 2020, Vol. 40
2020, Vol. 40文章信息
- 马建业, 李占斌, 马波, 李朝栋, 肖俊波, 张乐涛
- MA Jiangye, LI Zhanbin, MA Bo, LI Chaodong, XIAO Junbo, ZHANG Letao
- 黄土区小流域植被类型对沟坡地土壤水分循环的影响
- Effects of vegetation types in small watershed on soil water cycle in gully-slope land of loess region
- 生态学报. 2020, 40(8): 2698-2706
- Acta Ecologica Sinica. 2020, 40(8): 2698-2706
- http://dx.doi.org/10.5846/stxb201902120248
-
文章历史
- 收稿日期: 2019-02-12
- 网络出版日期: 2019-12-31





2. 广西壮族自治区水利水电勘察设计研究院, 南宁 530000;
3. 河南大学环境与规划学院, 开封 475004
2. Guangxi Zhuang Autonomous Region Water Conservancy and Hydropower Investigation and Design Institute, Nanning 530000, China;
3. College of Environment and Planning, Henan University, Kaifeng 475004, China
黄土高原地区生态环境脆弱, 水土流失严重, 水资源匮乏。降水作为该地区主要的输入水源, 其是如何穿过深厚的黄土包气带补给地下水是学者们长期以来所关心的问题[1], 深入揭示黄土中水分运移机制对于研究降水入渗补给地下水过程以及评价土壤水库的稳定性至关重要。近年来, 随着大面积的退耕还林(草)措施以及淤地坝建设的实施, 导致流域下垫面条件发生了较大的变化, 对流域水循环过程也产生了深刻影响[2]。如徐飞等[3]发现下垫面会通过改变蒸发和入渗方式影响山区水文循环。夏军等[4]则研究了下垫面特征对流域产汇流的影响。环境同位素作为有效的天然示踪剂, 能够有效地识别和反映流域不同水体特征, 识别土壤的入渗、蒸发、蒸腾等一些其他技术获取较为困难水文过程信息, 因而在研究土壤水分运移过程方面具有较大的优势, 也是研究流域水循环的重要手段[5]。如Liu等[6]研究发现降水在土壤中的入渗补给深度不会超过2—3 m。Song等[7]利用氢氧同位素对太行山典型的降水-土壤水-地下水转化关系进行了研究。但目前多集中于土壤水分来源以及与其他水体转化的定性分析。地形地貌条件作为影响土壤水分运移的重要因素之一, 在小流域尺度以沟缘线为界, 可将坡面分为梁峁坡和沟坡地[8], 沟坡地具有水分条件较好, 多松散碎屑物, 水土流失强度较大等特点[9-10], 这一区域的土壤水分可获得性被认为是制约植被生存与生长的关键因子[10], 也是土壤侵蚀需要重点治理的区域, 有利于从径流的形成区对流域侵蚀进行控制[11], 但目前有关土壤水分研究主要集中在梁峁坡, 而对沟坡土壤水分循环的研究较少。因此, 为深入了解不同下垫面植被类型对沟坡土壤水分循环特征影响, 本研究以黄土丘陵区纸坊沟小流域的刺槐林和草地为研究对象, 基于氢氧同位素示踪技术, 对0—100 cm土壤水的运移过程、水分来源以及对地下水的补给比例方面进行定量估算, 揭示草地和刺槐林对土壤水分运移的影响, 可为沟坡地生态修复措施的选择以及评价提供参考依据。
1 材料与方法 1.1 研究区概况纸坊沟流域(36°43′—36°46′ N, 109°14′—109°16′ E)位于陕西省安塞县境内, 是延河支流杏子河下游的一级支沟, 流域面积8.27 km2, 呈南北向狭长形, 平均海拔在1041—1425 m, 属暖温带半干旱季风气候区, 年均气温8.8 ℃, 年日总时数为2145 h, 年辐射总量为493 kJ/cm2, 年均蒸发量1463 mm, 年均降水量549.1 mm, 降水年际变化大且年内分配不均, 7—9月份的降水量占全年降水量的61.1%。主要土壤类型为黄绵土, 地带性土壤为黑垆土。潜水含水层主要为第四系冲积层潜水和浅层风化壳裂隙水[11]。地形破碎, 沟壑密度为8.07 km/km2, 先后经历了植被严重破坏期、继续破坏期、不稳定期、稳定恢复改善期和良性生态初步形成期, 下垫面主要植被类型发生了较大的变化, 影响了流域水循环过程[12]。
