 2020, Vol. 40
2020, Vol. 40文章信息
- 马源, 杨洁, 张德罡, 周恒, 周会程, 陈建纲
- MA Yuan, YANG Jie, ZHANG Degang, ZHOU Heng, ZHOU Huicheng, CHEN Jiangang
- 高寒草甸退化对祁连山土壤微生物生物量和氮矿化速率的影响
- Effects of alpine meadow degradation on soil microbial biomass and nitrogen mineralization rate in the Qilian Mountains
- 生态学报. 2020, 40(8): 2680-2690
- Acta Ecologica Sinica. 2020, 40(8): 2680-2690
- http://dx.doi.org/10.5846/stxb201902010231
-
文章历史
- 收稿日期: 2019-02-01
- 网络出版日期: 2019-12-31





草地是陆地生态系统中最重要的组成部分, 占据了我国国土面积的40%左右[1]。近几十年, 由于人们对草原的过度放牧和草地资源的过度开垦, 使高寒草甸区植物群落的多样性发生显著的变化, 从而导致草地退化面积不断扩大, 最终导致草地生态系统的紊乱[2]。据统计发现, 祁连山高寒草甸区60%左右的草原发生重度退化, 草地超载率达到70%左右, 这些问题的发生都必将造成草地初级生产力氮素的流失和土壤微生物量的周转率下降。氮元素作为限制草地营养转化的关键元素之一, 其重要性不言而喻[3], 土壤微生物量作为土壤有机物质的活性成分, 其周转率的快慢和对土壤养分变化敏感性, 对草地养分循环有显著作用, 是最主要的土壤养分库之一[4]。因此, 土壤铵态氮、土壤硝态氮以及土壤微生物量氮等物质是评价土壤生产力的关键指标, 对草地生态系统中氮素的调控和转化具有重要意义。
实际上, 研究人员早在30多年前已了解土壤微生物量是造成有机物分解、养分矿化和养分循环的主要因素[5]。在全球范围内对不同土壤类型、不同生态环境的土壤理化性质以及微生物特征的研究发现, 植物群落的改变与土壤有机碳和土壤总氮有明显的相关性, 这与植物凋落物对土壤的回馈有关[6], 反过来思考, 植物残体的转化过程必然影响土壤中微生物量的活跃性[7]。同时有研究表明, 过度开垦所导致的草地退化能够改变土壤微生物的群落结构, 使草地中土壤微生物量降低, 活性减少[8]。由于土壤微生物对土壤的变化相比较土壤有机质更具灵敏性, 因此, 通过分析土壤的微生物特性, 同样能够准确和快速的反映出土壤的变化信息[9]。土壤中氮元素的内循环是草地生态系统的关键功能, 他占全球氮植物需求的88%左右, 该过程将有机氮矿化为NH4+, 并通过氧化作用将NH4+氧化成亚硝酸盐和NO3-, 最后这些矿物形式的N通过微生物的同化作用被植物吸收[10]。土壤氮矿化是影响植物和微生物中无机氮有效性的最重要的内部生态系统过程之一, 而土壤氮的有效性则与土壤有机质分解过程中矿化或固定的N量有直接关系[11]。研究证实, 土壤微生物活性的增加与N矿化的增加和土壤潜在的氮元素供应有关[12-13]。由此可见, 草地退化过程中对土壤氮周转具有一定的影响, 并且在一定程度上影响草地生态系统中土壤氮储量和氮素循环, 进而影响土壤的氮供应潜力。现今, 国内外学者已在不同方面解释说明了退化高寒草甸的特征和退化机理, 但对于草原在退化过程中对氮矿化的影响等方面所知甚少, 且相关机理尚不明晰。
因此, 本研究通过选取祁连山东缘4个退化程度的高寒草甸草原(未退化草甸、轻度退化草甸、中度退化草甸、极度退化草甸), 开展随退化程度的加剧土壤氮矿化速率、土壤氮矿化相关的土壤微生物特性以及土壤因子和植被等进行相关研究, 以期揭示高寒草甸区草地退化过程中土壤氮矿化的主要因素和特点, 为退化草地生态恢复过程中氮素的供给提供一定的理论支撑。
1 材料与方法 1.1 样地概况研究地位于甘肃省祁连山东缘地区(102°44′11″—102°46′17″E, 37°11′42″—37°13′5″N), 该样地类型为寒温潮湿高山草甸类, 土壤类型为亚高山草甸土, 土壤含水量(50%—80%), 土壤pH值(6.94—8.17)。海拔(2869.8—3008.3 m), 年均温(-0.1—0.6 ℃), 全年≥0℃的积温为1360℃左右, 降雨量446 mm左右, 主要集中在7—9月, 年均蒸发量(1483—1614 mm), 气候寒冷潮湿, 水热同期, 平均相对湿度55%, 年日照时间2600 h。主要植物种类包括:珠芽蓼(Polygonum viviparum)、早熟禾(Poa annua)、披碱草(Elymus dahuricus)、线叶嵩草(Kobresia capillifolia)、矮嵩草(Kobresia humilus)、甘肃棘豆(Oxytropis kansuensis)、唐松草(Thalictrum spp)、萎陵菜(Potentilla spp)、凤毛菊(Saussurrea spp)、扁蓿豆(Melissilus ruthenicus)、乳白香青(Anaphalis lactea)、狼毒(Stellera chamaejasme)等, 样地概况见表 1。
