [21] |
Wang RL, Yu GR, He NP, Wang QF, Zhao N, Xu ZW, Ge JP. Latitudinal variation of leaf stomatal traits from species to community level in forests:linkage with ecosystem productivity. Scientific Reports, 2015, 5: 14454. DOI:10.1038/srep14454 |
| [22] |
He NP, Liu CC, Piao SL, Sack L, Xu L, Luo YQ, He JS, Han XG, Zhou GS, Zhou XH, Lin Y, Yu Q, Liu SR, Sun W, Niu SL, Li SG, Zhang JH, Yu GR. Ecosystem traits linking functional traits to macroecology. Trends in Ecology & Evolution, 2019, 34(3): 200-210. |
| [23] |
Ma ZQ, Guo DL, Xu XL, Lu MZ, Bardgett RD, Eissenstat DM, McCormack ML, Hedin LO. Evolutionary history resolves global organization of root functional traits. Nature, 2018, 555(7694): 94-97. DOI:10.1038/nature25783 |
| [24] |
Wright IJ, Dong N, Maire V, Prentice IC, Westoby M, Díaz S, Gallagher RV, Jacobs BF, Kooyman R, Law EA, Leishman MR, Niinemets V, Reich PB, Sack L, Villar R, Wang H, Wilf P. Global climatic drivers of leaf size. Science, 2017, 357(6354): 917-921. DOI:10.1126/science.aal4760 |
| [25] |
Borgy B, Violle C, Choler P, Garnier E, Kattge J, Loranger J, Amiaud B, Cellier P, Debarros G, Denelle P, Diquelou S, Gachet S, Jolivet C, Lavorel S, Lemauviel-Lavenant S, Mikolajczak A, Munoz F, Olivier J, Viovy N. Sensitivity of community-level trait-environment relationships to data representativeness:a test for functional biogeography. Global Ecology and Biogeography, 2017, 26(6): 729-739. DOI:10.1111/geb.12573 |
| [26] |
何念鹏, 张佳慧, 刘聪聪, 徐丽, 陈智, 刘远, 王瑞丽, 赵宁, 徐志伟, 田静, 王情, 朱剑兴, 李颖, 侯继华, 于贵瑞. 森林生态系统性状的空间格局与影响因素研究进展——基于中国东部样带的整合分析. 生态学报, 2018, 38(18): 6359-6382. |
| [27] |
Kattge J, Díaz S, Lavorel S, Prentice I C, Leadley P, Bönisch G, Garnier E, Westoby M, Reich PB, Wright IJ, Cornelissen JHC, Violle C, Harrison SP, van Bodegom PM, Reichstein M, Enquist BJ, Soudzilovskaia NA, Ackerly DD, Anand M, Atkin O, Bahn M, Baker TR, Baldocchi D, Bekker R, Blanco CC, Blonder B, Bond WJ, Bradstock R, Bunker DE, Casanoves F, Cavender-Bares J, Chambers JQ, Chapin III FS, Chave J, Coomes D, Cornwell WK, Craine JM, Dobrin BH, Duarte L, Durka W, Elser J, Esser G, Estiarte M, Fagan WF, Fang J, Fernández-Méndez F, Fidelis A, Finegan B, Flores O, Ford H, Frank D, Freschet GT, Fyllas NM, Gallagher RV, Green WA, Gutierrez AG, Hickler T, Higgins SI, Hodgson JG, Jalili A, Jansen S, Joly CA, Kerkhoff AJ, Kirkup D, Kitajima K, Kleyer M, Klotz S, Knops JMH, Kramer K, Kühn I, Kurokawa H, Laughlin D, Lee TD, Leishman M, Lens F, Lenz T, Lewis SL, Lloyd J, Llusià J, Louault F, Ma S, Mahecha MD, Manning P, Massad T, Medlyn BE, Messier J, Moles AT, Müller SC, Nadrowski K, Naeem S, Niinemets V, Nöllert S, Nüske A, Ogaya R, Oleksyn J, Onipchenko VG, Onoda Y, Ordoñez J, Overbeck G, Ozinga WA, Patiño S, Paula S, Pausas JG, Peñuelas J, Phillips OL, Pillar V, Poorter H, Poorter L, Poschlod P, Prinzing A, Proulx R, Rammig A, Reinsch S, Reu B, Sack L, Salgado-Negre B, Sardans J, Shiodera S, Shipley B, Siefert A, Sosinski E, Soussana JF, Swaine E, Swenson N, Thompson K, Thornton P, Waldram M, Weiher E, White M, White S, Wright SJ, Yguel B, Zaehle S, Zanne AE, Wirth C. TRY-a global database of plant traits. Global Change Biology, 2011, 17(9): 2905-2935. DOI:10.1111/j.1365-2486.2011.02451.x |
| [28] |
何念鹏, 刘聪聪, 张佳慧, 徐丽, 于贵瑞. 植物性状研究的机遇与挑战:从器官到群落. 生态学报, 2018, 38(19): 6787-6796. |
| [29] |
Li N N, He NP, Yu GR, Wang QF, Sun J. Leaf non-structural carbohydrates regulated by plant functional groups and climate:evidences from a tropical to cold-temperate forest transect. Ecological Indicators, 2016, 62: 22-31. DOI:10.1016/j.ecolind.2015.11.017 |
| [30] |
Li Y, Liu CC, Zhang JH, Yang H, Xu L, Wang QF, Sack L, Wu XQ, Hou JH, He NP. Variation in leaf chlorophyll concentration from tropical to cold-temperate forests:association with gross primary productivity. Ecological Indicators, 2018, 85: 383-389. DOI:10.1016/j.ecolind.2017.10.025 |
| [31] |
Reichstein M, Bahn M, Mahecha MD, Kattge J, Baldocchi DD. Linking plant and ecosystem functional biogeography. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(38): 13697-13702. DOI:10.1073/pnas.1216065111 |
| [32] |
Connolly J, Bell T, Bolger T, Brophy C, Carnus T, Finn JA, Kirwan L, Isbell F, Levine J, Lüscher A, Picasso V, Roscher C, Sebastia MT, Suter M, Weigelt A. An improved model to predict the effects of changing biodiversity levels on ecosystem function. Journal of Ecology, 2013, 101(2): 344-355. DOI:10.1111/1365-2745.12052 |
| [33] |
Croft H, Chen JM, Luo XZ, Bartlett P, Chen B, Staebler RM. Leaf chlorophyll content as a proxy for leaf photosynthetic capacity. Global Change Biology, 2017, 23(9): 3513-3524. DOI:10.1111/gcb.13599 |
| [34] |
Chen Z, Yu G R, Wang Q F. Ecosystem carbon use efficiency in China:variation and influence factors. Ecological Indicators, 2018, 90: 316-323. DOI:10.1016/j.ecolind.2018.03.025 |
| [35] |
He Y, Piao S L, Li X Y, Chen A P, Qin D H. Global patterns of vegetation carbon use efficiency and their climate drivers deduced from MODIS satellite data and process-based models. Agricultural and Forest Meteorology, 2018, 256-257: 150-158. DOI:10.1016/j.agrformet.2018.03.009 |