 2020, Vol. 40
2020, Vol. 40文章信息
- 刘金亮, 薛滨, 姚书春, 程龙娟, 李玲玲
- LIU Jinliang, XUE Bin, YAO Shuchun, CHENG Longjuan, LI Lingling
- 湖泊水生植物稳定碳同位素分馏机制与应用研究进展
- Mechanisms of stable carbon isotope fractionation of aquatic plants and the research advances of application
- 生态学报. 2020, 40(8): 2533-2544
- Acta Ecologica Sinica. 2020, 40(8): 2533-2544
- http://dx.doi.org/10.5846/stxb201903060419
-
文章历史
- 收稿日期: 2019-03-06
- 网络出版日期: 2019-12-31





2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
水生植物是湖泊生态系统的重要组成部分, 是维持湖泊生态系统多样性和稳定性的重要基础与支撑[1]。碳是植物生长不可或缺的重要元素之一, 水生植物体内稳定碳同位素组成不仅记录了植物生长过程中外界环境信息的变化, 还可以揭示碳的生物地球化学循环过程[2]。水生植物稳定碳同位素已被广泛应用于研究水生生态系统演变和营养传递规律等方面, 逐步成为研究水生植物与环境间相互关系的重要方法[2-3]。
与陆地植物相比, 水生植物由于生活在特定的水体环境中, 其碳在合成过程中的分馏机制较复杂, 影响因素众多, 系统的了解水生植物稳定碳同位素的分馏模式和影响其组成变化的主要因素, 对于研究湖泊食物网结构和生态系统演变具有重要意义。本文详细的介绍了水生植物稳定碳同位素的分馏过程和影响其碳同位素分馏的主要因素, 阐述了水生植物稳定碳同位素在水域生态学和湖泊古环境研究中的应用, 探讨了研究中存在的问题, 并以太湖典型水生植物开展实例研究, 对本领域未来的研究趋势进行了展望。
1 水生植物稳定碳同位素分馏 1.1 水生植物稳定碳同位素的分馏机制在光合作用过程中, 植物通过吸收CO2与H2O合成有机质, Friend等[4]研究发现在这个过程中植物体内的碳同位素发生了不同程度的分馏:首先, 大气中的CO2在向植物气孔扩散过程中会发生同位素分馏, 导致植物体内CO2的δ13C较大气中CO2的δ13C低;其次, CO2从细胞间隙进入叶肉细胞时会遇到一定的阻力, 由于动力学分馏效应的影响, 质量较轻的12CO2由于扩散速率快而更易被植物吸收。最后, 植物体内的CO2在羧基多肽酶的作用下发生羧化反应, 溶解在细胞内的12CO2与核酮糖1, 5-二磷酸核酮糖羧化酶/加氧酶(Rubisco)通过酶的作用被优先结合到磷酸甘油酸中(PGA), 而剩余的CO2富集重13CO2。水生植物主要包括水生维管束植物和高等藻类(以浮游植物为主), 而水生维管束植物又可依据它们的生境进一步划分为沉水型、挺水型、漂浮型和浮叶型植物[5]。其中水生维管束植物的碳同位素分馏方式与陆地C3、C4植物相似[6], 其具体可表示为:
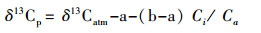
|
(1) |

|
(2) |
式中, δ13Cp与δ13Catm分别代表植物和大气中CO2的δ13C值;a和b分别为大气中的CO2经扩散作用进入植物体内细胞时所产生的分馏(约为4.4‰)和植物体内的CO2经核酮糖二磷酸羧化酶羧化时产生的分馏系数(约为27‰);Ci/Ca分别为植物光合作用过程中, 叶片细胞内外的CO2浓度;b4为磷酸烯醇式丙酮酸羧化酶(PEP)羧化过程中的同位素分馏(约为-5.9‰);φ为C4脱羧释放CO2时从PCK循环泄露到PCA循环的比例;ai是泄露过程中发生的碳同位素分馏(约为1.8‰)。
而浮游植物的碳同位素分馏可表示为[7]:
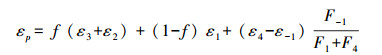
|
(3) |
其中, f为浮游植物细胞内HCO3-所占吸收总碳的比重;F1和F-1为CO2进出细胞的通量, F4为HCO3-进入细胞的通量;ε-1为HCO3-进入植物体内细胞时产生的分馏, ε1与ε2分别为CO2进出植物体内细胞时产生的分馏, ε3为CO2与HCO3-平衡时产生的分馏, ε4为植物固碳过程中Rubisco羧化酶羧化产生的分馏, 约为-31‰—-22‰[8]。
由公式(1)和(2)可以看出, 水生维管束植物的δ13C组成与它们光合作用过程中叶片内外的CO2浓度密切相关(Ci/Ca), 而Ci/Ca则主要受植物的光合作用效率和植物的气孔导度所控制, 植物光合作用效率和气孔导度除受自身生理特性影响之外, 还主要受植物生长过程中外界环境因子的变化(如温度、水深等)所影响。公式(3)中浮游植物的δ13C组成主要受它们吸收CO2与HCO3-所占的比重和通量所影响。由以上公式可见, 水生植物在光合作用过程中碳同位素产生分馏主要发生在植物吸收CO2、CO2扩散和固碳羧化酶羧化三个过程中(图 1), 因此水生植物的δ13C变化主要与其碳吸收同化机制和植物气孔导度有关[9]。

早在20世纪70年代, 就有学者发现陆生植物的δ13C可以用来区分不同植物的光合作用类型[11]。根据植物不同光合作用的途径可以将陆地植物划分为C3、C4和CAM植物, 不同类型的植物具有不同的δ13C。C3植物的δ13C相对偏轻, 主要分布在-33‰—-21‰之间, 光合作用碳同化的途径主要通过卡尔文循环, 大气中CO2被细胞吸收后经扩散发生动力分馏, 之后被1, 5-二磷酸核酮糖(RuBP)固定再次发生动力分馏(图 2)。C4植物的δ13C值相对偏重, 主要分布在-16‰—-9‰之间。C4植物能够在干旱环境下关闭气孔防止水分流失, 利用细胞间低浓度的CO2进行光合作用维持其生长(图 2)。CAM植物分布较少, 其δ13C值介于-20‰—-10‰之间, 夜间植物固定CO2比例和环境中水分条件是影响CAM植物δ13C变化的主要因素[12]。