| 指标 Factors |
退化程度Degradation grade | |||
| 未退化草地ND | 轻度退化草地LD | 中度退化草地MD | 极度退化草地SD | |
| 植物盖度Plant coverage/% | 96±2.31a | 82.67±2.33b | 65±3.18c | 37.33±2.60d |
| 植物物种数Plant species richness | 15±1.57a | 12±0.62b | 10±1.68bc | 8±1.84c |
| 植物高度Plant height/cm | 19.06±1.74a | 9.85±0.77b | 6.16±0.89c | 2.06±0.52d |
| 地上生物量Aboveground biomass/(g/m2) | 403.33±14.07a | 364.38±5.06b | 245.66±2.36c | 99.31±6.53d |
| 地下生物量Underground biomass/(g/m2) | 1569.63±30.52a | 723.17±31.64b | 1130.26±27.66c | 266.35±44.58d |
| 土壤有机碳Soil organic carbon/(g/kg) | 90.57±1.3a | 80.21±1.09b | 70.41±1.85c | 60.15±1.4d |
| 土壤全氮Total nitrogen/(g/kg) | 4.84±0.05a | 4.09±0.08b | 3.17±0.14c | 2.57±0.18d |
| 土壤含水量Soil moisture(SM)/% | 41.86±1.25a | 35.29±0.84b | 26.83±0.63c | 14.77±0.38d |
| pH | 8.00±0.10a | 7.76±0.07a | 7.44±0.05b | 6.99±0.08c |
| 不同小写字母表示不同退化程度间差异性显著(P < 0.05), ND:未退化Non-degraded grassland;LD:轻度退化Light degraded grassland;MD:中度退化Moderate degraded grassland;SD:极度退化Severely degraded grassland | ||||
2018年8月根据草地退化相关分级标准[14], 在甘肃省天祝县金强河草原站附近选取未退化草地(Non-degraded grassland, ND), 轻度退化草地(Light degraded grassland, LD), 中度退化草地(Moderate degraded grassland, MD)以及极度退化草地(Severely degraded grassland, SD)4个样地, 每个样地主要优势物种分别是, 未退化样地优势物种:珠芽蓼、垂穗披碱草、线叶嵩;轻度退化样地优势物种:线叶嵩草、矮生嵩草、扁蓿豆;中度退化样地优势物种:矮生嵩草、线叶嵩草、扁蓿豆;极度退化样地优势物种:乳白香青、矮生嵩草、披碱草。
每个样地设置1个大样方(50 m × 50 m)。并且在大样方的四个角和中心各设置一个小样方(1 m × 1 m), 用方格法测定每个小样方中植物高度, 植被盖度, 植物种类, 植物数量以及地上生物量, 并且选取3个小样方采集表层(0—10 cm)土壤样品, 每个样品分为2部分, 一部分在4℃下保存为新鲜土壤, 进行土壤铵态氮、硝态氮、微生物量和氮矿化速率的测定;另一部分阴干和均质化, 过2 mm的筛, 进行土壤理化性质分析(见表 1)。
1.3 指标测定与方法土壤有机碳(Soil organic carbon, SOC), 土壤全氮(Soil total nitrogen, TN), 土壤含水量(Soil moisture, SM)的测定依照《土壤分析技术规范》的方法[15];铵态氮(NH4+), 硝态氮(NO3-)参照鲍士旦[16]《土壤农化分析(第三版)》的方法;土壤pH用去CO2的蒸馏水(1:2.5土水比的浸提液)浸提后用pH计测定[15]。土壤样品中蛋白酶(Protease)、脲酶(Urease)、β-乙酰葡糖胺糖苷酶(β-N-acetyl glucosaminidase, NAG)和亮氨酸氨基肽酶(Leucine aminopeptidase, LAP)的活性通过使用苏州科铭生物公司的试剂盒进行测定。
土壤微生物生物量碳(Microbial biomass carbon, MBC)和微生物生物量氮(Microbial biomass nitrogen, MBN)采用氯仿熏蒸提取方法测量[17];通过调节土壤样品至田间持水量的50%后, 测量25℃土样在实验室培养中释放的CO2的量获得土壤基础呼吸(Soil basal respiration, SBR)[18], 方法如下, 将20 g新鲜土样密封并在黑暗中保持10 d, 后用5 mL 1 mol/L NaOH吸收放出的CO2, 最后通过1 mol/L HCl滴定过量的NaOH来测定;代谢熵(qCO2), 土壤基础呼吸量与单位土壤微生物量碳的比率[18];微生物熵(CMBC/ CSOC), 通过微生物生物量碳的含量与土壤有机碳的比率来计算[18]。