1.2 样品采集本研究选择纸坊沟流域沟坡主要下垫面植被类型刺槐林和草地为研究对象[12], 分别选择刺槐林地和草地各2块, 刺槐林地样地编号分别为B—1, B—2, 草地样地编号分别为G—1, G—2, 样地面积均设定为100 m2, 具体信息详见表 1。分别于2016年6月至11月对样地0—100 cm的土壤样品以及样地附近的降水、地表水、地下水样品进行采集, 采样周期为30 d, 若遇着连续降雨, 则进行区间加测。
| 植被类型 Vegetation types |
样地 Sample plot |
经纬度 Longitude and Latitude |
地形 Topography |
坡度 Slope |
海拔A ltitude/m |
土壤 Soil type |
主要植物种 Major plant species |
生长年限 Growth years/a |
| 荒草地 Grassland |
G—1 | 36°44′29.86″N 109°14′48.60″E |
沟坡 | 18° | 1163 | 黄绵土 | 铁杆蒿+茭蒿+白羊草 Artemisia gmelinii+ Artemisia giraldii+ Bothriochloai schaemum |
18 |
| G—2 | 36°45′41.08″N 109°15′44.43″E |
10° | 1083 | 黄绵土 | 白羊草+艾蒿 Bothriochloai schaemum+ Artemisia giraldii |
12 | ||
| 刺槐林 Black locust forest |
B—1 | 36°45′06.72″N 109°15′24.16″E |
沟坡 | 12° | 1125 | 黄绵土 | 刺槐Robiniap seudoacacia | 32 |
| B—2 | 36°45′05.89″N 109°15′17.60″E |
8° | 1116 | 黄绵土 | 刺槐Robiniap seudoacacia | 32 |
具体为在每个样地各布设1个J 16022型雨量筒用来收集降水样品。布设前, 向雨量筒内雨水收集器中加入适量液体石蜡油防止水分蒸发, 采样时使用15 mL一次性注射器抽取石蜡油层以下的降水[13], 共需布设4个雨量筒。地表水分别于样地下部邻近的沟道内采集, 采集时将样品瓶伸入水面以下30 cm处, 以防止水面蒸发分馏的影响, 由于B—1样地和B—2样地位于沟道同一断面, 因此共布设3个地表水采样点。且该流域只有沟口存在居民饮用水井, 井深约为13 m, 为地下潜水, 因此地下水样品于沟口进行采集, 取样时先用水桶将地下水提升至地面, 于阴凉处迅速取样, 取样方法与地表水一致, 共布设1个采样点。上述水样每次采集均做2次重复, 采集的水样盛装于15 mL棕色玻璃试剂瓶中, 拧紧瓶盖并用封口膜密封, 以防止水分蒸发损失, 采样点布设见图 1。在研究时段内共采集降水样品48个, 地表水样品36个, 地下水样品12个。

|
| 图 1 研究区位置及采样点分布 Fig. 1 Study area location and sampling point distribution |
在4个样地坡面上, 使用土钻在坡中位置选择2个采样点, 分别采集0—5, 5—10, 10—15, 15—20, 20—40, 40—60, 60—80, 80—100 cm共8个土层的土样, 分析时以每个土层范围的平均深度为标准, 即2.5, 7.5, 12.5, 17.5, 30, 50, 70, 90 cm。采集的土壤样品盛装于20 mL棕色玻璃试剂瓶中, 密封并冷藏, 防止蒸馏作用对土壤水氢氧同位素组成造成影响。在研究时段内共采集刺槐林土壤样270个, 荒草地土壤样品265个。