与陆地植物相比, 水生植物的δ13C分布范围较离散(图 3), 介于-50‰—-11‰之间[14]。在水生植物中同样分布着C3、C4和CAM植物, 但大部分水生植物属于C3型[15]。挺水植物可以直接利用大气中的CO2进行光合作用, 其δ13C分布在-30‰—-24‰之间, 与C3植物的δ13C变化范围相重合, 相似光合作用途径的水生植物种类还包括浮叶植物和漂浮植物[15]。相反, 沉水植物由于水中碳供给不足, 可以利用溶解的HCO3-作为碳源, 导致其δ13C值较C3植物偏重, 并且与C4植物的δ13C变化范围有所重叠。如Chappuis等[5]研究发现伊乐藻属(Elodea canadensis)和眼子菜属(Potamogeton alpinus)等沉水植物的δ13C偏重, 属于C4光合途径。浮游植物可以利用水中溶解的CO2和部分HCO3-作为碳源, 当利用水中溶解的CO2进行光合作用时, 其δ13C组成偏轻, 与C3植物相类似;当浮游植物利用HCO3-进行光合作用时, 其δ13C值偏重, 与部分C4植物δ13C的变化范围相重叠[3]。此外, 也有部分水生植物的δ13C介于沉水植物与浮游植物的δ13C之间, 与CAM植物的δ13C变化范围相似, 如挺水植物中的华水韭(Isoetes howellii)就是CAM光合途径[16]。沉水CAM植物发育了良好的表面扩散边界层, 具有CAM暗反应来固定无机碳, 能增加对CO2的亲和力以及可以利用HCO3-作为碳源[17]。

水生植物中的碳主要源于大气、水体和沉积物的孔隙水中, 来源较为广泛[18]。不同碳源具有不同的碳同位素信号, 并且不同类型的水生植物利用无机碳的形式也不一样, 复杂的水体环境使得影响水生植物碳同位素化学分馏的因素众多。
2.1 碳源水中溶解无机碳(DIC)的形式主要有三种:溶解水中的二氧化碳[(CO2)aq]、离子态的碳酸氢盐(HCO3-)和碳酸盐(CO32-)。在大多数的水体(pH>7)中, 溶解于水中的CO2在DIC三种形式中所占的比例较低, HCO3-比例较大, 并且当pH升高时, HCO3-/CO2的比重急剧上升[19]。Madsen[20]研究发现CO2在水体中的扩散速率较慢, 浓度较空气中偏低(在15℃为6 μmol/L), 而且由于植物细胞外扩散边界层较厚, 导致沉水植物可吸收利用的CO2不足。因此, 沉水植物为适应水体环境中碳浓度不足的条件, 形成了可以利用无机碳的生理结构和吸收HCO3-的酶系统, 并且可通过碳酸酐酶来催化HCO3-与CO2之间的转化, 维持较高的光合作用速率[21]。由于HCO3-比溶解态的CO2更富集δ13C, 偏重约7‰—11‰, 导致沉水植物的δ13C值偏重[22]。大气中CO2的δ13C值在-7.8‰左右, 且变化较小, 而挺水植物、浮叶植物和漂浮植物的叶片暴露在大气中, 主要受大气中CO2的δ13C影响, 使得这三种类型植物的δ13C变化相对较小。而浮游植物进行光合作用可以利用水中溶解的CO2和部分HCO3-作为碳源, 导致浮游植物的δ13C变化范围相对较大。此外, 水体环境中CO2在水-气的扩散过程和碳酸盐的化学风化与矿化过程等会改变水中DIC的δ13C, 从而进一步影响水生植物的δ13C[23]。因此, 不同生活型的水生植物受不同碳源的δ13C影响, 导致不同水生植物的δ13C存在一定的差异[24]。
2.2 水体DIC浓度沉水植物在光合作用过程中, 溶解于水中的CO2是最容易利用的碳源, 但是由于大多数水体(pH>7)中无机碳离子形态主要是HCO3-, 因此沉水植物可能会更多的吸收HCO3-, 但是沉水植物通过光照后H-ATP酶活性提高, 使得表皮中的pH降低, 最终导致HCO3-/CO2的比率降低, 从而可以吸收利用更多的CO2[25]。沉水植物碳同位素和溶解无机碳浓度的关系可表示为[6]:
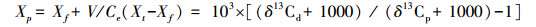
|
(4) |
其中, Xt和Xf为细胞吸收环境中CO2和固碳所产生的同位素效应;V是植物体细胞内外CO2的浓度梯度;Ce为水中溶解的CO2浓度;δ13Cd和δ13Cp分别是溶解的无机碳和沉水植物的碳同位素。由上式可以看出水中溶解无机碳的浓度越低, 沉水植物的δ13C越偏重。林清[26]和刘勇丽[27]通过室内控制实验和野外实验也证实了沉水植物的δ13C与水中DIC的浓度具有负相关的关系, 由此可见水体中DIC的浓度是影响沉水植物δ13C变化的重要因素。
2.3 水体pHHinga等[28]研究表明水体的pH不仅可以直接影响水生植物的碳同位素分馏, 也可以通过改变碳酸盐的化学性质间接的影响水生植物碳同位素的平衡分馏。首先, 水体的pH升高会导致CO2中富集12C, 水生植物受同位素平衡分馏影响而δ13C偏负。其次, 水体的pH会影响浮游植物光合作用过程中无机碳的传输、转化(碳浓缩机制)和碳酸酐酶的活性, 从而改变浮游植物的δ13C[29-30]。此外, 水体的pH可以控制水生植物细胞内CO2的渗漏。Sharkey等[31]提出碳源与细胞内CO2的外逸是影响植物碳同位素变化的重要因素, pH增加通过碳浓缩机制引起细胞内CO2的渗漏减少, 导致碳同位素分馏减少。最后, 外界环境中pH也会影响水生植物细胞内的pH, 进而可以通过调控细胞内的[CO2(aq)]和CO2的渗漏影响水生植物的碳同位素分馏[32]。
此外, 水体的pH还决定了水体中不同形式无机碳组成所占的比重, 当水体的pH处于5时, 此时水体中溶解的无机碳形式主要为CO2;当pH在7.5—8之间时, 水体中溶解的无机碳形式以HCO3-为主, 但是仍有部分可供吸收的CO2[33]。水体中pH的变化决定了水体中溶解无机碳的形式, 因此水体的pH也可以间接的影响水生植物的δ13C值。如Maberly[34]和Xu等[35]研究证实水体的pH升高, 导致水体中溶解的DIC不足, 使得水中的碳低于水生植物的需求, 从而抑制了碳同位素的分馏。
2.4 其他因素① 温度。温度不仅可以通过改变水生植物光合作用过程中的反应速率影响碳同位素分馏, 也可以通过调节水生植物体内的生物化学过程和生理变化影响碳同位素分馏[36]。如林清[26]通过室内控制实验, 发现温度升高, 龙须眼子菜的碳同位素分馏减少, 二者之间具有明显的相关性。②水体富营养化。水体富营养化会导致水体中DIC消耗增加, 进而改变水体的pH值, 从而可以间接的影响水生植物的δ13C[37]。张金美等[38]通过探讨水华条件下鄱阳湖水生植物碳同位素的变化特性, 证实了马来眼子菜的δ13C受水华条件影响显著, 可以被用于指示水体环境灾变。③水位。湖泊不同水深处的水生植物的生境存在差异, 不同水深的碳源也分布不同, 从而也会影响水生植物的碳同位素组成[39-40]。
不同生活型的水生植物吸收利用不同形式的碳源, 使得不同水生植物的δ13C存在显著差异, 而水体环境要素(pH、DIC浓度、温度和营养盐浓度等)可以直接通过影响水生植物的碳同位素分馏过程或者间接的影响其吸收利用碳源的δ13C, 从而改变水生植物的δ13C。此外, 水生植物稳定碳同位素的分馏过程除受外界环境因素影响之外, 还受自身遗传特性和生理学特征所控制。目前对水生植物自身生理特性的变化与环境因子之间的对应关系研究相对较少, 要全面理解水生植物稳定碳同位素对环境变化的响应机理, 需要加强植物生理学、生态学和同位素地球化学等多学科之间的交叉研究。
