 2020, Vol. 40
2020, Vol. 40文章信息
- 邓秀秀, 施征, 肖文发, 曾立雄, 雷蕾
- DENG Xiuxiu, SHI Zheng, XIAO Wenfa, ZENG Lixiong, LEI Lei
- 干旱和遮荫对马尾松幼苗生长和光合特性的影响
- Effects of drought and shading on growth and photosynthetic characteristics of Pinus massoniana seedlings
- 生态学报. 2020, 40(8): 2735-2742
- Acta Ecologica Sinica. 2020, 40(8): 2735-2742
- http://dx.doi.org/10.5846/stxb201901310223
-
文章历史
- 收稿日期: 2019-01-31
- 网络出版日期: 2019-12-31





2. 南京林业大学南方现代林业协同创新中心, 南京 210037
2. Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing 210037, China
水分和光照是限制植物生长的主要环境因子。当水分和光照发生变化时, 会导致植物形态结构和光合特性的改变。干旱胁迫使叶片气孔关闭或叶肉细胞损伤、光合酶活性降低, 引起植物的光合速率降低[1], 同时破坏叶绿体结构, 使叶绿素含量降低[2], 影响对光能的吸收和利用。在弱光环境中, 植物对光能的捕获受到限制, 制约了植物的光合能力, 引起光合速率的下降, 但会增加光合色素含量[3], 以增大对光能的捕获能力。而干旱与遮荫的交互作用对植物生长的影响有四种假说:“权衡理论”[4], 认为在遮荫环境下, 植物倾向于向地上部分分配更多的生物量, 而减小向根中分配, 影响水分的吸收, 加重干旱对植物生长的影响;“相互作用理论”[5], 认为中度遮荫能缓解干旱的负面影响, 而高光照和过度遮荫则会加重干旱的影响;“促进理论”[6], 认为遮荫可以改善干旱影响下的微环境, 从而缓解植物受干旱的影响;“独立影响理论”[7], 认为二者的交互作用不存在影响, 相互独立。
马尾松(Pinus massoniana)广泛分布于我国亚热带东部地区, 不仅是造林的主要先锋树种, 还是我国主要的产脂树种[8], 发挥着重要的生态和经济作用, 具有喜光、耐干旱、耐贫瘠的生长特性。受光照环境异质性和降水分布不均的影响, 马尾松的生长难免受干旱和弱光环境的胁迫, 且相关研究表明, 干旱会显著抑制马尾松的生长[9-10], 在遮荫环境中马尾松的适应能力较差[11], 但尚不清楚马尾松的光合生理特征对干旱和遮荫环境的响应情况, 且遮荫能否缓解干旱对马尾松植株生长的影响也还未见报道。因此, 本研究以2年生马尾松幼苗为对象, 通过模拟遮荫和干旱环境, 测定马尾松植株的生长指标、光合生理指标和光合色素含量, 研究其生长和光合特性在干旱和遮荫环境下的变化特征, 并验证遮荫和干旱的交互作用对马尾松生长的影响符合哪种假说, 以揭示马尾松植株对干旱和遮荫环境的响应规律和适应机制, 为林木造林管理提供理论依据。
1 材料与方法 1.1 试验材料试验材料为2年生的马尾松容器苗, 来自浙江省龙泉市林业科学研究院苗圃。2018年3月中旬将300株2年生的马尾松幼苗移栽入花盆(直径32 cm, 高22 cm)中, 每盆1株, 种植土壤为马尾松林下土。置于湖北秭归三峡库区森林生态系统国家定位观测研究站(110°54′E, 30°53′N, 海拔296 m), 进行相同光照和水分条件管理。