土壤净氮矿化, 硝化和氨化速率的测定参考Trap方案进行测定[19], 将新鲜土壤分为两部分A和B进行分析。称取10 g新鲜土样铵态氮(NH4+)和硝态氮(NO3-)的浓度作为培养前的初始值(A);称取10 g新鲜土样放入240 mL组培瓶, 用聚乙烯膜封口于25℃的培养室中培养10 d后分别测定铵态氮和硝态氮的浓度并作为培养后终值(B)。
土壤矿化指标计算公式采用下面公式进行计算[19]
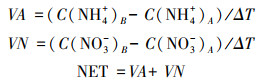
|
式中, VA、VN和NET分别表示土壤净硝化速率、土壤净氨化速率和土壤净氮矿化速率, 单位均为mg kg-1 d-1;ΔT表示为培养时间, 单位为d;C(NH4+)A和C(NO3-)A分别表示培养前土壤中铵态氮和硝态氮含量, C(NH4+)B和C(NO3-)B分别表示培养后土壤中铵态氮和硝态氮含量, 单位均为mg/kg。
1.4 数据分析采用SPSS 18.0软件对相关数据进行单因素(one-way ANOVA)方差分析, 采用Duncan法进行多重比较, 统计显著水平为P < 0.05。图表中所有数据均表示为平均值±标准误(Mean ± SE)。Canoco 5.0统计软件土壤氮矿化数据进行趋势对应分析(DCA)计算出第一排序轴的梯度范围(lengths of gradient), 根据其lengths of gradient的数值小于3.0的基础上, 采用冗余分析法(redundancy analysis, RDA), 对土壤氮矿化数据、土壤理化指标以及植物因子进行排序分析。
2 结果与分析 2.1 高寒草甸退化对植物和土壤理化特征的影响由表 1可知, 对于高寒草甸植物而言, 随退化程度加剧, 植物盖度由96%降低至37.33%, 且四个退化程度间均具有显著性差异(P < 0.05), 植物高度从未退化草地的19.06 cm降低至2.06 cm, 并且4个退化梯度间均具有显著性差异(P < 0.05), 同时, 物种数由15种下降至8种, 且ND、LD和SD间差异显著(P < 0.05)。通过对比发现, 地上生物量由403 g/m2降低至99.31 g/m2、地下生物量由1569.63 g/m2降低至266.35 g/m2, 表明随草地退化程度加剧, 不同退化程度间地上生物量和地下生物量均具有显著性差异(P < 0.05)。
对高寒草甸土壤特征等相关因子的研究发现(表 1), 随高寒草甸退化程度加剧, SOC含量呈逐渐降低趋势, 且四个退化梯度间均具有显著性差异(P < 0.05), TN含量和SM的变化趋势与SOC的变化趋势一致, 且不同退化梯度间均具有显著性差异(P < 0.05), 土壤pH值的变化整体呈下降趋势, 其中ND和LD退化程度间差异不显著(P>0.05), 但与其他两个退化梯度(MD和SD)间均具有显著性差异(P < 0.05)。
2.2 高寒草甸退化对土壤微生物属性的影响高寒草甸退化对MBC、MBN、MBC/MBN、SBR、qCO2和CMBC /CSOC有一定的影响, 并且表现出一定的规律性。研究发现, 随退化程度加剧, MBC、MBN和SBR呈逐渐降低趋势(图 1), 其中MBC的变化从486.32 mg/kg下降到237.77 mg/kg, MBN的变化从34.72 mg/kg下降到19.11 mg/kg, SBR速率由53.5 mg kg-1 d-1下降至24.5 mg kg-1 d-1, 且4个退化程度间均具有显著性差异(P < 0.05)。

|
| 图 1 高寒草甸不同退化程度土壤微生物属性的变化 Fig. 1 Variation of soil microbial properties of alpine meadow under different degradation degrees 小写字母表示不同退化程度间差异性显著(P < 0.05),ND:未退化Non-degraded grassland;LD:轻度退化Light degraded grassland;MD:中度退化Moderate degraded grassland;SD:极度退化Severely degraded grassland |
由图 1所示, qCO2的变化范围为0.11—0.32(d-1)。研究发现, 随着退化程度的加剧, qCO2由未退化草地到重度退化草地表现为降低的趋势, 但差异不显著(P>0.05);而SD与ND、LD、和MD相比, qCO2最高且差异性最显著(P < 0.05)。由图 1所示, CMBC /CSOC的变化范围为0.051—0.038(%), 整体表现为随退化程度的加剧呈逐渐降低趋势, 其中ND、LD、和MD无明显差异(P>0.05), 但与SD相比差异性显著(P < 0.05)。由图 1所示, 微生物生物量碳氮比(MBC/MBN)的变化范围从14.74下降到12.41, 整体表现为随退化程度加剧呈先上升后下降趋势, 其中ND、LD和MD三个退化草地间差异不显著(P>0.05), 但与SD退化草甸间差异显著(P < 0.05)。