3 水生植物稳定碳同位素的应用 3.1 在生态学中的应用 3.1.1 在湖泊食物网中的应用水生植物可为浮游动物和鱼类等水生生物等提供食物来源, 是驱动水生食物网能量流动与传递的重要基础, 而稳定碳同位素技术可以用来指示食物的来源, 区分不同初级生产者对消费者碳源的贡献[41]。生态系统中不同初级生产者的δ13C存在明显的差异, 而这种差异可用于识别消费者的食物来源和追踪生态系统中的碳流动。同时, 根据不同水生生物δ13C的富集特征可以确定它们在食物网中的营养级[42]。在研究湖泊食物网结构与营养级关系时, 利用一种同位素往往存在不足, 而且不同水生植物的δ13C存在时空上的差异, 植物不同组织的δ13C也不同[43]。因此基于具体的研究目标, 选择合适的多元素同位素分析方法与样品的处理可能是解决湖泊生态系统中物质传输和能量流动等科学问题的关键。
3.1.2 在碳循环研究中的应用水生植物是驱动水体环境碳循环的关键因素, 浮游植物通过光合作用可以促进碳在POC和DIC等不同形态之间的转换, 从而利用浮游植物的δ13C可以定量计算水体中POC内外源的贡献份额[44]。岩溶地区的水生植物拥有巨大的固碳量, 是稳定的生物碳汇。岩溶区淡水的pH多高于7, 溶解的无机碳形式主要以HCO3-为主。水生植物可以利用碳酸酐酶吸收HCO3-进行光合作用, 因此根据水生植物的δ13C可推断出水生植物光合作用固定HCO3-的比例, 进而研究水生植物代谢活动对岩溶碳汇的贡献与影响。如章程等[45]和李瑞等[46]通过测试岩溶湿地水生植物的δ13C, 估算了研究区不同水生植物利用光合作用固定HCO3-的比例, 发现水生植物的固碳效果十分显著, 水生植物光合作用与岩溶作用结合显示出巨大的碳汇效应。水生植物光合作用对湖泊初级生产力和碳元素的地球化学循环具有举足轻重的作用, 理清水生植物对湖泊生态系统碳库的贡献量, 对于研究区域和全球碳循环具有重要意义。
3.1.3 在湖泊营养化研究中的应用不同环境条件下的水生植物具有不同的δ13C, 当湖泊的营养水平发生变化时, 藻类与高等水生植物产生竞争, 导致水生植物的δ13C发生改变。如Wang等[47]研究发现湖泊水生植物的δ13C随水体营养水平升高而降低, 二者之间具有显著的负相关关系, 挺水植物的δ13C可作为湖泊营养水平变化的代用指标。而张金美等[38]探讨了水华条件下马来眼子菜(Potamogeton malaianus)的δ13C变化特征, 结果显示马来眼子菜对环境变化的响应较敏感, 能够很好的指示环境灾变。水生植物是联系微观生态学和宏观生态学的重要桥梁, 而碳同位素具有分布广泛、测量较为成熟等优点, 深入开展水生植物碳同位素在生态学中的研究, 对于我们研究湖泊生态系统的演化与保护具有重要的现实意义。
3.2 在古环境研究中的应用湖泊沉积物是环境气候变化的敏感指示器, 沉积物中的有机质碳同位素变化可以反演古气候的变化历史、判断沉积物中有机质的来源和计算湖泊生产力的变化, 是良好的古气候信息载体之一[48]。水生植物残体作为沉积物中有机质的重要组成成分, 其碳同位素信号包含了丰富的古环境信息。Edwards等[49]首次利用湖泊沉积物中纤维素的δ13C和δ18O重建了加拿大Shield Lake过去一万年以来温度和湿度的变化历史, 变化趋势可分为“冷干-暖干-暖湿-冷湿”四个阶段, 揭示了湖泊沉积物纤维素同位素在古气候变化研究中的巨大潜力。此后, 我国陈毅风等[50]利用泸沽湖沉积物中纤维素的δ13C反演了研究区一百年以来的气候变化。此外, 湖泊沉积物中不同来源的有机质具有不同的δ13C, 利用水生植物残体的δ13C同时结合其他生物学指标可以判断湖泊有机质的来源。如Street-Perrott等[51]测试了爱沙尼亚Äntu Sinijärv Lake水生植物的δ13C和生物标志物等指标, 确定了该湖沉积物中的有机质来源主要为内生, 全新世以来湖泊有机质的输入主要可划分为三类:微藻类(11.4—10.4 ka BP)、轮藻类(10.4—3.5 ka BP)和苔藓类(3.5—0.23 ka BP)。此外, 不同类型水生植物的光合作用途径不同, 其13C富集也不同, 进而可以利用沉积物纤维素的δ13C判定历史时期该地区的植被类型和古水文特征, 从而反演古湖泊的生产力状况[52-53]。总体来看, 目前利用湖泊沉积物纤维素碳同位素在古环境研究中相对较少, 其主要原因是由于湖泊沉积物中植物纤维素含量较低, 提取过程较为复杂, 样品量难以满足测量要求。
虽然水生植物的δ13C已被广泛应用于研究湖泊现代生态系统变化和古环境演化中, 取得了显著的研究成果, 但是由于水生植物中包含了许多次级组分, 如脂类、果胶、木质素和综纤维素(纤维素与半纤维素的总称)等, 不同组分由于化学合成途径不同致其δ13C存在一定的差异, 这对于应用水生植物碳同位素研究湖泊环境变化的结果解译会存在一定的影响, 而这往往容易被忽视[54]。由上述可见, 学者们在利用水生植物δ13C研究湖泊现代生态环境变化时多选用水生植物的全样作为研究载体, 而在研究湖泊古环境演化时多选用水生植物的纤维素组分作为研究载体, 对于研究不同时间尺度上湖泊环境变化的载体存在差异。此外, 利用湖泊沉积物中纤维素组分的碳同位素研究环境变化最早是参考树轮的研究方法, 而在树轮研究中不同组分(全样、综纤维素与α纤维素)的碳同位素对环境变化的响应存在差异, 而这种现象是否存在于湖泊水生植物中尚未知晓[55]。因此, 湖泊水生植物不同组分的碳同位素对环境变化的响应是否存在差异, 水生植物的何种组分更适用于作为研究环境变化的载体需要开展进一步相应的研究, 这对于利用水生植物碳同位素更加准确的研究湖泊环境变化具有重要的现实意义。
4 研究实例:太湖目前, 国内关于水生植物稳定碳同位素的研究较国外相对较少, 而且围绕水生植物不同组分的碳同位素信号特征的区域调查还未见报道。基于此, 以太湖流域水生植物为研究对象, 分析水生植物不同组分δ13C的组成特征及其对环境因素的响应差异, 旨在研究在湖泊环境中水生植物的何种组分(全样、综纤维素与α纤维素)更适用于研究环境变化, 为利用水生植物δ13C更准确的研究环境变化提供数据支持和实验支撑。
