 2020, Vol. 40
2020, Vol. 40文章信息
- 王银平, 匡箴, 蔺丹清, 李佩杰, 杨彦平, 刘凯
- WANG Yinping, KUANG Zhen, LIN Danqing, LI Peijie, YANG Yanping, LIU Kai
- 长江安庆新洲水域鱼类群落结构及多样性
- Community structure and species diversity of fish around the Xinzhou shoal in the Anqing section of the Yangtze River, China
- 生态学报. 2020, 40(7): 2417-2426
- Acta Ecologica Sinica. 2020, 40(7): 2417-2426
- http://dx.doi.org/10.5846/stxb201901160130
-
文章历史
- 收稿日期: 2019-01-16
- 网络出版日期: 2019-12-26





 (Hemiculter bleekeri Warpachowsky,1887)、银鮈(Squalidus argentatus Sauvage et Dabry,1874)和似鳊(Pseudobrama simoni Bleeker,1864)。4种摄食功能群中,杂食性(42.19%)和肉食性(35.94%)鱼类物种数比例较高;3种生态类群中,淡水定居性鱼类占绝对优势(84.37%);3种栖息水层类型中,底层鱼类物种数比例较高,占37.50%。大型经济鱼类占总渔获物比例低(< 0.01%),但个体较大,因而相对重要性指数(IRI)高。总体上,新洲鱼类群落多样性和丰富度指数较高,均匀度指数偏低,个体小型化趋势明显。捕捞强度过大、水利工程建设导致的江湖阻隔及外来物种入侵是新洲水域渔业资源衰退的主要因素。由此,建议持续开展长江渔业资源监测,加强长江干流沙洲水域渔业资源保护。
(Hemiculter bleekeri Warpachowsky,1887)、银鮈(Squalidus argentatus Sauvage et Dabry,1874)和似鳊(Pseudobrama simoni Bleeker,1864)。4种摄食功能群中,杂食性(42.19%)和肉食性(35.94%)鱼类物种数比例较高;3种生态类群中,淡水定居性鱼类占绝对优势(84.37%);3种栖息水层类型中,底层鱼类物种数比例较高,占37.50%。大型经济鱼类占总渔获物比例低(< 0.01%),但个体较大,因而相对重要性指数(IRI)高。总体上,新洲鱼类群落多样性和丰富度指数较高,均匀度指数偏低,个体小型化趋势明显。捕捞强度过大、水利工程建设导致的江湖阻隔及外来物种入侵是新洲水域渔业资源衰退的主要因素。由此,建议持续开展长江渔业资源监测,加强长江干流沙洲水域渔业资源保护。安庆江段位于长江下游安徽境内, 上起宿松县汇口镇, 下至枞阳县老洲镇, 全长243 km, 约占整个安徽江段总长的2/3。长江安庆江段作为下游典型的感潮河段, 受径流和潮汐的双重影响[1]。在潮水的作用下, 该江段水流平缓, 沙洲众多, 良好的水文环境和丰富的饵料生物使其成为长江下游水生动物优良的栖息和繁育场所[2]。长江安庆江段属分汊型河道, 沿岸有许多湖泊和支流汇入长江, 再加上河道弯曲, 在水力条件的作用下形成了十分多样的栖息地环境, 鱼类种类繁杂, 渔业资源丰富[2-3]。该江段分布有安庆市江豚自然生态保护区、长江刀鲚国家级水产种质资源保护区等重要生态敏感区, 是鱼类觅食、栖息的重要场所[4]。新洲是安庆江段众多沙洲中的一个, 四面滨江, 潮落即洲, 潮涨即江, 常水位水域面积12.24 km2。同时, 新洲水域位于刀鲚保护区的核心区, 其科研价值和环境生态地位不容小觑。
随着经济的快速发展, 人类涉水活动趋于频繁, 长江下游水文情势、水温和泥沙条件等已发生改变[5], 水利枢纽工程建设进一步加速了鱼类等水生生物栖息环境的改变[6]。沙洲水域的鱼类群落将会如何响应有待研究, 因此亟待加强对长江鱼类群落结构及其多样性的系统研究。长江安庆江段鱼类研究始于20世纪90年代初, 有报道共进行过数次详实不等的渔业资源调查, 内容主要集中在鱼类种属记录及渔业生产状况[7], 人类活动对鱼类多样性的影响等方面。近年来, 关于安庆段鱼类群落结构和多样性特征的研究较少, 仅张敏莹等[2]依据1990—2004年长江下游渔业资源动态监测数据, 从群落结构及生物多样性变化等方面对安庆江段鱼类群落进行了描述, 刘明典等[8]对安庆江段春季鱼类群落结构及物种多样性现状进行研究。鱼类群落结构变化是对人为影响及水域环境变化的响应, 其演变过程不仅可能导致渔业功能的退化, 也会使水域生态系统失去自我调控的重要功能, 因此保持特定水域鱼类群落结构的合理性是长江渔业管理与生态系统恢复的关键[9]。本研究采用网簖、多目刺网和地笼等多种网具对长江安庆新洲水域鱼类群落结构和多样性空间分布特征进行调查, 以探究鱼类群落结构特征及渔业资源现状, 同时为安庆江段渔业资源保护和恢复提供科学依据。