2.3 高寒草甸退化对土壤净氮矿化, 硝化和氨化的速率的影响由图 2所示, 随退化程度加剧, 高寒草甸土壤净氨化速率整体呈下降趋势, 其中ND净氨化速率最高为8.64 mg kg-1 d-1, SD净氨化速率最低为1.51 mg kg-1 d-1, 且四个退化程度草地间均具有显著性差异(P < 0.05)。由图 2所示, 随退化程度加剧, 高寒草甸土壤净硝化速率整体呈上升趋势, 其中ND净硝化速率最低, 为5.31 mg kg-1 d-1, SD净硝化速率最高, 为10.71 mg kg-1 d-1, 且四个退化程度草地间均具有显著性差异(P < 0.05)。由图 2可知, 随退化程度加剧, 高寒草甸土壤净氮矿化速率整体呈下降趋势, 其中SD净氮矿化速率最低, 为9.49 mg kg-1 d-1, ND净氮矿化速率最高, 为14.9 mg kg-1 d-1, 且四个退化程度草地间均具有显著性差异(P < 0.05)。

|
| 图 2 高寒草甸退化对土壤净氨化速率、净硝化速率和净氮矿化速率的影响 Fig. 2 Effects of alpine meadow degradation on soil ammoniation rate, nitrification rate and net N mineralization rate |
高寒草甸的退化与土壤氮矿化相关酶(土壤蛋白酶、脲酶、β-乙酰葡糖胺糖苷酶和亮氨酸氨基肽酶)活性关系密切, 由图 3所示, 土壤蛋白酶、UE和LAP的活性随草地退化程度的加剧呈逐渐降低的趋势, 其活性变化范围分别为:土壤蛋白酶活性由0.62 mg g-1 d-1下降到0.34 mg g-1 d-1;UE由0.91 mg g-1 d-1下降到0.55 mg g-1 d-1;LAP由27.07 μmol g-1 d-1下降至22.11 μmol g-1 d-1。通过方差分析可知, 土壤蛋白酶和UE在4个退化程度间均具有显著性差异(P < 0.05), 而LAP的活性在ND和LD间差异不显著(P>0.05), 与MD和SD相比均具有显著性差异(P < 0.05)。

|
| 图 3 高寒草甸不同退化程度土壤酶活性的变化特征 Fig. 3 Changes of soil enzyme activity of alpine meadow under different degradation degrees |
由图 3所示, 土壤中NAG的活性从ND到MD呈逐渐降低趋势, 其活性变化从1.13 μmol g-1 d-1降至0.71 μmol g-1 d-1, 且差异性显著(P < 0.05), 但SD中NAG的活性则高于其他3个退化程度的草地, 与ND相比差异不显著(P>0.05), 但与LD和SD相比差异性显著(P < 0.05)。
2.5 不同退化程度高寒草甸土壤氮矿化与土壤性质和植物因子间的关系分析基于CANOCO 5.0软件分析, 将不同退化程度高寒草甸土壤氮矿化速率进行趋势对应分析(DCA), 并对不同氮矿化速率进行分组和排序, 计算出第一轴的梯度范围(lengths of gradient)是0.78, 这表明氮矿化和土壤性质之间适应于多元统计RDA分析。通过Pearson相关系数分析结合RDA分析了土壤氮矿化与土壤理化性质、酶活性以及植物因子之间的相互关系(图 4)。RDA分析表明:第一、二排序轴特征值分别为0.955、0.035, 同时第一、二排序轴累计解释率分别为95.84%、3.25%, 因此说明第一、二排序轴能够很好地反映植物矿化速率与土壤理化性质和植物因子之间的关系, 且主要由第一排序轴决定。通过RDA分析和Pearson相关分析发现, 高寒草甸退化过程中土壤净氨化速率与土壤蛋白酶、脲酶、亮氨酸氨基肽酶、土壤基础呼吸、土壤微生物生物量碳、土壤微生物生物量氮、植物高度、植被覆盖度以及地下生物量间呈极显著正相关(相关系数分别为R=0.938、R=0.958、R=0.922、R=0.980、R=0.966、R=0.946、R=0.958、R=0.978、R=0.962);土壤蛋白酶、脲酶、亮氨酸氨基肽酶、植物高度、植被覆盖度、地上生物量、微生物生物量碳、微生物生物量氮、微生物代谢熵和土壤基础呼吸与土壤净氮矿化速率之间呈极显著正相关(相关系数分别为R=0.938、R=0.933、R=0.928、R=0.947、R=0.976、R=0.953、R=0.948、R=0.933、R=0.724和R=0.961);土壤净硝化速率与土壤蛋白酶、脲酶、亮氨酸氨基肽酶、植物高度、植被覆盖度、地上生物量、微生物生物量碳、微生物生物量氮、微生物代谢熵、土壤基础呼吸之间呈极显著负相关(相关系数分别为R=-0.906、R=-0.951、R=-0.885、R=-0.936、R=-0.947、R=-0.937、R=-0.951、R=-0.926、R=-0.756和R=-0.967)。

|