太湖(30°55′—32°32′N, 119°52′—121°36′E)是中国第三大淡水湖, 湖泊总面积约2338 km2, 平均水深1.9 m(图 4)。于2018年4月和7月在太湖入湖河流处(R)和湖区(TH)共14个样品点采集了大型水生植物和水样, 本文主要选择研究区盖度较高的挺水植物菰(Zizania latifolia)、沉水植物狐尾藻(Myriophyllum verticillatum)和马来眼子菜(Potamogeton wrightii Morong)以及浮叶植物菱角(Trapa bispinosa)为研究对象, 共采集到44个植物样品和28个水样, 样品低温保存带回实验室以备分析。

|
| 图 4 采样点位置示意图 Fig. 4 Location of the sampling points R代表入湖河流处的采样点, TH代表太湖湖区的采样点 |
在实验室将采集的植物按照“碱-亚氯酸钠法”标准程序提取综纤维素(HC)和α纤维素(AC)[56]。植物样品的δ13C(共132个样品)在中国科学院南京地理与湖泊研究所测定, 测定仪器为MAT253, 测试精度为±0.15‰。在现场采样过程中利用便携式YSI仪器测定水体pH、溶解氧(DO)和水温等理化指标。将采集的水样冷藏带回实验室进行总氮、总磷和溶解无机碳浓度等指标分析, 实验测定方法均参照《湖泊富营养化调查规范》中的方法[57]。
太湖地区菰的δ13C变化范围在-28.25‰ ± 0.93‰(平均值±标准差)之间, 狐尾藻的δ13C变化范围在-12.64‰ ± 2.14‰之间, 马来眼子菜的δ13C变化范围在-15.24‰ ± 2.25‰之间, 菱角的δ13C变化范围在-27.15‰ ± 1.34‰之间, 其中, 沉水植物的δ13C空间变异最大(图 5)。由图 5可见, 沉水植物的δ13C较挺水植物和浮叶植物的δ13C偏重, 并且这三种类型植物全样的δ13C(δ13CW)较纤维素组分的δ13C偏轻, 综纤维素的δ13C(δ13CHC)与α纤维素的δ13C(δ13CAC)较为接近。菰和菱角的δ13C分布与陆地C3植物的变化范围所重叠, 而狐尾藻和马来眼子菜的δ13C分布与陆地C4植物的δ13C变化范围所重合。

|
| 图 5 水生植物各组分的δ13C分布特征 Fig. 5 The distribution characteristics of δ13C of the different components in macrophytes |
由于水生植物不同组分的δ13C对环境变化响应敏感性的研究主要关注的是各组分δ13C的变化趋势与环境要素变化趋势的差异, 因此各组分δ13C变化趋势的差异大小是判断哪种组分更适用于研究环境变化的载体[55]。利用SPSS 23.0对四种植物不同组分的δ13C进行单因素方差分析(one-way ANOVA), 结果显示菰和狐尾藻三种组分之间的δ13C存在显著的差异(P < 0.01), 而马来眼子菜和菱角各组分之间的δ13C差异并不显著, 我们猜测这可能与这两种水生植物所采集到的样品量较少有关。尽管如此, 在利用水生植物稳定碳同位素研究环境变化时, 应谨慎考虑所选用的研究载体。
为了进一步探讨水生植物不同组分的δ13C对环境因子的响应差异, 利用统计软件(Origin 8.0和SPSS 23.0)分析了水生植物不同组分的δ13C与水体理化特征的关系。结果显示pH是影响水生植物δ13C变化的主要因素, 其中菰、狐尾藻和马来眼子菜的δ13C与pH具有正相关关系, 而菱角的δ13C与pH呈负相关(图 6)。由图 6可见, 这四种水生植物α纤维素组分的δ13C对环境的响应较全样和综纤维素组分更加显著, 总体来看水生植物α纤维素组分更适用于作为研究水生态环境变化的研究载体。

|
| 图 6 水生植物各组分的δ13C与pH的相关性 Fig. 6 Correlation of δ13C of macrophytes with pH |
研究结果显示水生植物α纤维素组分的δ13C对水体环境的响应较全样与综纤维素更为明显(图 6), 推测这主要是由于植物中的木质素和抽提物等次级组分对全样的δ13C信号具有一定的干扰。水生植物组成较复杂, 包含了综纤维素、木质素、脂类和果胶等不同组分, 这些不同的组分因化学途径和合成过程存在差异而具有不同的δ13C[58]。在植物全样中, 木质素所占比重较大, 而且木质素与综纤维素合成的时间也存在差异, 木质素可以利用不同时段的光合产物而合成自身, 因此记录了不同时段的δ13C信号, 从而会导致全样的δ13C因木质素干扰对环境响应较综纤维素组分较差[59]。此外, 全样中还包含了许多抽提物, 如淀粉、单糖和油脂等。抽提物在全样中所占比重的变化和迁移也会导致全样和综纤维素的δ13C存在差异, 许多关于树轮和陆地植物的研究也建议去除全样中的抽提物来研究环境变化[60]。纤维素是由葡萄糖以β-1, 4糖苷键组成的链状高分子化合物, 根据手型结构不同可分为α、β、γ纤维素, 其中α纤维素占据很大部分, β和γ纤维素很少。与综纤维素相比, α纤维素组成成分单一、理化性质相对稳定, 有固定的分子式, 对环境变化响应更为敏感[61]。因此, 在利用水生植物δ13C研究环境变化时选用水生植物的α纤维素组分作为研究载体可能会更准确的反映环境变化的信息, 这为利用水生植物稳定碳同位素更准确的研究环境变化提供了实验支持和数据支撑, 对进一步挖掘水生植物碳同位素的环境信息具有重要意义。
5 小结与展望水生植物是湖泊生态系统的重要组成部分, 影响其稳定碳同位素分馏的因素较复杂, 一定程度上限制了其应用, 在今后的研究中应首先加强对水生植物自身生理学特性的研究。植物为适应环境变化可能会使叶片厚度和气孔密度等发生改变, 从而会影响植物的气孔导度和CO2的扩散速率等生理活动, 导致δ13C发生变化[62]。因此, 促进植物生理学、生态学和同位素地球化学的交叉研究, 将有利于更加全面的认识和理解水生植物δ13C变化的机理。其次, 拓宽水生植物α纤维素δ13C在湖泊环境中的应用。目前, 许多学者为拓宽湖泊沉积物纤维素稳定同位素在古环境中的应用空间, 已建立了有效、简便的纤维素提取方法[63]。随着纤维素提取方法的日渐成熟, 而水生植物α纤维素是记录环境变化的良好载体, 有必要开展深入的研究。同时结合其他多种同位素分析方法, 应用统计模型区分不同因子对水生植物δ13C的影响。将水生植物稳定碳同位素分析结合非线性响应函数和遥感技术等, 从中提取更多的环境信息, 加强与其他环境指标的对比, 推动水生植物稳定碳同位素更深层次的发展。
| [1] |
朱金格, 胡维平, 刘鑫, 邓建才. 湖泊水动力对水生植物分布的影响. 生态学报, 2019, 39(2): 454-459. |
| [2] |