1 材料与方法 1.1 采样方法于2017年4月、7月、10月和12月对安庆江段新洲水域鱼类群落进行季节性调查, 设置新洲头(30°29′31.19″N, 117° 9′27.81″E)、新河口(30°29′13.11″N, 117° 8′31.52″E)和鹅毛洲尾(30°31′33.82″N, 117°11′16.96″E)共3个采样区域(图 1)。采用网簖(网长20 m, 网宽10 m, 网目1 cm)进行鱼类样品采集, 地笼(网长10 m, 网目1 cm)和多目刺网(网长150 m, 网目从小到大分别为1.2、2、4 cm)补充鱼类种类调查, 地笼和多目刺网每月中下旬连续采样调查3 d, 网簖和地笼放置24 h后收集渔获物。多目刺网当日晚上放置, 次日清晨起网收集。渔获物收集后立即放置于冰盒上, 带回实验室依据相关文献鉴定到种[10], 利用数显游标卡尺和电子天平测定所有鱼类的体长和体重。体重精确到0.1 g, 体长精确到0.01 mm。

|
| 图 1 新洲水域鱼类采样区域 Fig. 1 Fish sampling transects in waters of Xinzhou |
各采样区域总氮(TN)、总磷(TP)、透明度(SD)和叶绿素a(Chl.a)等水质指标及岸线分布特征测定与鱼类采样同步进行, 具体结果见表 1。
| 采样区域 Sample area |
水深 Depth/m |
透明度 Transparency/m |
总氮 Total nitrogen/(mg/L) |
总磷 Total phosphorus/(mg/L) |
叶绿素a Chlorophyll a/(μg/L) |
高锰酸盐指数 Permanganate index/(mg/L) |
岸线特征 Shoreline feature |
| 新洲 | 28.81 | 0.33 | 2.17 | 0.12 | 3.41 | 2.83 | 石质固化岸线, 洲头为浅型沙滩 |
| 新河口 | 25.93 | 0.29 | 2.19 | 0.12 | 3.41 | 2.69 | 泥质岸线, 浅水区有湿生植物 |
| 鹅毛洲 | 27.51 | 0.30 | 2.16 | 0.12 | 4.02 | 2.74 | 砂质岸线 |
按照鱼类的栖息环境和洄游方式, 新洲水域鱼类存在3种生态类群[11]。参照鱼类食性文献资料和食性类型划分方法, 新洲水域鱼类可划分为肉食性、杂食性、植食性、浮游食性和碎屑食性5类[10]。根据已有文献[4], 将初次性成熟小于2龄, 最大体长小于24 cm的鱼类划为小型鱼类。根据鱼类在垂直空间的分布差异, 将其分为中上层、中下层和底层鱼类[12]。
1.2.2 鱼类优势种鱼类优势种利用相对重要性指数[13](index of relative importance, IRI)进行表征。
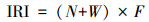
|
式中, IRI为相对重要性指数, N为第i种鱼的尾数占总尾数的百分比, W为第i种鱼的质量占总质量的百分比, F为第i种鱼出现次数占总调查样点的百分比。
将IRI ≥500的物种定为优势种, 100 ≤IRI<500的物种定为常见种, 10≤IRI<100的物种定为一般种, IRI<10的物种定为少见种[14]。
1.2.3 生物多样性指数鱼类物种调查数据采用Shannon-Weiner指数(H)[15], Wilhm改进指数(H′)[16], Pielou均匀度指数(E)[17]和Simpson优势集中度指数(C)[14]进行分析评价。计算公式如下:
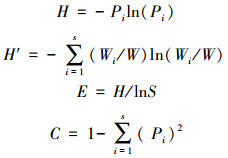
|
式中, Pi为鱼类群落中第i种鱼的个体数占所有鱼类总个体数的比例。W为渔获物总生物量, Wi为第i个物种生物量, S代表所有鱼类种类数。
采用单因素方差法(ANOVA, One-way analysis of variance)对渔获物数量及重量、物种数季节及采样区域间的差异进行检验分析, 若差异显著, 再次进行Tukey多重比较。利用IBM SPSS Statistics 24.0进行数据统计分析。