1.2 试验设计2018年7月1日, 选择120株生长一致(平均株高为65.52 cm, 平均基径为11.61 mm)的马尾松幼苗放入透明遮雨棚中, 进行光照和水分处理, 分别为对照(土壤水分为70%饱和含水量和全光照, CK), 干旱(土壤水分为30%饱和含水量和全光照, DR), 遮荫(土壤水分为70%饱和含水量和光照强度为全光照的30%, LL), 干旱和遮荫交互处理(土壤水分为30%饱和含水量和光照强度为全光照的30%, DRLL)。共4个处理, 每个处理30个重复。遮荫处理采用市场上销售的遮荫网进行遮荫处理, 干旱通过整盆称重法控制, 每天18:00补充当天损失的水分。
1.3 生长指标的测量分别于处理前和处理2个月后用游标卡尺和钢尺测定试验植株的基径和株高, 并计算基径和株高的增长量:处理后的基径(株高)-处理前的基径(株高)。采集植株的当年生针叶, 首先, 用钢尺测量一部分针叶的长度, 每株取平均值作为1次重复。然后, 将一部分针叶利用扫描仪获取针叶的图像, 经过PS软件识别, 计算针叶的叶面积, 将针叶放入65℃烘箱中, 烘干至恒重之后称量干物质量, 比叶面积的计算为叶面积/干物质量。一部分当年生针叶在采完后立即称重(记为鲜叶重), 放入烘箱中, 65℃烘干至恒重后称重(记为干叶重), 针叶含水量的计算为(鲜叶重-干叶重)/鲜叶重×100%[12]。每个处理选4株, 作为4个重复。
1.4 叶片光合参数的测定用便携式光合测定仪Li-6400测定叶片气体交换参数, 测量时分别设定全光照和全光照30%处理的光照强度为1000 μmol m-2 s-1和300 μmol m-2 s-1。每处理选择4株幼苗, 每株幼苗在茎干上选择2簇生长健康的当年生叶分别连续测3次, 测定净光合速率Pn、胞间CO2浓度Ci、气孔导度Gs和蒸腾速率Tr, 并计算植株的瞬时水分利用效率:WUE=Pn/Tr和气孔限制值Ls=1-Ci/Ca(Ca为空气中的CO2浓度)。
1.5 叶绿素含量的测定选用当年生针叶用于叶绿素含量的测定, 每个处理4个重复。针叶叶绿素a(Chla)、叶绿素b(Chlb)、总叶绿素(Chlt)和类胡萝卜素(Car)含量采用乙醇提取法测定[13]。
1.6 可塑性指数(PI)计算
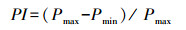
|
式中, Pmax为各参数在所有处理中的最大平均值, Pmin为各参数在所有处理中的最小平均值。
1.7 数据处理采用SPSS 18.0进行数据进行统计分析, 利用LSD进行显著性检验, 用Origin 9.0软件作图。
2 结果与分析 2.1 干旱和遮荫处理对马尾松幼苗生长的影响如图 1所示, 马尾松幼苗基径和株高的增长量在干旱、遮荫和二者的交互处理下均显著减小, 且二者的交互处理加重了干旱和遮荫单一处理下的减小趋势。与对照处理相比, 在干旱、遮荫以及二者的交互处理下基径和株高的增长量分别减少了52.76%、42.77%和84.22%, 28.08%、42.78%和81.36%。

|
| 图 1 干旱和遮荫处理下马尾松幼苗生长量的差异 Fig. 1 The difference of growth increment in P. massoniana seedlings under drought and shading treatment 不同字母表示同一指标在处理之间存在显著差异(P < 0.05) |
马尾松幼苗的针叶特征受干旱和遮荫的影响相对较小,但其在不同处理下的变化趋势存在差异。从图 2可知,与对照相比,针叶长度受干旱影响下虽未发生显著改变,但却减小了9.80%;遮荫处理显著增加了针叶长度,与对照相比,增加了15.96%;二者的交互处理显著减小了针叶长度,减小了14.41%。各处理下,针叶的相对含水量和比叶面积与对照相比均未发生显著改变,但均在干旱和二者的交互处理下呈减小的趋势,在遮荫处理下呈增加的趋势。

|
| 图 2 干旱和遮荫处理下马尾松幼苗当年生针叶形态特征差异 Fig. 2 The difference of morphological features in current-year needles of P. massoniana seedlings under drought and shading treatment |