Van Hardenbroek M, Chakraborty A, Davies K L, Harding P, Heiri O, Henderson A C G, Holmes J A, Lasher G E, Leng M J, Panizzo V N, Roberts L, Schilder J, Trueman C N, Wooller M J. The stable isotope composition of organic and inorganic fossils in lake sediment records:Current understanding, challenges, and future directions. Quaternary Science Reviews, 2018, 196: 154-176. DOI:10.1016/j.quascirev.2018.08.003 |
| [3] |
韩琼, 王宝利. 浮游植物稳定碳同位素分馏机制及环境应用. 生态学杂志, 2017, 36(5): 1436-1443. |
| [4] |
Friend A D, Woodward F I. Evolutionary and ecophysiological responses of mountain plants to the growing season environment. Advances in Ecological Research, 1990, 20: 59-124. DOI:10.1016/S0065-2504(08)60053-7 |
| [5] |
Chappuis E, Seriñá V, Martí E, Ballesteros E, Gacia E. Decrypting stable isotope (δ13C and δ15N) variability in aquatic plants. Freshwater Biology, 2017, 62(11): 1807-1818. |
| [6] |
Farquhar G D, O'Leary M H, Berry J A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Australian Journal of Plant Physiology, 1982, 9(2): 121-137. |
| [7] |
Burkhardt S, Riebesell U, Zondervan I. Effects of growth rate, CO2 concentration, and cell size on the stable carbon isotope fractionation in marine phytoplankton. Geochimica et Cosmochimica Acta, 1999, 63(22): 3729-3741. DOI:10.1016/S0016-7037(99)00217-3 |
| [8] |
Tcherkez G G B, Farquhar G D, Andrews T J. Despite slow catalysis and confused substrate specificity, all ribulose bisphosphate carboxylases may be nearly perfectly optimized. Proceedings of the National Academy of Sciences of the United States of America, 2006, 103(19): 7246-7251. DOI:10.1073/pnas.0600605103 |
| [9] |
Diefendorf A F, Mueller K E, Wing S L, Koch P L, Freeman K H. Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(13): 5738-5743. DOI:10.1073/pnas.0910513107 |
| [10] |
易现峰. 稳定同位素生态学. 北京: 中国农业出版社, 2007: 46-46.
|
| [11] |
Bender M M, Rouhani I, Vines H M, Black C C. 13C/12C ratio changes in crassulacean Acid metabolism plants. Plant Physiology, 1973, 52(5): 427-430. |
| [12] |
Osmond C B, Allaway W G, Sutton B G, Troughton J H, Queiroz O, Lüttge U, Winter K. Carbon isotope discrimination in photosynthesis of CAM plants. Nature, 1973, 246(5427): 41-42. DOI:10.1038/246041a0 |
| [13] |
林光辉. 稳定同位素生态学. 北京:高等教育出版社, 2013, 80-81. |
| [14] |
Keeley J E, Sandquist D R. Carbon:Freshwater plants. Plant, Cell & Environment, 1992, 15(9): 1021-1035. |
| [15] |
Keeley J E. CAM photosynthesis in submerged aquatic plants. The Botanical Review, 1998, 64(2): 121-175. DOI:10.1007/BF02856581 |
| [16] |
Keeley J E, Bowes G. Gas exchange characteristics of the submerged aquatic crassulacean acid metabolism plant, Isoetes howellii. Plant Physiology, 1982, 70(5): 1455-1458. DOI:10.1104/pp.70.5.1455 |
| [17] |
Madsen T V, Sand-Jensen K. Photosynthetic carbon assimilation in aquatic macrophytes. Aquatic Botany, 1991, 41(1/3): 5-40. |
| [18] |