2 结果 2.1 鱼类种类组成本次调查共采集渔获物135.4 kg, 共计15049尾, 鱼类64种, 隶属6目11科48属(表 2)。鲤科鱼类占优势, 共29属40种, 占总物种数的62.5%;其次为鲿科6种, 占9.4%;鳅科和沙塘鳢科占6.3%, 均为4种;其他7科种类数均较少, 共占总物种数的15.6%。其中, 小型鱼类总计32种, 占总物种数的50.0%, 数量占总量的69.3%。3个采样区域中, 新河口水域鱼类种类最多为52种, 新洲头和鹅毛洲尾均为50种。4个采样季节中, 夏季物种最多为44种, 春季最少为28种。
| 物种 Species |
生态类群 Ecological guilds |
||
| 鲱形目Clupeiformes | |||
| 鳀科Engraulidae | |||
| 刀鲚Coilia nasus* | C | RS | U |
| 鲤形目Cypriniformes | |||
| 鲤科Cyprinidae | |||
| 鲢Hypophthalmichthys molitrix | P | RL | U |
| 鳙Aristichthys nobilis | P | RL | U |
| 鲤Cyprinus carpio | O | SF | D |
| 鲫Carassius auratus | De | SF | L |
| 鳊Parabramis pekinensis | H | SF | L |
| Hemiculter leucisculus* |
P | SF | U |
| 鲂Megalobrama skolkovii | O | SF | L |
| 鳡Elopichthys bambusa | C | RL | U |
| 草鱼Ctenopharyngodon idellus | H | RL | L |
| 青鱼Mylopharyngodon piceus | C | RL | L |
| 花魚骨Hemibarbus maculatus* | C | SF | L |
| 麦瑞加拉鲮Cirrhinus mrigalcarp* | O | SF | D |
| 华鳈Sarcocheilichthys sinensis* | C | SF | L |
| 蛇鮈Saurogobio dabryi* | O | SF | L |
| 似鳊Pseudobrama simoni* | P | SF | L |
| 似鱎Toxabramis swinhonis * | P | SF | U |
| 银鮈Squalidus argentatus* | O | SF | L |
| 银鲴Xenocypris argentea | H | SF | L |
| 飘鱼Pseudolaubuca sinensis* | O | SF | U |
| 麦穗鱼Pseudorasbora parva* | O | SF | U |
| 棒花鱼Abbottina rivularis* | O | SF | D |
| 达氏鲌Culter dabryi | C | SF | L |
| 蒙古鲌Culter monggolicus | C | SF | U |
| 翘嘴鲌Culter alburnus | C | SF | U |
| 贝氏Hemiculter bleekeri* |
P | SF | U |
| 赤眼鳟Squaliobarbus curriculus | O | SF | U |
| 团头鲂Megalobrama amblycephala | H | SF | L |
| 黑鳍鳈Sarcocheilichthys nigripinnis | O | SF | D |
| 黄尾鲴 Xenocypris davidi | H | SF | D |
| 细鳞鲴Xenocypris microlepis | O | SF | L |
| 兴凯鱊Acheilognathus chankaensis* | De | SF | U |
| 斑条鱊Acheilognathus taenianalis* | O | SF | U |
| 宽鳍鱲Zacco platypus | O | SF | U |
| 马口鱼Opsariicjthys bidens | C | SF | U |
| 寡鳞飘鱼Pseudolaubuca engraulis | O | SF | U |
| 高体鳑鲏Rhodeus ocellatus * | De | SF | U |
| 中华鳑鲏Rhodeus sinensis | O | SF | D |
| 点纹银鮈Squalidus wolterstorffi* | O | SF | L |
| 红鳍原鲌Cultrichthys erythropterus | C | SF | U |
| 小口小鳔鮈Microphysogobio microstomus* | O | SF | L |
| 鳅科Cobitidae | |||
| 泥鳅Misgurnus anguillicaudatus* | O | SF | D |