1.3 数据处理(1) 土壤水抽提。将采集到的土壤样品带回西安理工大学水资源研究所实验室, 使用LI-2000液态水真空抽提系统(LICA, China)进行土壤水抽提。
(2) 氢氧同位素测定。水样的氢氧同位素测定在西安理工大学同位素分析实验室进行。使用液态水同位素分析仪(DLT—100, LGR公司, 美国)测定水样样品的氢氧同位素。δD和δ18O同位素比值是通过相对于维也纳标准平均海水(VSMOW)的千分率(‰)计算, 其精度分别为0.5‰和0.15‰。公式为:
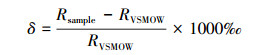
|
(1) |
式中, Rsample为水样中D/H或18O/16O的比率, 而RVSMOW为VSMOW标准水样D/H或18O/16O的比率。通过不同水源同位素的对比, 应用二端元混合模型, 可知水体的水分来源比例[14-15]:
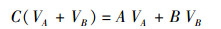
|
(2) |
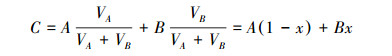
|
(3) |
式中, A:降水氢氧同位素值/‰;B:地表水/地下水氢氧同位素值/‰;C:土壤水氢氧同位素值/‰;x:地表水/地下水所占比例/%;1-x:降水所占比例/%。
(3) 试验数据统计分析。数据分析采用Excel完成, 显著性分析由SPSS20.0完成, 所有图表均运用Excel2010和AutoCAD2007完成。
2 结果与分析 2.1 流域不同水体氢氧同位素统计特征由于水体来源以及经受的蒸发分馏程度不同, 致使各水体的氢氧同位素存在差异, 对流域不同水体氢氧同位素浓度进行统计分析, 如表 2所示。纸坊沟流域不同水体的δD和δ18O基本符合降水>刺槐林地土壤水>草地土壤水>地表水>地下水, 降水的同位素最富集, 变异系数均值最高(0.42), 地下水同位素最贫化, 变异系数均值较低(0.06), 这与赵宾华等[5]在王茂沟得到的结果一致。刺槐林地和草地土壤水变异系数相差较小, 分别为0.16和0.18。但各水体的氢氧同位素在统计学上差异均不显著(P>0.05), 可能是由于各水体之间连通性较好, 存在相互转化引起的。标准差反映了氢氧同位素值的离散程度, 降水的同位素离散程度最高, 可能是因为研究区地处西北内陆干旱区, 雨季温度较高, 降水在降落过程中受蒸发的影响, 重同位素逐渐富集, 且易受季风气候的影响, 致使降水同位素的最为富集且变异性以及离散程度较高[16]。而地下水由于不止受到雨季降水的补给, 还受到地表水和冬季同位素较为贫化的降雪的补给, 且黄土高原地区地下水埋藏较深, 蒸发分馏作用对其影响较小, 导致地下水同位素整体呈贫化现象[17]。而刺槐林地土壤水氢氧同位素较草地富集, 可能是由于刺槐具有较大的冠幅, 在降雨过程中存在二次截留现象, 延长了降水补给土壤水的过程, 增大了蒸发分馏作用, 致使林地土壤水同位素较为富集。
| 不同水体 Water bodies |
δD/‰ | δ18O/‰ | ||||||||
| 最大值 Max |
最小值 Min |
均值 Average |
标准差 Std |
最大值 Max |
最小值 Min |
均值 Average |
标准差 Std |
|||
| 降水Precipitation | 18.47 | 95.08 | 57.12 | 24.26 | 3.39 | 14.11 | 7.48 | 3.17 | ||
| 地表水Surface water | 59.44 | 65.40 | 62.72 | 2.03 | 7.65 | 9.68 | 8.47 | 0.48 | ||
| 地下水Groundwater | 56.25 | 66.02 | 63.47 | 2.46 | 6.69 | 8.91 | 8.87 | 0.73 | ||
| 土壤水Soil water | 草地 | 41.28 | 77.80 | 59.66 | 11.00 | 10.56 | 10.56 | 8.08 | 1.43 | |