从表 1可知, 受干旱和遮荫影响下马尾松幼苗的气体交换参数呈现不同程度的影响。与对照相比, 干旱处理下植株的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和细胞间隙CO2浓度(Ci)均显著减小, 分别减小37.55%、62.08%、75.21%和31.09%, 水分利用效率(WUE)和气孔限制值(Ls)显著增大, 分别增大67.96%和123.68%;遮荫处理下植株的Pn、Tr和Gs显著减小, 分别减少42.22%、40.16%和49.11%, Ci、WUE和Ls基本未发生改变;二者的交互处理下各参数的变化趋势同干旱处理较一致, Pn、Tr、Gs和Ci显著减小, 分别减小64.58%、77.39%、85.70%和30.54%, WUE和Ls显著增大, 分别增大60.78%和122.37%。
| 指标Indexes | CK | DR | LL | DRLL |
| 净光合速率(Pn)Net photosyntheic rate/(μmol m-2 s-1) | 14.82±1.03a | 9.26±1.62b | 8.56±0.63b | 5.25±0.78c |
| 气孔导度(Gs)Stomatal conductance/(mmol m-2 s-1) | 0.36±0.05a | 0.09±0.03c | 0.18±0.04b | 0.05±0.02c |
| 细胞间隙CO2浓度(Ci) Intercellular CO2 concentration/(μmol m-2 s-1) |
307.79±11.72a | 212.09±37.67b | 307.43±12.04a | 213.77±39.31b |
| 蒸腾速率(Tr)Transpiration rate/(mmol m-2 s-1) | 3.97±0.32a | 1.51±0.42c | 2.38±0.36b | 0.90±0.26d |
| 水分利用效率(WUE)Water use efficency /(μmol/mmol) | 3.75±0.42b | 6.29±0.87a | 3.63±0.30b | 6.03±1.10a |
| 气孔限制值(Ls)Stomatal limitation/(%) | 0.21±0.03b | 0.47±0.10a | 0.22±0.030b | 0.46±0.10a |
| CK:对照,Control;DR:干旱,Drought;LL:遮荫,Shading;DRLL:干旱遮荫,Drought plus shading; 不同字母表示同一指标在处理之间存在显著差异(P < 0.05) | ||||
从表 2可知, 干旱对马尾松针叶光合色素的影响较小, 与对照相比, 叶绿素a、叶绿素b、类胡萝卜素、总叶绿素、叶绿素a/b以及类胡萝卜素/总叶绿素基本未发生改变。遮荫处理下各光合色素的含量显著增加, 与对照相比, 叶绿素a、叶绿素b、类胡萝卜素和总叶绿素的含量分别增加54.02%、71.10%、38.63%和58.45%, 叶绿素a/b和类胡萝卜素/总叶绿素显著减小, 分别减小10.48%和12.17%。二者的交互处理同遮荫处理一致均增大了各光合色素的含量, 与对照相比, 叶绿素a、叶绿素b、类胡萝卜素和总叶绿素的含量分别增加44.02%、59.48%、41.58%和48.02%, 叶绿素a/b显著减小, 减小了9.63%, 而类胡萝卜素/总叶绿素未发生显著改变。
| 指标Indexes | CK | DR | LL | DRLL |
| 叶绿素a Chla/(mg/g) | 0.76±0.05b | 0.82±0.11b | 1.16±0.12a | 1.09±0.18a |