| 中华花鳅Cobitis sinensis* | O | SF | D |
| 紫薄鳅Leptobotia taeniaps* | C | SF | D |
| 花斑副沙鳅Parabotia fasciata* | O | SF | D |
| 斗鱼科Belontiidae | |||
| 圆尾斗鱼Macropodus ocellatus* | O | SF | L |
| 刺鳅科Mastacembelidae | |||
| 中华刺鳅Mastacembelus aculeatus | C | SF | D |
| 鮨科Serranidae | |||
| 鲈形目Perciformes | |||
| 鳜Siniperca chuatsi | C | SF | D |
| 大眼鳜Siniperca kneri | C | SF | D |
| 中国花鲈Lateolabrax maculatus | C | RS | U |
| 沙塘鳢科Odontobutidae | |||
| 乌鳢Channa argus | C | SF | D |
| 沙塘鳢Odontobutis obacurus* | C | SF | D |
| 小黄黝鱼Micropercops swinhonis* | O | SF | L |
| 子陵吻虾虎Rhinogobius giurinus* | C | RS | D |
| 鲇形目Siluriformes | |||
| 鲿科Bagridae | |||
| 黄颡鱼Pseudobagrus fulvidraco* | O | SF | D |
| 长吻鮠Leiocassis longirostris | C | RL | D |
| 瓦氏黄颡鱼Pelteobagrus vachelli | O | SF | D |
| 圆尾拟鲿Pseudobagrus tenuis | O | SF | D |
| 长须黄颡鱼Pelteobagrus eupogon* | O | SF | D |
| 光泽黄颡鱼Pelteobaggrus nitidus* | O | SF | D |
| 鲇科Siluridae | |||
| 鲇Silurus asotus | C | SF | D |
| 大口鲇Silurus meriordinalis | C | SF | D |
| 颌针鱼目Beloniformes | |||
| 鱵科Hemirhamphidae | |||
| 间下鱵Hyporhamphus intermedius* | C | RS | U |
| 胡瓜鱼目Osmeriformes | |||
| 银鱼科Salangidae | |||
| 陈氏短吻银鱼Salangichthys tangkahkeii* | C | SF | U |
| *:小型鱼类Small fish; O:杂食性Omnivore; P:浮游生物食性Planktivore; H:植食性Herbivore; De:碎屑食性Detritivore; C:肉食性Carnivore; U:中上层Upper; L:中下层Lower; D:底层Demersal; RL:河湖洄游性River-lake migratory; SF:淡水定居性Sedentary fish; RS:江海洄游性River-sea migratory | |||
相对重要性指数(IRI)值高于500的鱼类有鳊(Parabramis pekinensis Basilewsky, 1855)、鲤(Cyprinus carpio Linnaeus, 1758)、贝氏
| 物种 Species |
数量百分比N/% Quantity percentage |
重量百分比W/% Weight percentage |
出现频率F/% Frequency |
相对重要性指数IRI Index of relative importance |
| 鳊Parabramis pekinensis | 5.47 | 12.42 | 83.33 | 1491.05 |
| 鲤Cyprinus carpio | 3.34 | 33.29 | 33.33 | 1221.02 |
| 贝氏Hemiculter bleekeri* |
23.22 | 5.00 | 25 | 705.45 |
| 银鮈Squalidus argentatus* | 8.01 | 0.45 | 83.33 | 704.71 |
| 似鳊Pseudobrama simoni* | 7.77 | 1.48 | 58.33 | 539.58 |
| 刀鲚Coilia nasus* | 3.03 | 0.68 | 100 | 371.19 |
| 鲫Carassius auratus | 2.93 | 2.24 | 58.33 | 301.67 |