Milligan H E, Pretzlaw T D, Humphries M M. Stable isotope differentiation of freshwater and terrestrial vascular plants in two subarctic regions. Écoscience, 2010, 17(3): 265-275. DOI:10.2980/17-3-3282 |
| [19] |
Madsen T V. Growth and photosynthetic acclimation by Ranunculus aquatilis L. in response to inorganic carbon availability. New Phytologist, 1993, 125(4): 707-715. DOI:10.1111/j.1469-8137.1993.tb03919.x |
| [20] |
Madsen T V. Resistance to CO2 fixation in the submerged aquatic macrophyte Callitriche stagnalis scop. Journal of Experimental Botany, 1984, 35(3): 338-347. DOI:10.1093/jxb/35.3.338 |
| [21] |
林清, 王绍令. 沉水植物稳定碳同位素组成及影响因素分析. 生态学报, 2001, 21(5): 806-809. DOI:10.3321/j.issn:1000-0933.2001.05.018 |
| [22] |
Raven J A. Endogenous inorganic carbon sources in plant photosynthesis. New Phytologist, 1972, 71(2): 227-247. DOI:10.1111/j.1469-8137.1972.tb04069.x |
| [23] |
Wang F S, Liu C Q, Wang B L, Liu X L, Li G R, Guan J, Yao C C, Wu Y Y. Disrupting the riverine DIC cycling by series hydropower exploitation in Karstic area. Applied Geochemistry, 2011, 26 Suppl: S375-S378. |
| [24] |
温周瑞, 徐军, 谢平. 太湖高等水生植物稳定碳、氮同位素特征. 湖泊科学, 2016, 28(1): 163-169. |
| [25] |
Van T K, Haller W T, Bowes G. Comparison of the photosynthetic characteristics of three submersed aquatic plants. Plant Physiology, 1976, 58(6): 761-768. DOI:10.1104/pp.58.6.761 |
| [26] |
林清. 温度和无机碳浓度对龙须眼子菜(Potamogeton pectinatus)碳同位素分馏的影响. 生态学报, 2008, 28(2): 570-576. DOI:10.3321/j.issn:1000-0933.2008.02.015 |
| [27] |
刘勇丽.太湖流域水生植物稳定碳氮同位素的生态学研究[D].湘潭: 湘潭大学, 2015: 31-37. http://cdmd.cnki.com.cn/Article/CDMD-10530-1015620448.htm
|
| [28] |
Hinga K R, Arthur M A, Pilson M E Q, Whitaker D. Carbon isotope fractionation by marine phytoplankton in culture:The effects of CO2 concentration, pH, temperature, and species. Global Biogeochemical Cycles, 1994, 8(1): 91-102. DOI:10.1029/93GB03393 |
| [29] |
Thode H G, Shima M, Rees C E, Krishnamurty K V. Carbon-13 isotope effects in systems containing carbon dioxide, bicarbonate, carbonate, and metal ions. Canadian Journal of Chemistry, 1965, 43(3): 582-595. |
| [30] |
Allemand D, Furla P, Bénazet-Tambutté S. Mechanisms of carbon acquisition for endosymbiont photosynthesis in Anthozoa. Canadian Journal of Botany, 1998, 76(6): 925-941. DOI:10.1139/b98-086 |
| [31] |
Sharkey T D, Berry J A, Raschke K. Starch and sucrose synthesis in Phaseolus vulgaris as affected by light, CO2, and abscisic acid. Plant Physiology, 1985, 77(3): 617-620. |
| [32] |
Trimborn S, Lundholm N, Thoms S, Richter K U, Krock B, Hansen P J, Rost B. Inorganic carbon acquisition in potentially toxic and non-toxic diatoms:the effect of pH-induced changes in seawater carbonate chemistry. Physiologia Plantarum, 2008, 133(1): 92-105. DOI:10.1111/j.1399-3054.2007.01038.x |
| [33] |