| 图 4 土壤氮矿化与土壤环境因子的RDA分析 Fig. 4 Redundancy analysis for soil nitrogen mineralization and soil environmental factors PRO:土壤蛋白酶Protease;UE:土壤脲酶Urease;LAP:亮氨酸氨基肽酶Leucine aminopeptidase;NAG:β-乙酰葡糖胺糖苷酶β-1, 4-N-acetylglucosaminidase;MBC:土壤微生物生物量碳Microbial biomass C;MBN:土壤微生物生物量碳Microbial biomass N;MBC:N:土壤微生物生物量碳氮比Microbial biomass C/N;MBC:SOC:微生物熵Microbial quotient;SBR:土壤基础呼吸Soil basal respiration;qCO2:代谢熵Metabolic quotient;VA:净氨化速率Amination rate;VN:净消化速率Nitrification rate;NET:净氮矿化速率Net N mineralization rate;Height:植被高度;Coverage:植被盖度;Aboveground biomass:地上生物量;Underground biomass:地下生物量 |
高寒草地退化过程在排序轴中也能得到体现, 沿第一轴依次为ND、LD、MD以及SD, 其中, LD分布在土壤养分含量丰富的第一轴左侧, ND分布在土壤呼吸和净氨化速率较大的第一轴左下角, MD靠近原点且沿第一轴分布, SD分布在净消化速率和净氮矿化速率较为严重的第一排序轴右下角, 表明在祁连山高寒草甸区, 草地具有退化趋势, 并且不同的退化草地对土壤养分的需求也各不相同, MD可能成为草地退化的起始阶段或者最终阶段, LD主要作为中间的过渡阶段, 受到土壤理化性质的影响较为显著。
3 讨论 3.1 高寒草甸的退化对微生物特性和酶活性的影响MBC和MBN是土壤有效养分的重要来源, 也是衡量微生物活性的重要指示性因子[20]。有研究指出, 当草地发生退化时引起了土壤养分和MBC和MBN的降低, 并导致土壤肥力下降以及微生物活性的降低, 从而使土壤氮矿化发生变化[21]。作为衡量土壤微生物活性总指标的土壤基础呼吸, 反映了土壤微生物活性和土壤物质代谢的强度[22]。有研究指出, 草地发生退化时土壤呼吸逐渐降低, 这说明草地退化导致土壤质地发生变化, 从而抑制了土壤微生物的活性[23], 同样Anderson等[24]研究指出活跃的微生物群落均有助于土壤基础呼吸。本研究结果显示, 高寒草甸区, 草地退化降低了土壤中MBC、MBN的含量和土壤的呼吸速率, 这与王学霞等[25]、赵玉皓等[26]的结论具有相似性。因此, 草地退化使土壤微生物的活性降低, 从而导致土壤净氮矿化速率呈下降趋势。qCO2通常被解释为微生物的应激指数, 可用于评估土壤微生物群落在底物利用方面的效率[27]。在本研究中观察到随退化程度加剧, SD中qCO2的值显著高于其他3个样地, 这说明低qCO2值可能表明在使用有机碳进行生物合成时土壤微生物量具有较高的效率, 这进一步证明Behera等[28]的观点, 微生物生物量的更高效率表明土壤中有较多的可利用有机碳的参与和能够通过呼吸减少碳素的损失, 从而降低qCO2。CMBC/CSOC表示MBC相对于SOC的积累潜力的指数, 能够用于评估草地退化引起的变化[29]。本研究中发现, 随退化程度加剧CMBC/CSOC表现为降低趋势, 其中SD中CMBC/CSOC显著低于其他3个样地, 这就表明ND、LD和SD中SOC转化为MBC的效率更高。因此, SD中较低的CMBC/CSOC比率意味着该样地中微生物群落在SOC转化为微生物量方面效率较低, 或者样地中某些不可被利用的碳物质产生[30]。
本研究发现, 随高寒草甸退化程度的加剧, 土壤中与氮矿化有关的酶均表现出一定的规律性, 并且主要受到土壤微生物活性的影响。土壤在氮矿化过程中, 土壤微生物则通过调控相关氮素转化的关键酶, 从而控制土壤中氮素的矿化作用[31]。本研究发现, 高寒草地在退化过程中对氨化速率和硝化速率具有较深的影响, 因此, 导致氨化速率下降和硝化速率上升的因素可能与土壤中氮素分解的酶有一定关系。而土壤中蛋白酶、脲酶、NAG和LAP等酶的活性都与氮素转化有直接关系, 能够作为土壤氮矿化的关键酶。本研究发现, 随退化程度加剧, 土壤酶的活性呈逐渐降低趋势, 这与王学霞等[25]的研究有相似的结论。说明在高寒草甸退化过程中, 微生物生物量的降低必然会导致土壤酶活性的下降, 从而使土壤氮矿化速率降低。而氮矿化过程中氨化速率又起到了决定性作用, 这同样可能与这四种酶有关系。同时, 有研究发现, 土壤中可利用有机氮的释放是在这些酶的催化下进行的[32], 因此, 这些酶的活性强弱又是土壤氮循环的限速步骤, 而Chen等[33]研究发现酶对有机氮的降解是氮转化过程中的限制环节, 这与本研究结论相似。