| 刺槐林地 | 42.30 | 76.69 | 58.05 | 7.90 | 5.05 | 10.06 | 7.74 | 1.40 | ||
图 2为不同水体的氢氧同位素关系图。由图可知, 研究时段内的降水线方程为δD=7.65δ18O+6.77(R2=0.98), 斜率和截距均小于全球大气降水线(δD=8.0δ18O+10), 主要由于水汽从沿海向内陆传输过程中的二次分馏以及内陆水汽的补给所致。草地土壤水蒸发线方程为δD=6.96δ18O-3.41(R2=0.81), 刺槐林地蒸发线方程为δD=5.10δ18O-18.62(R2=0.81), 土壤水的斜率和截距均小于降水线, 说明降水是土壤水的主要补给来源, 且在降水补给土壤水之前经历了较强的非平衡蒸发作用[18], 刺槐林地的斜率和截距小于草地, 表明刺槐林地土壤水经受的蒸发作用较草地更强, 且草地土壤水更多来自降水补充, 刺槐林地接受前期土壤水补给的比例较大[19]。且刺槐林地和草地蒸发线在地下水和地表水所在区域附近相交, 说明地表水、地下水和土壤水存在相互补给的可能。

|
| 图 2 不同水体δD和δ18O关系 Fig. 2 The relationship between δD and delta δ18O in different water bodies |
土壤水氢氧同位素贫化峰值的运移对降水在土壤中的入渗过程具有较好的指示作用。本研究分别于7月27日、8月1日、8月6日对刺槐林地和草地的土壤水氢氧同位素进行了监测, 不同时间段土壤水氢氧同位素随土层深度的变化特征如图 3所示。表 3为监测间隔期间的降水量, 降雨形式均为短阵型暴雨。G—1样地中, 7月27日的δ18O贫化峰值出现在7.5 cm处, 8月1日和8月6日的贫化峰值出现在17.5 cm处, 水分入渗速率为2 cm/d。G—2样地中, 7月27日、8月1日、8月6日δ18O贫化峰值分别出现在7.5 cm、17.5 cm、35 cm处, 入渗速率分别为2 cm/d和3.5 cm/d。B—1样地中, 7月27日的δ18O贫化峰值出现在12.5 cm处, 8月1日和8月6日的贫化峰值出现在17.5 cm处, 水分入渗速率为1 cm/d。B—2样地中, 7月27日、8月1日、8月6日δ18O贫化峰值分别出现在7.5 cm、17.5 cm、35 cm处, 入渗速率分别为2 cm/d和3.5 cm/d。综上, 在短阵型暴雨条件下, 草地土壤水分入渗速率较林地高1 cm/d, 且入渗主要发生在0—50 cm土层。一方面可能是因为冠层的截留作用, 使得落到林地土壤表面的降水强度和降水量偏低, 而土壤水分入渗速率与降雨强度具有较强的相关性[20], 导致刺槐林地土壤水分入渗速率偏低。而草地由于地表植被盖度较大, 对地表径流的阻滞作用较强, 使得更多的水分入渗到土壤中, 增大了水分入渗速率。另一方面, 耿韧等[21]在纸坊沟流域的研究结果表明刺槐林表层土壤孔隙度为51.83%, 以铁杆蒿和长茅草为主要群落的草地土壤孔隙度分别为56.05%和55.85%, 较刺槐林地偏高。且实地调查发现沟坡部位土层深厚的区域多以刺槐林为主, 而草地一般位于土层较薄, 且下部多碎石坡面, 进一步说明了草地坡面土壤孔隙结构较好, 有利于土壤水分入渗。但由于监测时段内的降雨主要为短阵性暴雨, 大部分以地表径流的形式汇入到沟道流失, 少部分水分入渗至土壤中, 消减了土壤水分入渗速率的差异。

|
| 图 3 8月份土壤水分运移过程 Fig. 3 Soil water transport in August 图中B为刺槐林地(Black locust forest), C为草地样地(Grassland) |
| 时间Time | 7.23—7.27 | 7.27—8.1 | 8.1—8.6 |
| 降水量Precipitation/mm | 7 | 10 | 7 |