| 叶绿素b Chlb/(mg/g) | 0.26±0.03b | 0.28±0.04b | 0.45±0.04a | 0.42±0.08a |
| 类胡萝卜素Car/(mg/g) | 0.17±0.01b | 0.17±0.01b | 0.24±0.02a | 0.24±0.03a |
| 总叶绿素Chlt/(mg/g) | 1.02±0.08b | 1.10±0.14b | 1.62±0.17a | 1.51±0.26a |
| 叶绿素a/b Chla/b | 2.87±0.16a | 2.97±0.23a | 2.57±0.05b | 2.59±0.13b |
| 类胡萝卜素/总叶绿素Car/Chlt | 0.17±0.005a | 0.16±0.01ab | 0.15±0.01b | 0.16±0.02ab |
| 不同字母表示同一指标在处理之间存在显著差异(P < 0.05) | ||||
植株性状的可塑性变化与植株适应环境的能力和资源的可利用情况有关。从图 3可知, 整体上看, 马尾松幼苗受光照和水分影响下可塑性能力的差异不大(PI水分总平均=0.26, PI光照总平均=0.26), 但不同性状受光照和水分影响的可塑性能力存在差别。具体表现为, 植株的生长、针叶特征受光照和水分影响下的可塑性能力较一致, 其可塑指标的平均值分别为植株的生长:PI水分=0.40、PI光照=0.43, 针叶特征:PI水分=0.07、PI光照=0.09;光合色素含量受光照影响下的可塑性能力比水分大440%(PI水分=0.05、PI光照=0.27);气体交换参数受光照影响下的可塑性能力比水分小53%(PI水分=0.50、PI光照=0.24)。

|
| 图 3 光照和水分影响下可塑性指数的双变量示图 Fig. 3 The bivariate expression of plasticity index under the influence of light and water |
环境发生变化时, 植物的生长将受到一定程度的影响, 这主要归结于光合产物合成能力的改变。在本研究中, 干旱和遮荫均抑制了马尾松幼苗基径和株高的生长, 这与相关学者的研究结果一致[12, 14], 这是因为在受到光照和水分条件制约的影响下, 植株的净光合速率下降, 光合产物的合成受到影响, 从而限制了植株的增粗和增高生长。而在干旱与遮荫的交互处理下, 马尾松幼苗基径和株高的生长量, 以及净光合速率均受到了更严重的抑制, 说明遮荫未能缓解干旱对植株的负面影响。二者的交互作用对马尾松幼苗生长的影响不符合“权衡理论”、“促进理论”和“独立影响理论”, 更倾向于符合“相互作用理论”。这与Guo等[15]对三角枫(Acer buergerianum)的研究结果一致, 而与刘翠菊等[12]对耐冬山茶(Camellia japonica)研究中, 发现二者交互作用的影响符合“促进理论”的结论不一致, 这种结果的差异可能与不同物种对不同环境因子的响应程度存在差别有关。
环境胁迫对植物生长的影响机制因不同的环境因子而存在差别。通常认为, 在干旱环境中, 植物光合速率下降分别受“气孔限制”[14, 16]和“非气孔限制”[12, 17]的影响。根据Farquhar等[18]的光合控制理论认为, 当气孔导度、细胞间隙CO2浓度降低, 气孔限制值增大时, 净光合速率的降低主要受“气孔限制”的影响;而当细胞间隙CO2浓度升高、气孔限制值降低时, 净光合速率的降低则主要受“非气孔限制”的影响。在本研究中, 干旱处理下马尾松植株Pn的下降伴随着Gs和Ci的下降和Ls的升高, 因此可以认为马尾松幼苗光合速率的下降受到了“气孔限制”的影响, 导致进入叶片的CO2受到阻碍, 减少了植株进行光合作用时所需原料的供应。植株的蒸腾速率显著减小, 水分利用效率显著增加, 这与气孔关闭减少了针叶表面的水分蒸发, 提高了植株对水分的利用效率[19]有关。