3.2 高寒草甸退化对土壤中N供应能力的影响本研究结果发现, 高寒草甸退化过程中, 氨化速率是净氮矿化速率的主要成分, 这说明高寒草甸退化过程中NH4+是高寒草甸中无机氮的主要形式。与SD比, LD的无机氮可利用性最高, 这应该是退化不严重的样地(ND、LD、MD)土壤肥力较高的原因, 这与Shi[34]的观点具有相似的结论, 土壤有机质的质量和数量被认为是N矿化潜力的主要决定因素。通过对不同退化程度高寒草甸土壤培养后发现, 土壤硝化速率和氨化速率与土壤微生物的活性有一定关系。本研究发现, SD样地氨化速率表现为负值, 说明NH4+的耗损高于产出, 这进一步指出大量的微生物参与了N的固定, 并且Vitousek和Melillo[35]的研究也指出, 土壤微生物固定NH4+的速率与所在环境的干扰有直接关系。在本研究中, SD样地出现负氨化率符合这一发现, 因此极度退化的高寒草地中负氨化速率表明微生物固定NH4+的重要性, 同时也能够揭示了不稳定碳化合物的有限可利用性。Azam等[36]指出NH4+是土壤中微生物同化作用首选N形式, 本研究也同样表明, 虽然SD样地中NO3-转化率较高, 但净氨化速率则表现为负速率, 这说明土壤微生物在同化时可能受到了底物的限制作用, 因此草地在退化过程中NH4+-N的微生物同化作用优先级高于NO3--N的微生物固定作用,
有研究发现当微生物固定含氮量较低的土壤时, 可能会导致NO3--N受到损失[37], 因此, 本研究的一个重要发现是土壤微生物NO3--N固定化, 表明微生物在控制NO3--N浓度中的重要作用。一般而言, 土壤有机碳较低时将会限制净氮矿化的速率, 本研究表明, 随退化程度加剧, 土壤中SOC含量降低, 净氮矿化速率也表现为降低的趋势, 这与Hart等[38]研究, 土壤中氮的矿化速率与可利用有机碳的含量呈线性正相关结果具有一致性。可见, 高寒草甸不同退化程度的净氮矿化速率表明了微生物的活性和可利用有机碳的数量。
3.3 退化草地土壤氮矿化与土壤因子和植物因子间的关系本研究通过RDA分析和Pearson相关系数相结合的方法, 共同分析了土壤氮矿化速率与植物因子以及土壤微生物属性和土壤酶活性等土壤环境因子间的相关性, 研究结果表明, 土壤氮矿化中, 氨化速率和净氮矿化速率主要受到植物因子和土壤因子间(植物高度、植被盖度、地上生物量、PRO、UE、LAP、SBR、MBC和MBN)的影响, 并表现出极显著正相关。这说明高寒草甸退化过程氮矿化为微生物同化提供了可利用的氮, 以及支持这一过程的微生物活性和微生物生物量, 特别是在退化不严重的草甸土壤中。这种关系通常也表明, 草地退化中土壤净氮矿化和微生物的固定化可以通过碳矿化来估算。本研究发现高寒草甸退化时, 土壤氨化速率高于硝化速率, 这说明高寒草甸土壤氮矿化主要为氨化作用, 而通过多元分析发现, 氨化速率与净氮矿化速率之间的密切关联, 因此, 进一步表明NH4+-N的产生高度依赖于N矿化, 反映了有机氮在不同程度高寒草甸退化下对氮矿化的响应[34]。同时, 本研究发现, 草地退化过程中硝化速率呈逐渐升高的趋势, 这可能是因为高寒草甸退化时, 土壤氨化作用高于土壤硝化速率, 从而表现出净硝化速率的下降[39], 同时通过相关分析可知, 土壤硝化速率与氨化速率间呈显著负相关, 这就在一定程度上解释了净硝化速率下降的原因。
4 结论(1) 祁连山东缘高寒草甸退化过程中, 土壤净氮矿化率和氨化速率呈下降趋势, 硝化速率呈上升趋势, 并且该区域土壤氨化作用强烈, 导致该地区土壤无机氮主要以铵态氮为主。
(2) 土壤净氮矿化率和氨化速率作为相对有效性的指标显示了高寒草甸退化样地之间的明显差异, 特别是在未退化草甸和极度退化草甸的土壤中, 这说明微生物活性的重要作用, 并且突出了无机氮生产过程中发生微生物氮固定的重要性。
(3) 高寒草甸在退化过程中植物群落的结构遭受破坏, 导致土壤理化性质发生改变, 使土壤中相关氮素转化的关键酶活性受到抑制, 从而导致高寒草甸土壤氮矿化逐渐降低。
| [1] |
沈海花, 朱言坤, 赵霞, 耿晓庆, 高树琴, 方精云. 中国草地资源的现状分析. 科学通报, 2016, 61(2): 139-154. |
| [2] |
Gos P, Loucougaray G, Colace M P, Arnoldi C, Gaucherand S, Dumazel D, Girard L, Delorme S, Lavorel S. Relative contribution of soil, management and traits to co-variations of multiple ecosystem properties in grasslands. Oecologia, 2016, 180(4): 1001-1013. DOI:10.1007/s00442-016-3551-3 |
| [3] |
Irisarri J G N, Derner J D, Porensky L M, Augustine D J, Reeves J L, Mueller K E. Grazing intensity differentially regulates ANPP response to precipitation in North American semiarid grasslands. Ecological Applications, 2016, 26(5): 1370-1380. DOI:10.1890/15-1332 |
| [4] |