降水降落到地面进入土壤以后, 除部分供给植物正常生命活动和蒸发消耗外, 其余部分会继续以活塞流和优先流的形式向深层入渗, 补给深层土壤水和地下水。而在干旱时节, 表层土壤水分亏缺, 地下水和深层土壤水会以水或汽的形式沿毛细带向上层运移补充表层土壤水, 以保证植物的正常生长。然而土壤水-地下水之间的转化并不是普遍存在的, 本研究分别挑选发生土壤水对地下水补给的事件以及地下水补给土壤水的事件, 统计各事件的不同水体氢氧同位素均值, 用于探究土壤水与地下水之间的相互转化比例。表 3为降水和地下水对0—100 cm土层土壤水的补给比例。由表可知, B—1样地中, 降水和地下水对0—100 cm土壤水的补给比例分别为44.87%和55.13%, B—2样地中, 补给比例分别为33.20%和66.80%, G—1样地中, 降水和地下水对土壤水的补给比例分别为50.04%和49.96%, G—2样地的补给比例分别为68.20%和31.80%。综上, 刺槐林地0—100 cm土壤水中约有39.03%来自于降水, 60.97%来自于地下水补给, 草地土壤水中降水和地下水的补给比例分别为59.12%和40.88%, 这与图 2得到草地土壤水主要受降水补给, 林地土壤水中前期土壤水所占比例较高的结论一致。表 4为不同植被类型下土壤水对地下水的补给比例, 由表可知, 只在B—1和G—1样地中估算得到了土壤水补给地下水的比例, 分别为51.12%和51.64%, 而在B—2和G—2样地中未估算得到土壤水对地下水发了生补给, 说明短期内土壤水对地下水的补给并不是普遍存在的, 可能与土壤内部结构以及地质条件相关。综上, 草地土壤水对地下水的补给比例较刺槐林地高0.52%, 差异较小。
| 植被类型 Vegetation types |
样地编号 Sample number |
δ18O | 补给比例Conversion ratio/% | ||||
| 降水 Precipitation |
土壤水 Soil water |
地下水 Groundwater |
降水 Precipitation |
地下水 Groundwater |
|||
| 刺槐林地 | B—1 | 7.93 | 8.03 | 7.02 | 44.87 | 55.13 | |
| Black locust forest | B—2 | 6.33 | 7.20 | 9.07 | 33.20 | 66.80 | |
| 均值 | 39.03 | 60.97 | |||||
| 草地Grassland | G—1 | 7.51 | 8.03 | 8.55 | 50.04 | 49.96 | |
| G—2 | 6.33 | 8.16 | 9.07 | 68.20 | 31.80 | ||
| 均值 | 59.12 | 40.88 | |||||
| 植被类型 Vegetation types |
样地编号 Sample number |
δ18O | 补给比例Conversion ratio/% | ||||
| 土壤水 Soil water |
地表水 Surface water |
地下水 Groundwater |
土壤水 Soil water |
地表水 Surface water |
|||
| 刺槐林地 | B—1 | 9.80 | 8.31 | 9.07 | 51.12 | 48.88 | |
| Black locust forest | B—2 | — | — | — | — | — | |
| 均值 | 51.12 | 54.52 | |||||
| 草地Grassland | G—1 | 9.48 | 7.79 | 8.67 | 51.64 | 48.36 | |
| G—2 | — | — | — | — | — | ||
| 均值 | 51.64 | 48.36 | |||||