另外, 马尾松幼苗的针叶长度和比叶面积显著减小, 针叶含水量有所减小, 但变化不显著。说明通过针叶形态特征和气孔限制等的调节减小了马尾松在干旱胁迫下水分损失的机会, 维持了相对较稳定的含水量, 表现了较强的保水能力, 以保证马尾松植株在生长过程和生理活动中所需水分的供应。相关学者表明, 干旱胁迫可使叶绿素的合成受阻或加速叶绿素的降解[20-21], 进而减弱对光能的吸收和传递能力, 引起植株光合功能的改变。而在本研究中, 马尾松幼苗的叶绿素含量、叶绿素a/b和类胡萝卜素/总叶绿素的比值均未在干旱处理下发生显著改变, 表明植株对光能的吸收和利用能力并未因干旱而受到较大影响。
遮荫处理显著减小了马尾松植株的Pn, 同时使Gs和Tr也均显著下降, 说明在受弱光环境的影响下, 马尾松植株的蒸腾速率因气孔导度的闭合而有所下降, 这与魏明月等[22]对烟草(Nicotiana tabacum)以及吕程瑜和刘艳红[23]对梓叶槭(Acer catalpifolium)的研究结果一致。气孔闭合会限制对CO2的吸收, 而马尾松植株的Ci却保持不变, 且遮荫减少了植株的净光合速率, 表明此时对CO2的吸收和同化情况仍保持相对协调。此外, 相关研究表明, 在弱光环境中, 植株可通过增大针叶长度、比叶面积[24]和光合色素含量[25-28]来增大对光能的捕获机会和对光能的吸收和利用能力, 以适应弱光环境。在本研究中, 遮荫处理不仅增大了马尾松植株的针叶长度和比叶面积, 还显著增加了植株的叶绿素a、叶绿素b、总叶绿素和类葫芦卜素含量, 这就保证了在弱光环境中马尾松幼苗能尽可能多的吸收和利用光能进行光合作用, 缓解一部分受光照不足的影响。同时, 遮荫处理使马尾松针叶的叶绿素a/b显著减小, 说明叶绿素b的增长量相对更高, 根据薛思雷等[29]的研究认为, 叶绿素b相对含量的增高可提高对弱光的捕获能力, 由此可见马尾松通过对光合色素含量的调节增强了对弱光环境的适应;类胡萝卜素/总叶绿素的值显著减小, 而类胡萝卜素具有猝灭过剩光能的作用[30], 说明相较于强光环境, 马尾松植株在弱光环境中对过剩光能淬灭能力会有所减弱, 这也是植株对胁迫环境长期适应的体现。
在干旱和遮荫的交互处理下, 马尾松植株的Pn下降伴随着Gs和Ci下降以及Ls的升高, 表明光合速率的下降仍受到了“气孔限制”的影响;且植株的气孔导度显著小于干旱和遮荫的单一处理, 这与Quero等[6]对欧洲栓皮栎(Quercus suber)、冬青栎(Quercus ilex)、非洲栎(Quercus canariensis)和比利牛斯栎(Quercus pyrenaica)以及刘翠菊等[12]对耐冬山茶的研究中, 发现遮荫处理增大了干旱胁迫下叶片的气孔导度, 缓解了一部分水分亏缺影响的研究结果不一致, 说明在本研究中, 遮荫未能缓解干旱胁迫中马尾松幼苗受水分亏缺的影响。植株的针叶长度和比叶面积的变化同干旱处理一致, 均显著减小, 即在弱光、缺水的环境中, 马尾松植株的针叶会优先保证适应干旱, 保证针叶的含水量, 以维持正常的生理活动。但光合色素含量的变化趋势同遮荫处理一致, 均显著增加, 而叶绿素a、叶绿素b和总叶绿素的增长量小于遮荫单一处理, 说明干旱减弱了在遮荫环境中针叶光合色素的合成能力。综上所述, 二者的交互处理不仅未能缓解干旱对马尾松植株的负面影响, 还削弱了对光能的捕获能力和机会。这也解释了植株的净光合速率进一步受到抑制的原因, 证实了干旱和遮荫二者的交互作用对马尾松幼苗生长的影响更符合“相互作用理论”。