Benedict C R, Wong W W L, Wong J H H. Fractionation of the stable isotopes of inorganic carbon by seagrasses. Plant Physiology, 1980, 65(3): 512-517. DOI:10.1104/pp.65.3.512 |
| [34] |
Maberly S C. Diel, episodic and seasonal changes in pH and concentrations of inorganic carbon in a productive lake. Freshwater Biology, 1996, 35(3): 579-598. |
| [35] |
Xu J, Cao T, Zhang M, Li Z Q, Zhang M, Ni L Y, Xie P. Isotopic turnover of a submersed macrophyte following transplant:the roles of growth and metabolism in eutrophic conditions. Rapid Communications in Mass Spectrometry, 2011, 25(21): 3267-3273. DOI:10.1002/rcm.5224 |
| [36] |
Sackett W M, Eckelmann W R, Bender M L, Bé A W H. Temperature dependence of carbon isotope composition in marine plankton and sediments. Science, 1965, 148(3667): 235-237. DOI:10.1126/science.148.3667.235 |
| [37] |
Xu J, Xie P, Zhang M, Zhou Q, Zhang L, Wen Z R, Cao T. Icefish (Salangidae) as an indicator of anthropogenic pollution in freshwater systems using nitrogen isotope analysis. Bulletin of Environmental Contamination and Toxicology, 2007, 79(3): 323-326. |
| [38] |
张金美, 张萌, 匡武名, 祝国荣, 王玉玉, 卢龙, 徐军. 水华条件下鄱阳湖区植物叶片碳氮同位素特性. 环境科学研究, 2016, 29(5): 708-715. |
| [39] |
Hu X P, Burdige D J, Zimmerman R C. δ13C is a signature of light availability and photosynthesis in seagrass. Limnology and Oceanography, 2012, 57(2): 441-448. DOI:10.4319/lo.2012.57.2.0441 |
| [40] |
梁红, 黄林培, 陈光杰, 康文刚, 刘园园, 王教元, 朱庆生, 刘术, 邓颖. 滇东湖泊水生植物和浮游生物碳、氮稳定同位素与元素组成特征. 湖泊科学, 2018, 30(5): 1400-1412. |
| [41] |
Caut S, Angulo E, Courchamp F. Variation in discrimination factors (Δ15N and Δ13C):the effect of diet isotopic values and applications for diet reconstruction. Journal of Applied Ecology, 2009, 46(2): 443-453. DOI:10.1111/j.1365-2664.2009.01620.x |
| [42] |
Bade D L, Pace M L, Cole J J, Carpenter S R. Can algal photosynthetic inorganic carbon isotope fractionation be predicted in lakes using existing models?. Aquatic Sciences, 2006, 68(2): 142-153. DOI:10.1007/s00027-006-0818-5 |
| [43] |
陈展彦, 武海涛, 王云彪, 吕宪国. 基于稳定同位素的湿地食物源判定和食物网构建研究进展. 应用生态学报, 2017, 28(7): 2389-2398. |
| [44] |
Wang B L, Liu C Q, Peng X, Wang F S. Mechanisms controlling the carbon stable isotope composition of phytoplankton in karst reservoirs. Journal of Limnology, 2013, 72(1): 127-139. |
| [45] |
章程, 谢运球, 宁良丹, 玉宏, 汪进良, 李凤. 桂林会仙岩溶湿地典型水生植物δ13C特征与固碳量估算. 中国岩溶, 2013, 32(3): 247-252. DOI:10.3969/j.issn.1001-4810.2013.03.001 |
| [46] |
李瑞, 于奭, 孙平安, 何师意, 原雅琼, 熊志斌. 贵州茂兰板寨水域水生植物δ13C特征及光合作用固定HCO3-碳量估算. 中国岩溶, 2015, 34(1): 9-16. |
| [47] |
Wang J Z, Gu B H, Ewe S M L, Wang Y, Li Y C. Stable isotope compositions of aquatic flora as indicators of wetland eutrophication. Ecological Engineering, 2015, 83: 13-18. DOI:10.1016/j.ecoleng.2015.06.007 |
| [48] |
刘卫国, 李祥忠, 王政, 王欢业, 刘虎, 张博, 张欢. 西北干旱区湖泊碳同位素与环境变化. 中国科学:地球科学, 2019, 49(8): 1182-1196. |
| [49] |