在黄土丘陵沟壑区, 沟坡是小流域尺度上水土流失最为活跃的部分, 了解其水分特征对于沟坡地的综合治理具有重要作用[11]。而不同的下垫面植被类型, 会影响土壤水分入渗以及产汇流等水文过程[22-23], 因此探究不同植被类型下土壤水分来源和去向对于土壤水库稳定性评估以及生态修复植物类型的选择具有重要指导作用[24]。本研究中, 刺槐林地0—100 cm土壤水中降水所占的比例较草地低9.51%, 一方面可能是因为草本植物群落缺乏较大冠层对降雨进行截留, 到达地面的降水量较大, 使得更多的水分通过入渗的方式进入土壤。同时草地土壤较好的土壤孔隙结构和较大的入渗速率, 水分能够较快的入渗至下层土壤, 降低蒸发损失, 使得草地土壤水中降水的比例较刺槐林地偏大。黄土高原地区包气带深厚, 地下水埋深较大, 一般在20 m以下, 地下水沿着土壤毛管孔隙上升补给0—100 cm土层土壤存在较大困难。然而本研究结果表明部分样地存在地下水补给表层土壤水的可能, 一方面可能是因为研究样地位于流域沟坡, 土层较薄, 且靠近沟道底部, 地下水埋深较浅, 存在地下水向上补给的可能。另一方面可能是因为本研究中的对土壤水发生补给的地下水实质上是深层土壤水, 据程立平等[25]和Tan等[26]在黄土高原的研究结果分析发现, 在4 m以下的土壤剖面氢氧同位素较为接近于地下水, 而端元模型是基于输入单元与输出单元的估算模型, 未涉及到具体补给过程, 可能造成信息误差, 为验证这一结果, 本研究基于已有的研究数据, 以50—100 cm土层土壤水表征深层土壤水, 以0—50 cm土层土壤水表征表层土壤水, 对0—50 cm土层土壤水接受50—100 cm土层土壤水补给的比例进行了估算, 结果表明, 0—50 cm土层土壤水中, 50—100 cm土层土壤水补给所占比例为46.87%, 较为接近地下水对0—100 cm土层土壤水补给的平均比例(50.93%), 说明存在深层土壤水补给表层土壤水的可能, 因此可推断本研究中对0—100 cm土层发生补给的地下水本质上为深层土壤水。本研究中刺槐林地深层土壤水对表层土壤水的补给比例较草地的比例高9.51%, 可能是因为刺槐根系分布深度较大, 对20 cm以下的土壤水大量消耗, 土水势较强, 并且刺槐林所在区域土壤多为均质黄土, 土壤孔隙较小, 且沟坡地下水位较高, 下层土壤水分含量较高, 在土水势差的作用下, 深层土壤水向上层运移, 加上植物根系对土壤水分的吸收, 容易在深层出现土壤水分干层[27]。草地根系分布深度较浅, 下层土壤含水率高, 土壤孔隙较大, 毛管孔隙力小, 不利于深层土壤水分向上层运移。
黄土高原地区地下水主要来源于降水入渗穿过包气带进行土壤水深层运移补给, 但也可通过径流汇聚在低洼处再在局部地区进行地下水补给[28], 这从另一方面验证了纸坊沟流域地下水可能来源于土壤水与地表水的共同补给这一假设。同时本研究发现草地土壤水对地下水补给比例较刺槐林地高0.52%, 但差异较小。这可能与包气带土壤水分运移及其地下水补给具有很强的尺度依赖性有关[29], 在小流域尺度, 主要受地形和地貌影响, 而在区域尺度上, 则受到植被覆盖因素影响较大[30], 可能B—1样地和B—2样地距离太近, 地形地貌条件没有明显的差异, 造成土壤水对地下水的补给比例差异较小。而由于刺槐等乔木树种根系较为发达, 对深层土壤水分的利用强度较大, 在干旱季节几乎没有水分能够接受补给, 而草地深层土壤水分利用较少, 能够发生深层运移的可能, 造成草地对地下水的补给较刺槐林地偏高。同时在B—2和G—2样地未发现明显的土壤水补给地下水的现象, 可能与土壤内部的孔隙通道有关, 程立平等[25]研究发现黄土孔隙和裂隙是黄土地下水贮存和运移的主要场所, 形体大、数量少、分布不均、连通性较好的裂隙和空洞则成为黄土中地下水的主导运移通道。这些孔隙通道的形成条件不仅与植物的根系相关, 还受地质地貌条件以及土壤内部等多种因素的影响, 并不是所有的孔隙通道均具有连通性, 造成不同样地对地下水的补给情况出现较大的差异。综上可以发现沟坡地下垫面植被类型由草地转为刺槐林的过程中, 可能发生地下水补给减少的情况, 但变幅较小。
4 结论本研究基于氢氧同位素分析了纸坊沟流域沟坡地不同植被类型下0—100 cm土壤水的入渗速率、水分来源与去向等科学问题, 得到以下结论:
(1) 与草地相比, 刺槐林地土壤水的氢氧同位素更为富集, 受到的蒸发分馏作用更强。草地和刺槐林地土壤水蒸发线方程为:δD=6.96δ18O-3.41, δD=5.10δ18O-18.62。在短阵型暴雨条件下, 草地土壤水分入渗速率较刺槐林地高1 cm/d, 且入渗主要发生在0—50 cm土层。