4 结论本研究表明, 干旱、遮荫均抑制了马尾松幼苗的生长, 二者的交互作用符合“相互作用理论”。马尾松的针叶形态和光合生理特征在干旱和遮荫环境下呈现不同的响应和适应机制。干旱环境下, 马尾松植株通过减小针叶长度和比叶面积, 减少了水分损失的机会;通过气孔关闭, 减少蒸腾速率, 提高水分利用效率, 使植株净光合速率的下降受到了“气孔限制”的影响。遮荫环境下, 马尾松植株通过增大针叶长度、比叶面积和光合色素含量, 提高对光能的捕获能力和机会, 来适应弱光环境。在二者交互处理下, 马尾松植株的生长和净光合速率均进一步降低;遮荫环境不仅未能缓解受干旱的影响下使针叶长度、比叶面积和气孔导度减小的趋势, 还使光合色素含量的增长量受干旱的影响下有所减小, 从而加重了对马尾松生长的影响。
| [1] |
Lawlor D W, Cornic G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant, Cell & Environment, 2010, 25(2): 275-294. |
| [2] |
Bartels D, Sunkar R. Drought and salt tolerance in plants. Critical Reviews in Plant Sciences, 2005, 24(1): 23-58. |
| [3] |
刘建锋, 杨文娟, 江泽平, 郭泉水, 金江群, 薛亮. 遮荫对濒危植物崖柏光合作用和叶绿素荧光参数的影响. 生态学报, 2011, 31(20): 5999-6004. |
| [4] |
Smith T, Huston M. A theory of the spatial and temporal dynamics of plant communities. Vegetatio, 1989, 83(1/2): 49-69. |
| [5] |
Holmgren M, Scheffer M, Huston A M. The interplay of facilitation and competition in plant communities. Ecology, 1997, 78(7): 1966-1975. |
| [6] |
Quero J L, Villar R, Marañón T, Zamora R. Interactions of drought and shade effects on seedlings of four Quercus species:physiological and structural leaf responses. New Phytologist, 2006, 170(4): 819-834. |
| [7] |
Holmgren M. Combined effects of shade and drought on tulip poplar seedlings:trade-off in tolerance or facilitation?. Oikos, 2000, 90(1): 67-78. |
| [8] |
邓慧华, 洪滔, 吴承祯, 谢安强, 林晗, 洪伟. 马尾松(Pinus massoniana)内生真菌的产油脂效果. 应用与环境生物学报, 2014, 20(3): 377-381. |
| [9] |
喻方圆, 徐锡增, Guy R D. 水分和热胁迫处理对4种针叶树苗木气体交换和水分利用效率的影响. 林业科学, 2004, 40(2): 38-44. |
| [10] |
杜明凤, 丁贵杰, 赵熙州. 不同家系马尾松对持续干旱的响应及抗旱性. 林业科学, 2017, 53(6): 21-29. |
| [11] |
梁春, 林植芳, 孔国辉. 不同光强下生长的亚热带树苗的光合-光响应特性的比较. 应用生态学报, 1997, 8(1): 7-11. |
| [12] |
刘翠菊, 郭霄, 王奎玲, 刘庆超, 孙迎坤, 姜新强, 刘庆华. 耐冬山茶对不同光照和水分的生理生态学响应. 应用生态学报, 2018, 29(4): 1125-1132. |
| [13] |
李合生. 现代植物生理学 (第二版). 北京: 高等教育出版社, 2006.