Hong J T, Wang X D, Wu J B. Effects of soil fertility on the N:P stoichiometry of herbaceous plants on a nutrient-limited alpine steppe on the northern Tibetan Plateau. Plant and Soil, 2015, 391(1/2): 179-194. |
| [5] |
Powlson D S, Prookes P C, Christensen B T. Measurement of soil microbial biomass provides an early indication of changes in total soil organic matter due to straw incorporation. Soil Biology and Biochemistry, 1987, 19(2): 159-164. DOI:10.1016/0038-0717(87)90076-9 |
| [6] |
Wang Q R, Li Y C, Klassen W. Changes of soil microbial biomass carbon and nitrogen with cover crops and irrigation in a tomato field. Journal of Plant Nutrition, 2007, 30(4): 623-639. |
| [7] |
Jin H M, Sun O J, Liu J F. Changes in soil microbial biomass and community structure with addition of contrasting types of plant litter in a semiarid grassland ecosystem. Journal of Plant Ecology, 2010, 3(3): 209-217. |
| [8] |
Kasel S, Bennett L T. Land-use history, forest conversion, and soil organic carbon in pine plantations and native forests of south eastern Australia. Geoderma, 2007, 137(3/4): 401-413. |
| [9] |
Bossio D A, Girvan M S, Verchot L, Bullimore J, Borelli T, Albrecht A, Scow K M, Ball A S, Pretty J N, Osborn A M. Soil microbial community response to land use change in an agricultural landscape of western Kenya. Microbial Ecology, 2005, 49(1): 50-62. |
| [10] |
刘建国, 刘卫国. 微生物介导的氮循环过程研究进展. 草地学报, 2018, 26(2): 277-283. |
| [11] |
Owen J S, Wang M K, Wang C H, King H B, Sun H L. Net N mineralization and nitrification rates in a forested ecosystem in northeastern Taiwan. Forest Ecology and Management, 2003, 176(1/3): 519-530. |
| [12] |
Muruganandam S, Israel D W, Robarge W P. Nitrogen transformations and microbial communities in soil aggregates from three tillage systems. Soil Science Society of America, 2010, 74(1): 120-129. |
| [13] |
Campos A C, Etchevers J B, Oleschko K L, Hidalgo C M. Soil microbial biomass and nitrogen mineralization rates along an altitudinal gradient on the cofre de perote volcano (Mexico):the importance of landscape position and land use. Land Degradation & Development, 2014, 25(6): 581-593. |
| [14] |
张金屯. 数量生态学(第二版). 北京: 科学出版社, 2011.
|
| [15] |
全国农业技术推广服务中心. 土壤分析技术规范(第二版). 北京: 中国农业出版社, 2006.
|
| [16] |
鲍士旦. 土壤农化分析(第三版). 北京: 中国农业出版社, 2000.