(2) 草地土壤水中以降水补给为主, 所占比例为59.12%, 刺槐林地中以深层土壤水补给为主, 所占比例为60.97%, 说明由草地转为刺槐林地可能引起深层土壤的干层现象发生。地下水补给以沟坡地土壤水的垂向运移为主要补给形式, 草地土壤水对地下水的补给比例较刺槐林地高0.52%, 无显著差异。
| [1] |
李玉山. 黄土高原森林植被对陆地水循环影响的研究. 自然资源学报, 2001, 16(5): 427-432. DOI:10.3321/j.issn:1000-3037.2001.05.006 |
| [2] |
常恩浩, 李鹏, 张铁钢, 肖列, 徐国策, 赵宾华, 张祎. 旱季雨季对黄土丘陵退耕区植被根系分布及水分利用的影响. 农业工程学报, 2016, 32(24): 129-138. DOI:10.11975/j.issn.1002-6819.2016.24.017 |
| [3] |
徐飞, 贾仰文, 牛存稳, 刘佳嘉, 韩春苗. 北京清水河流域水循环垂直地带性研究. 水电能源科学, 2017, 35(8): 1-5. |
| [4] |
夏军, 乔云峰, 宋献方, 叶爱中, 张学成. 岔巴沟流域不同下垫面对降雨径流关系影响规律分析. 资源科学, 2007, 29(1): 70-76. DOI:10.3321/j.issn:1007-7588.2007.01.011 |
| [5] |
赵宾华, 李占斌, 李鹏, 肖列, 常恩浩, 张祎, 高蓓. 黄土区生态建设对流域不同水体转化影响. 农业工程学报, 2017, 33(23): 179-187. DOI:10.11975/j.issn.1002-6819.2017.23.023 |
| [6] |
Liu W Z, Zhang X C, Dang T H, Ouyang Z, Li Z, Wang J, Wang R, Gao C Q. Soil water dynamics and deep soil recharge in a record wet year in the southern Loess Plateau of China. Agricultural Water Management, 2010, 97(8): 1133-1138. DOI:10.1016/j.agwat.2010.01.001 |
| [7] |
Song X F, Wang P, Yu J J, Liu J R, Yuan R Q. Relationships between precipitation, soil water and groundwater at Chongling catchment with the typical vegetation cover in the Taihang mountainous region, China. Environmental Earth Sciences, 2011, 62(4): 787-796. DOI:10.1007/s12665-010-0566-7 |
| [8] |
李小曼, 王刚, 李锐. 基于DEM的沟缘线和坡脚线提取方法研究. 水土保持通报, 2008, 28(1): 69-72. |
| [9] |
杨新民, 杨文治, 马玉玺. 纸坊沟流域人工刺槐林生长状况与土壤水分条件研究. 水土保持研究, 1994, 1(3): 31-35, 42. |
| [10] |
胡相明, 程积民, 万惠娥, 赵艳云. 黄土丘陵区地形、土壤水分与草地的景观格局. 生态学报, 2006, 26(10): 3276-3285. DOI:10.3321/j.issn:1000-0933.2006.10.017 |
| [11] |
黄奕龙, 陈利顶, 傅伯杰, 黄志霖, 巩杰, 吴祥林. 黄土丘陵小流域沟坡水热条件及其生态修复初探. 自然资源学报, 2004, 19(2): 183-189. DOI:10.3321/j.issn:1000-3037.2004.02.007 |
| [12] |
张瑞.黄土高原土壤无机碳密度分布、储量及影响因素[D].西安: 中国科学院研究生院(教育部水土保持与生态环境研究中心), 2012.