Edwards T W D, McAndrews J H. Paleohydrology of a Canadian Shield lake inferred from 18O in sediment cellulose. Canadian Journal of Earth Sciences, 1989, 26(9): 1850-1859. DOI:10.1139/e89-158 |
| [50] |
陈毅风, 万国江. 泸沽湖沉积物α纤维素的提取及其稳定碳同位素研究初探. 地质地球化学, 1999, 27(4): 72-76. |
| [51] |
Street-Perrott F A, Holmes J A, Robertson I, Ficken K J, Koff T, Loader N J, Marshall J D, Martma T. The Holocene isotopic record of aquatic cellulose from Lake Äntu Sinijärv, Estonia:Influence of changing climate and organic-matter sources. Quaternary Science Reviews, 2018, 193: 68-83. DOI:10.1016/j.quascirev.2018.05.010 |
| [52] |
Wolfe B B, Falcone M D, Clogg-Wright K P, Mongeon C L, Yi Y, Benkert B, Amour N A S, Mark W A, Edwards T W D. Progress in isotope paleohydrology using lake sediment cellulose. Journal of Paleolimnology, 2007, 37(2): 221-231. DOI:10.1007/s10933-006-9015-8 |
| [53] |
Heyng A M, Mayr C, Lücke A, Wissel H, Striewski B. Late Holocene hydrologic changes in northern New Zealand inferred from stable isotope values of aquatic cellulose in sediments from Lake Pupuke. Journal of Paleolimnology, 2014, 51(4): 485-497. DOI:10.1007/s10933-014-9769-3 |
| [54] |
靳蕾, 王中良. 区域生态环境演变的湿地沉积物纤维素同位素研究进展. 环境科学与技术, 2012, 35(11): 26-31. DOI:10.3969/j.issn.1003-6504.2012.11.006 |
| [55] |
赵业思, 王建, 商志远. 树轮不同组分稳定碳同位素对气候变化响应敏感性:研究进展与评述. 生态学杂志, 2014, 33(9): 2538-2547. |
| [56] |
Yapp C J, Epstein S. Climatic significance of the hydrogen isotope ratios in tree cellulose. Nature, 1982, 297(5868): 636-639. DOI:10.1038/297636a0 |
| [57] |
金相灿, 屠清瑛. 湖泊富营养化调查规范(第二版). 北京: 中国环境科学出版社, 1990: 33-51.
|
| [58] |
Gleixner G, Danier H J, Werner R A, Schmidt H L. Correlations between the 13C content of primary and secondary plant products in different cell compartments and that in decomposing basidiomycetes. Plant Physiology, 1993, 102(4): 1287-1290. |
| [59] |
Taylor A M, Brooks J R, Lachenbruch B, Morrell J J, Voelker S. Correlation of carbon isotope ratios in the cellulose and wood extractives of Douglas-fir. Dendrochronologia, 2008, 26(2): 125-131. DOI:10.1016/j.dendro.2007.05.005 |
| [60] |
Eglin T, Maunoury-Danger F, Fresneau C, Lelarge-Trouverie C, Pollet B, Catherine L, Francois C, Damesin C. Biochemical composition is not the main factor influencing variability in carbon isotope composition of tree rings. Tree Physiology, 2008, 28(11): 1619-1628. DOI:10.1093/treephys/28.11.1619 |
| [61] |
Benner R, Fogel M L, Sprague E K, Hodson R E. Depletion of 13C in lignin and its implications for stable carbon isotope studies. Nature, 1987, 329(6141): 708-710. DOI:10.1038/329708a0 |
| [62] |
刘贤赵, 张勇, 宿庆, 田艳林, 全斌, 王国安. 现代陆生植物碳同位素组成对气候变化的响应研究进展. 地球科学进展, 2014, 29(12): 1341-1354. |
| [63] |
Anchukaitis K J, Evans M N, Lange T E, Smith D R, Leavitt S, Schrag D P. Consequences of a rapid cellulose extraction technique for oxygen isotope and radiocarbon analyses. Analytical Chemistry, 2008, 80(6): 2035-2041. |