|
| [17] |
王笛, 逄焕成, 李达, 陈金强, 辛晓平, 徐丽君, 唐雪娟, 郭明英, 朱树声. 苜蓿栽培草地微生物C、N及呼吸特性对不同播种量的响应. 草业学报, 2018, 27(3): 135-143. |
| [18] |
Jenkinson D S, Powlson D S. The effects of biocidal treatments on metabolism in soil-I. Fumigation with chloroform. Soil Biology and Biochemistry, 1976, 8(3): 167-177. DOI:10.1016/0038-0717(76)90001-8 |
| [19] |
Trap J, Bureau F, Akpa-Vinceslas M, Decaens T, Aubert M. Changes in humus forms and soil N pathways along a 130-year-old pure beech forest chronosequence. Annals of Forest Science, 2011, 68(3): 595-606. DOI:10.1007/s13595-011-0063-5 |
| [20] |
Wei Y, Yu L F, Zhang J C, Yu Y C, Deangelis D L. Relationship between vegetation restoration and soil microbial characteristics in degraded karst regions:a case study. Pedosphere, 2011, 21(1): 132-138. DOI:10.1016/S1002-0160(10)60088-4 |
| [21] |
杨怡, 欧阳运东, 陈浩, 肖孔操, 李德军. 西南喀斯特区植被恢复对土壤氮素转化通路的影响. 环境科学, 2018, 39(6): 2845-2852. |
| [22] |
张雪, 梅莉, 宋利豪, 刘力诚, 赵泽尧. 模拟氮沉降对马尾松土壤微生物群落结构及温室气体释放的影响. 生态学报, 2019, 39(6): 1917-1925. |
| [23] |
陈璟, 杨宁. 衡阳紫色土丘陵坡地不同恢复阶段土壤基础呼吸及代谢熵的变化. 热带亚热带植物学报, 2013, 21(6): 514-520. DOI:10.3969/j.issn.1005-3395.2013.06.004 |
| [24] |
Anderson T H, Domsch K H. Application of eco-physiological quotients (qCO2 and qD) on microbial biomasses from soils of different cropping histories. Soil Biology and Biochemistry, 1990, 22(2): 251-255. DOI:10.1016/0038-0717(90)90094-G |
| [25] |
王学霞, 董世魁, 高清竹, 张勇, 胡国铮, 罗文蓉. 青藏高原退化高寒草地土壤氮矿化特征以及影响因素研究. 草业学报, 2018, 27(6): 1-9. |
| [26] |
赵玉皓, 张艳杰, 严月, 刘玉槐, 徐燕, 刘苑秋, 郭晓敏, 娄翼来, 鲁顺保. 亚热带退化红壤区森林恢复类型土壤有机碳矿化对温度的响应. 生态学报, 2018, 38(14): 5056-5066. |
| [27] |
Dilly O, Munch J C. Ratios between estimates of microbial biomass content and microbial activity in soils. Biology and Fertility of Soils, 1998, 27(4): 374-379. DOI:10.1007/s003740050446 |
| [28] |
Behera N, Sahani U. Soil microbial biomass and activity in response to Eucalyptus plantation and natural regeneration on tropical soil. Forest Ecology and Management, 2003, 174(1/3): 1-11. |
| [29] |
Sparling G P. Ratio of microbial biomass carbon to soil organic carbon as a sensitive indicator of changes in soil organic matter. Australian Journal of Soil Research, 1992, 30(2): 195-207. DOI:10.1071/SR9920195 |
| [30] |
McCulley R L, Archer S R, Boutton T W, Hons F M, Zuberer D A. Soil respiration and nutrient cycling in wooded communities developing in grassland. Ecology, 2004, 85(10): 2804-2817. |
| [31] |
Cusack D F, Silver W L, Torn M S, Burton S D, Firestone M K. Changes in microbial community characteristics and soil organic matter with nitrogen additions in two tropical forests. Ecology, 2011, 92(3): 621-632. |
| [32] |
索沛蘅, 杜大俊, 王玉哲, 胡亚林, 刘先. 杉木连栽对土壤氮含量和氮转化酶活性的影响. 森林与环境学报, 2019, 39(2): 113-119. |
| [33] |
Chen H, Li D J, Zhao J, Xiao K C, Wang K L. Effects of nitrogen addition on activities of soil nitrogen acquisition enzymes:a meta-analysis. Agriculture, ecosystems & environment, 2018, 252: 126-131. |
| [34] |
Shi W, Yao H Y, Bowman D. Soil microbial biomass, activity and nitrogen transformations in a turfgrass chronosequence. Soil Biology and Biochemistry, 2006, 38(2): 311-319. |
| [35] |
Vitousek P M, Melillo J M. Nitrate losses from disturbed forests:patterns and mechanisms. Forest Science, 1979, 25(4): 605-619. |
| [36] |
Azam F, Simmons F W, Mulvaney R L. Immobilization of ammonium and nitrate and their interaction with native N in three Illinois Mollisols. Biology and fertility of Soils, 1993, 15(1): 50-54. DOI:10.1007/BF00336288 |
| [37] |
Sotta E D, Corre M D, Veldkamp E. Differing N status and N retention processes of soils under old-growth lowland forest in Eastern Amazonia, Caxiuanã, Brazil. Soil Biology and Biochemistry, 2008, 40(3): 740-750. DOI:10.1016/j.soilbio.2007.10.009 |
| [38] |
Hart S C, Binkley D, Perry D A. Influence of red alder on soil nitrogen transformations in two conifer forests of contrasting productivity. Soil Biology and Biochemistry, 1997, 29(7): 1111-1123. DOI:10.1016/S0038-0717(97)00004-7 |
| [39] |
刘欣, 黄运湘, 袁红, 潘复静, 何寻阳, 张伟, 王克林. 植被类型与坡位对喀斯特土壤氮转化速率的影响. 生态学报, 2016, 36(9): 2578-2587. |