| 中华花鳅Cobitis sinensis* | 2.46 | 0.067 | 100 | 252.86 |
| 紫薄鳅Leptobotia taeniaps* | 2.85 | 0.52 | 75 | 252.51 |
| 光泽黄颡鱼Pelteobaggrus nitidus* | 3.84 | 1.63 | 41.67 | 227.70 |
| 银鲴Xenocypris argentea | 4.90 | 3.29 | 25 | 204.64 |
| 鲢Hypophthalmichthys molitrix | 1.66 | 20.60 | 8.33 | 185.50 |
| 鲇Silurus asotus | 0.34 | 1.43 | 100 | 176.62 |
| 陈氏短吻银鱼Salangichthys tangkahkeii* | 2.90 | 0.087 | 41.67 | 124.54 |
| 鳙Aristichthys nobilis | 0.16 | 2.31 | 50 | 123.35 |
| 翘嘴鲌Culter alburnus | 1.14 | 0.43 | 75 | 117.75 |
| 子陵吻鰕虎Rhinogobius giurinus* | 2.41 | 0.072 | 41.67 | 103.42 |
| 点纹银鮈Squalidus wolterstorffi* | 1.04 | 0.053 | 75 | 81.71 |
| 红鳍原鲌Cultrichthys erythropterus | 0.47 | 0.28 | 100 | 74.80 |
| 麦瑞加拉鲮Cirrhinus mrigalcarp | 0.34 | 0.38 | 100 | 72.23 |
| *:小型鱼类Small fish | ||||
从洄游习性看, 淡水定居性鱼类最多, 有54种, 包括鲤、鲫、鳊、

|
| 图 2 新洲水域鱼类生态类群 Fig. 2 Fish ecological types in Xinzhou waters |
从鱼类栖息空间划分, 底层鱼类最多, 有24种, 包括鲤、麦瑞加拉鲮、黑鳍鳈(Sarcocheilichthys nigripinnis Günther, 1873)、黄颡鱼(Pseudobagrus fulvidraco Richardson, 1846)和大口鲇(Silurus meriordinalis Chen, 1977)等, 所占比例为39.39%(图 2)。中上层鱼类次之, 为22种, 包括鲢、鳙、
据摄食类型看, 以杂食性鱼类最多, 有27种, 包括鲤、鲂、麦穗鱼、棒花鱼(Abbottina rivularis Basilewsky, 1855)、黑鳍鳈和黄颡鱼等(图 2);肉食性鱼类次之, 有23种, 如刀鲚、鱤、青鱼(Mylopharyngodon piceus Richardson, 1846)、翘嘴鲌、鳜和大口鲇等;碎屑食性鱼类最少, 仅有3种, 分别为鲫(Carassius auratus Linnaeus, 1758)、兴凯鱊(Acheilognathus chankaensis Dybowsky, 1872)和高体鳑鲏(Rhodeus ocellatus Kner, 1866)。
2.4 鱼类多样性分析从多样性指数看, 新洲水域鱼类多样性较高, Shannon-Weiner指数(H)、Wilhm改进指数(H′)和Simpson优势集中度指数(C)变化范围分别为2.14—2.64、1.92—2.48和0.83—0.96。Pielou均匀度指数(E)亦较高, 变化范围为0.67—0.76(表 4)。
| 类别Items | 物种数Species | H | H′ | E | C | |
| 区域Area | 鹅毛洲尾 | 52 | 2.52±0.46 | 2.19±0.38 | 0.63±0.07 | 0.94±0.18 |
| 新河口 | 54 | 2.48±0.19 | 2.27±0.21 | 0.61±0.12 | 0.96±0.36 | |
| 新洲头 | 50 | 2.64±0.59 | 2.48±0.12 | 0.67±0.16 | 0.94±0.49 | |
| 季节Season | 春季 | 30 | 2.27±0.18 | 1.92±0.37 | 0.67±0.19 | 0.83±0.25 |
| 夏季 | 46 | 2.14±0.25 | 2.18±0.51 | 0.55±0.28 | 0.94±0.11 | |
| 秋季 | 42 | 2.19±0.34 | 2.18±0.10 | 0.58±0.20 | 0.93±0.35 | |
| 冬季 | 34 | 2.24±0.13 | 2.32±0.41 | 0.63±0.08 | 0.92±0.03 | |