|
| [13] |
王贺, 李占斌, 马波, 马建业, 张乐涛. 黄土高原丘陵沟壑区流域不同水体氢氧同位素特征——以纸坊沟流域为例. 水土保持学报, 2016, 30(4): 85-90, 135-135. |
| [14] |
Yeh H F, Lin H I, Lee C H, Hsu K C, Wu C S. Identifying seasonal groundwater recharge using environmental stable isotopes. Water, 2014, 6(10): 2849-2861. DOI:10.3390/w6102849 |
| [15] |
张翔, 李占斌, 李鹏. 淤地坝土壤水来源与分布研究. 西安理工大学学报, 2013, 29(1): 1-7. DOI:10.3969/j.issn.1006-4710.2013.01.001 |
| [16] |
王锐, 刘文兆, 宋献方. 黄土塬区土壤水分运动的氢氧稳定同位素特征研究. 水土保持学报, 2014, 28(3): 134-137, 184-184. |
| [17] |
姚俊强, 刘志辉, 郭小云, 王月健, 郑江华, 房世峰. 呼图壁河流域水体氢氧稳定同位素特征及转化关系. 中国沙漠, 2016, 36(5): 1443-1450. |
| [18] |
张北赢.黄土丘陵区小流域不同土地利用方式土壤水分动态规律研究[D].西安: 中国科学院研究生院(教育部水土保持与生态环境研究中心), 2008.
|
| [19] |
马建业, 李占斌, 马波, 王贺, 张乐涛, 李朝栋. 黄土高原丘陵区不同植被恢复方式下土壤水分特征——以桥子沟流域为例. 中国水土保持科学, 2017, 15(4): 8-15. |
| [20] |
王新平, 李新荣, 康尔泗, 张景光, 周海燕, 杨诗秀, 雷志栋. 腾格里沙漠东南缘人工植被区降水入渗与再分配规律研究. 生态学报, 2003, 23(6): 1234-1241. DOI:10.3321/j.issn:1000-0933.2003.06.027 |
| [21] |
耿韧, 张光辉, 李振炜, 胡国芳, 王浩, 朱良君. 基于分层抽样法的小流域土壤物理性质和有机质差异特征. 水土保持学报, 2014, 28(6): 194-199, 205-205. |
| [22] |
李燕.乌兰布和沙漠水汽热耦合运移规律及植被生态学意义[D].西安: 长安大学, 2013.
|
| [23] |
李民义, 张建军, 王春香, 王丹丹. 晋西黄土区不同土地利用方式对土壤物理性质的影响. 水土保持学报, 2013, 27(3): 125-130, 137-137. |
| [24] |
赵娜娜, 李传哲, 刘佳, 穆文彬, 于福亮, 邢九平. 不同下垫面的土壤水分时间稳定性分析. 南水北调与水利科技, 2015, 13(4): 765-770. |
| [25] |
程立平, 刘文兆.黄土高原地区地下水资源研究//中国地理学会2009百年庆典学术大会论文集.北京: 中国地理学会, 2009.
|
| [26] |
Tan H B, Liu Z H, Rao W B, Jin B, Zhang Y D. Understanding recharge in soilgroundwater systems in high loess hills on the Loess Plateau using isotopic data. CATENA, 2017, 156: 18-29. DOI:10.1016/j.catena.2017.03.022 |
| [27] |
侯庆春, 韩蕊莲, 韩仕锋. 黄土高原人工林草地"土壤干层"问题初探. 中国水土保持, 1999(5): 13-16. |
| [28] |
Favreau G, Leduc C, Marlin C, Dray M, Taupin J D, Massault M, Le Gal La Salle C, Babic M. Estimate of recharge of a rising water table in semiarid Niger from 3H and 14C modeling. Groundwater, 2002, 40(2): 144-151. DOI:10.1111/j.1745-6584.2002.tb02499.x |
| [29] |
Bormann H, Klaassen K. Seasonal and land use dependent variability of soil hydraulic and soil hydrological properties of two Northern German soils. Geoderma, 2008, 145(3/4): 295-302. |
| [30] |
赵文智, 周宏, 刘鹄. 干旱区包气带土壤水分运移及其对地下水补给研究进展. 地球科学进展, 2017, 32(9): 908-918. |