|
| [14] |
Wu J W, Li J Y, Su Y, He Q, Wang J H, Qiu Q, Ma J W. A morphophysiological analysis of the effects of drought and shade on Catalpa bungei plantlets. Acta Physiologiae Plantarum, 2017, 39(3): 80. |
| [15] |
Guo X, Guo W H, Luo Y J, Tan X F, Du N, Wang R Q. Morphological and biomass characteristic acclimation of trident maple (Acer buergerianum Miq.) in response to light and water stress. Acta Physiologiae Plantarum, 2013, 35(4): 1149-1159. |
| [16] |
Cavatte P C, Oliveira Á A G, Morais L E, Martins S C V, Sanglard L M V P, DaMatta F M. Could shading reduce the negative impacts of drought on coffee? A morphophysiological analysis. Physiologia Plantarum, 2012, 144(2): 111-122. |
| [17] |
León-Sánchez L, Nicolás E, Nortes P A, Maestre F T, Querejeta J I. Photosynthesis and growth reduction with warming are driven by nonstomatal limitations in a Mediterranean semi-arid shrub. Ecology and Evolution, 2016, 6(9): 2725-2738. |
| [18] |
Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology, 1982, 33: 317-345. |
| [19] |
Chaves M M, Flexas J, Pinheiro C. Photosynthesis under drought and salt stress:regulation mechanisms from whole plant to cell. Annals of Botany, 2009, 103(4): 551-560. |
| [20] |
王宇超, 王得祥, 彭少兵, 何帆. 干旱胁迫对木本滨藜生理特性的影响. 林业科学, 2010, 46(1): 61-67. |
| [21] |
姚春娟, 郭圣茂, 马英超, 赖晓莲, 杨肖华. 干旱胁迫对4种决明属植物光合作用和叶绿素荧光特性的影响. 草业科学, 2017, 34(9): 1880-1888. |
| [22] |
魏明月, 云菲, 刘国顺, 宋亮. 不同光环境下烟草光合特性及同化产物的积累与分配机制. 应用生态学报, 2017, 28(1): 159-168. |
| [23] |
吕程瑜, 刘艳红. 不同遮荫条件下梓叶槭幼苗生长与光合特征的种源差异. 应用生态学报, 2018, 29(7): 2307-2314. |
| [24] |
王凯, 朱教君, 于立忠, 孙一荣, 陈光华. 遮阴对黄波罗幼苗的光合特性及光能利用效率的影响. 植物生态学报, 2009, 33(5): 1003-1012. |
| [25] |
Wittmann C, Aschan G, Pfanz H. Leaf and twig photosynthesis of young beech (Fagus sylvatica) and aspen (Populus tremula) trees grown under different light regime. Basic and Applied Ecology, 2001, 2(2): 145-154. |
| [26] |
Zhang Y J, Yan F, Gao H, Xu Y Z, Guo Y Y, Wang E J, Li Y H, Xie Z K. Chlorophyll content, leaf gas exchange and growth of oriental lily as affected by shading. Russian Journal of Plant Physiology, 2015, 62(3): 334-339. |
| [27] |
白宇清, 谢利娟, 王定跃. 不同遮荫、土壤排水处理对毛棉杜鹃幼苗生长及光合特性的影响. 林业科学, 2017, 53(2): 44-53. |
| [28] |
Huang C J, Wei G, Jie Y C, Xu J J, Anjum S A, Tanveer M. Effect of shade on plant traits, gas exchange and chlorophyll content in four ramie cultivars. Photosynthetica, 2016, 54(3): 390-395. |
| [29] |
薛思雷, 王庆成, 孙欣欣, 张命军. 遮荫对水曲柳和蒙古栎光合、生长和生物量分配的影响. 植物研究, 2012, 32(3): 354-359. |
| [30] |
Hormaetxe K, Becerril J M, Fleck I, Pintó M, García-Plazaola J I. Functional role of red (retro)-carotenoids as passive light filters in the leaves of Buxus sempervirens L.:increased protection of photosynthetic tissues?. Journal of Experimental Botany, 2005, 56(420): 2629-2636. |