| H:香农威纳指数, Shannon-Weiner index; H′: Wilhm改进指数, Wilhm improvement index; E: Pielou均匀度指数, Pielou evenness index; C: Simpson优势集中度指数, Simpson dominance index | ||||||
新洲水域鱼类种类较多, 共64种(表 2), 但采样区域间的差别较小, 3个采样区域鱼类种数分别为52、54和50种(表 4)。鱼类物种数季节间差异明显, 夏、秋季种类较多, 分别为46和42种, 而春、冬季节较少, 分别仅有30和34种, 部分种类如华鳈(Sarcocheilichthys sinensis Bleeker, 1871)、青鱼、蛇鮈(Saurogobio dabryi Bleeker, 1871)、达氏鲌(Culter dabryi Bleeker, 1871)和团头鲂(Megalobrama amblycephala Yih, 1955)均出现在夏、秋季调查中, 而冬、春季未发现。新洲头鱼类的H、H′、E和C均高于新河口及鹅毛洲尾, 但多样性指数采样区域间差别不显著(P>0.05)。鱼类的H、H′、E和C均在冬季最高, 夏季次之, 季节间差异未达显著水平(P>0.05) (表 4)。
3 讨论 3.1 渔业资源现状与历史变化长江安庆江段江面开阔, 水流较缓, 下游江段在潮汐的顶托作用下泥沙加速沉积导致诸如新洲等的沙洲密布, 为鱼类的繁殖和索饵提供了优良的基础条件。本次新洲水域共调查到鱼类64种, 种类数与20世纪90年代的63种相近[7], 但明显低于2004年长江安庆段湖泊湿地鱼类调查的98种[6]。本次调查比张敏莹等[2]安庆江段调查的鱼类物种数多了近20种, 可能与研究区域间差异存在较大关系。本次研究对长江安庆段新洲3个不同生境类型的水域进行了调查, 新河口水域为浅型草滩, 鹅毛洲尾水域为浅型泥沙底质, 而新洲洲头水域为石质底质, 基本覆盖了各种鱼类栖息生境类型。张敏莹等[2]调查的沙漠洲和皖河口水域均为泥沙底质, 鱼类栖息生境较单一, 种类较少。本次研究与刘明典等[8]2015年长江安庆段鱼类的调查结果相近, 且鱼类种类组成相似。与芜湖和铜陵江段一样[18-19], 新洲水域鱼类也以鲤科鱼类物种数最多, 共29属40种, 占总物种数的62.50%。
从新洲水域鱼类组成来看, 鱼类存在明显资源小型化趋势, 与湖泊、水库等水体的研究结果相似[20]。大型经济鱼类数量减少, 青鱼、草鱼、鲢和鳙4种常见经济鱼类数量比例不足2%, 而小型鱼类比例达到渔获物总量的69.3%, 与2002—2004年相同江段小型鱼类占45.7%和2013—2014年的54.3%相比, 小型鱼类比例有明显增加(表 5)[2, 4]。鱼类群落小型化趋势在很大程度上是环境因素和捕捞因素共同作用的结果, 强大捕捞压力下, 高营养级捕食者(通常是个体较大、经济价值较高的种类)持续减少, 并导致渔获物组成向个体较小、营养层次较低、经济价值不高的种类转变[21]。另一方面, 自然水域中的小型鱼类会通过占据其他鱼类生态位从而加剧鱼类群落的不稳定性, 导致鱼类群落结构趋向小型化和单一化[22], 新洲水域小型鱼类比例高, 说明鱼类群落结构复杂度降低, 群落稳定性受到威胁, 需加强渔业资源保护。本次研究中, 部分洄游型鱼类(如日本鳗鲡Anguilla japonica Temminck et Schlegel, 1846)和河口鱼类(如鲻Mugil cephalus Linnaeus, 1758)未出现, 这与张晓可等[4]的研究一致, 但2002—2004年时洄游型物种还较多[2], 洄游型鱼类的减少可能与下游大量的闸坝建设及其他涉水工程有关。本次研究出现了如宽鳍鱲(Zacco platypus Temminck et Schlegel, 1846)、马口鱼(Opsariicjthys bidens Günther, 1873)、小口小鳔鮈和圆尾拟鲿(Pseudobagrus tenuis Günther, 1873)等先前文献中未采集到的物种。
| 变量Variables | 2002—2004年 | 2003—2010年 | 2013—2014年 | 2017年 |
| 参考文献References | 张敏莹等(2006)[3] | 徐东坡(2010)[2] | 张晓可(2016)[4] | 本次调查数据 |
| 网具类型Nets type | 定置张网 | 定置网 | 电拖网和多目刺网 | 网簖、地笼和多目刺网 |
| 渔获量Catches/kg | 43.19 | 1.39 | 135.4 | |
| 物种数Species | 46 | 50 | 35 | 64 |
| 四大家鱼比例Ratios of Chinese carps/% | 29.97 | 未统计 | <0.01 | 2.55 |
| 小型鱼类比例Ratios of small-sized fish/% | 45.65 | 46.30 | 54.30 | 69.34 |
新洲水域鱼类的Shannon指数变化范围为2.14—2.64, 处于Magurran[23]提出的多样性指数的一般范围(1.5—3.5)内, 同时丰富度指数和均匀度指数也处于正常范围内的较高水平(表 3)。丰富度指数和均匀度指数反映了水域中鱼类种类数与物种分布情况, 较高的丰富度指数和均匀度指数表明新洲水域鱼类群落结构复杂度较高, 群落具有一定稳定性。3个采样区域中, 新洲洲头水域鱼类多样性最高, 可能与复杂水文环境条件下形成的丰富多样栖息地环境有关。长江干流在新洲洲头分为北汊、中间夹江和南汊3股水道, 洲头北侧为夹江, 水体流速大, 水体混合度高, 喜激流鱼类种类多。洲头南侧为牛头山水道, 水流对洲头的冲击及水流方向的改变, 使得洲头南侧水体流速小, 形成回水区。回水区较缓的水体流速利于有机物在洲头水域的沉积, 浮游生物和底栖动物资源丰富, 从而为鱼类提供了丰富的食物来源[24]。同时水体泥沙沉积, 在洲头形成了大片浅滩, 枯水期部分露出水面, 丰水期没入水中, 水域环境良好, 有利于水生生物种群的生长与繁殖, 也为鱼类等提供了优良的栖息环境[25]。浅型沙洲水生植物种类及密度较高, 为不同繁殖和栖息类型鱼类提供了良好的生长繁殖环境, 因此鱼类种类多, 群落结构复杂[18]。从环境因子与多样性指数分析结果看, 新洲水域鱼类多样性与水体理化指标无显著相关性(P>0.05, 表 6)。另外, 在流域生态系统中, 不同河段或不同支流由于所处流域内空间位置上的差异, 即使其非生物环境条件较为一致, 但其生物环境因子及其生态过程往往存在差异, 这将导致这些河段或支流具有不同的鱼类群落结构[26]。
| 环境因子Environmental factors | H | H′ | E | C |
| 透明度Transparency | -0.135 | 0.008 | -0.640 | 0.430 |
| 总氮Total nitrogen | -0.104 | -0.304 | 0.129 | -0.213 |
| 总磷Total phosphorus | -0.224 | -0.101 | 0.422 | -0.522 |
| 叶绿素a Chlorophyll a | -0.360 | 0.294 | 0.025 | -0.173 |
| 高锰酸盐指数CODMn Permanganate Index | 0.019 | -0.447 | 0.722 | -0.776* |
| * P<0.05; H:香农威纳指数, Shannon-Weiner index; H′: Wilhm改进指数, Wilhm improvement index; E: Pielou均匀度指数, Pielou evenness index; C: Simpson优势集中度指数, Simpson dominance index | ||||
长江等河流系统的水文和水位等条件受季节性干旱和洪涝的影响, 具有更高的季节动态[27], 因此鱼类群落的物种组成和数量变化也比湖泊、水库等静水环境更为明显[28]。新洲水域鱼类群落季节间差别也非常明显, 夏、秋季鱼类物种数明显较冬、春季多, 季节间物种数最多相差18种。鱼类种群动态除受外界环境因素影响外, 其生活史不同阶段也影响着鱼类群落结构及其多样性, 如繁殖活动可引起大量补充群体的增加、洄游鱼类的周期性栖息地转化可引起鱼类群落组成的变化等[29]。绝大多数鱼类的繁殖活动集中在春季末期和夏季初期, 有大量补充群体加入, 因此夏、秋季鱼类物种数、个体数及多样性指数都较高, 这在本次研究中有明显的体现。冬季鱼类种类及多样性的显著下降可能与水温急剧降低造成的渔业资源贫乏有关[29]。因此, 本研究中鱼类多样性明显的季节变化很可能与鱼类自身潜在的繁殖期群体补充及越冬死亡有关。
新洲水域鱼类以杂食性鱼类为主, 这与黄浦江、黄河等大型河流的鱼类调查研究结果一致[12, 30-31]。与单一食性的鱼类相比, 杂食性鱼类具有更大生存优势, 其可摄食食物种类更加多样, 生态位幅度较宽, 因此对环境的适应能力更强。从空间分布来看, 底层鱼类在群落中占据优势地位, 这与被动型网具进行渔业资源调查方式有较大关系, 被动型网具通常放置在浅型河湾或沙洲附近, 底栖动物资源丰富, 能为更多鱼类提供充足的饵料[32]。本次研究中, 鱼类生态类群季节和采样区域间没有明显差异, 这可能与调查水域跨度较小有关。
3.3 鱼类多样性的影响因素自然水体中, 鱼类个体通过生长、繁殖和死亡等方式进行种群生活史, 同时也会通过迁入和迁出等行为方式来进行种间交流, 但总体上能够保持种群的动态平衡[33]。随着人类水域活动的加强, 鱼类种群平衡被破坏, 渔业资源小型化趋势明显。影响鱼类群落结构的主要人类活动主要有以下几个方面:(1)过度捕捞导致鱼类资源量明显下降、鱼类小型化。施炜纲等研究表明, 2001—2004春季禁渔期间(4—6月), 安庆江段鱼类物种多样性指数与中华绒螯蟹、刀鲚等的专项捕捞证的发放数量呈显著的负相关关系[34]。捕捞技术的提升, 渔民捕捞手段的多样化, 网具网眼越来越密, 导致捕捞鱼类的捕捞规格越来越小[35];另外, 鱼类繁殖季节大量亲鱼的捕捞对鱼类资源的补充群体造成的伤害也是渔业资源衰退的重要原因。(2)涉水工程建设破坏鱼类栖息环境, 洄游型鱼类产卵、繁殖需要的洄游通道被阻隔, 洄游型鱼类大量减少甚至灭绝[36]。建国以来, 安庆在其长江干流修建了诸如同马大堤、枞阳江堤和东兴圩等大大小小的江堤近300 km, 兴建整修大中型通江涵闸6座, 小型涵闸仅700座。同时, 湖口建闸形成江湖阻隔, 破坏了江湖复合生态系统, 导致沿江湿地溯河洄游鱼类消失, 江湖洄游鱼类生物多样性明显下降[6]。为满足经济发展的需要, 近年更是修建了码头、桥梁等涉水工程, 减少了鱼类适宜的栖息范围, 进一步压缩了鱼类的生存空间[8]。(3)航运繁忙, 安庆江段位于长江三角洲城市群上游, 是皖赣鄂三省交界处及全国重要的综合交通枢纽, 被视为连接中游与下游航道运输的咽喉要道, 航运十分繁忙。资料显示, 安庆江段船舶日均流量2007年为1375艘次, 2008年为1188艘次, 2007年长江安庆段各港口、码头进出港船舶达40846艘次。船舶数量大幅增加, 压缩了鱼类的活动空间, 直接影响到鱼类栖息地质量, 干扰了鱼类的摄食、繁殖和聚群等正常的行为活动。(4)外来种入侵, 调查发现, 麦瑞加拉鲮等外来种在渔获物中占有一定比列, 这些鱼类适应能力强、生长快、繁殖率高、扩散迅速, 与土著鱼类在空间、食物等方面已成竞争规模, 对土著种生存造成较大威胁, 从多方面威胁鱼类群落多样性的稳定。
4 结论与建议本研究对长江安庆江段新洲水域鱼类种类组成与多样性进行了周年调查, 共鉴定鱼类64种, 隶属6目11科48属, 主要为淡水定居性鱼类, 江河洄游型和江海洄游型鱼类较少。新洲水域水质环境良好, 栖息生境多样, 水生植被物种丰富, 为鱼类等水生生物提供了优良的生长繁殖条件。新洲水域小型鱼类占渔获物总数量的65%以上, 渔业资源小型化明显。基于长江水域渔业资源衰退和鱼类资源小型化的现状, 建议加强对长江渔业的监管与引导, 以确保长江下游渔业资源的可持续利用。
| [1] |
庄平, 王幼槐, 李圣法, 邓思明, 李长松, 倪勇. 长江口鱼类. 上海: 上海科学技术出版社, 2006.
|
| [2] |
徐东坡.长江下游鱼类群落结构及物种多样性的研究[D].南京: 南京农业大学, 2010.
|
| [3] |
张敏莹, 徐东坡, 刘凯, 施炜纲. 长江安庆江段鱼类调查及物种多样性初步研究. 湖泊科学, 2006, 18(6): 670-676. DOI:10.3321/j.issn:1003-5427.2006.06.017 |
| [4] |
张晓可, 于道平, 王慧丽, 万安, 蒋泽球, 陈寿文. 长江安庆段江豚主要栖息地鱼类群落结构. 生态学报, 2016, 36(7): 1832-1839. |
| [5] |
陈大庆, 段辛斌, 刘绍平, 施炜纲, 王博. 长江渔业资源变动和管理对策. 水生生物学报, 2002, 26(6): 685-690. DOI:10.3321/j.issn:1000-3207.2002.06.018 |
| [6] |
卢松, 陆林, 凌善金, 宣国富. 人类活动对安庆沿江湖泊湿地影响的初步研究. 长江流域资源与环境, 2004, 13(1): 65-71. DOI:10.3969/j.issn.1004-8227.2004.01.012 |
| [7] |
胡菊英, 姚闻卿. 长江下游安徽江段的鱼类. 安徽大学学报:自然科学版, 1996, 26(1): 96-101. |
| [8] |
刘明典, 李鹏飞, 黄翠, 段辛斌, 陈大庆, 刘绍平. 长江安庆段春季鱼类群落结构特征及多样性研究. 水生态学杂志, 2017, 38(6): 64-71. |
| [9] |
Scheffer M, Carpenter S, Foley J A, Folke C, Walker B. Catastrophic shifts in ecosystems. Nature, 2001, 413(6856): 591-596. DOI:10.1038/35098000 |
| [10] |
朱松泉. 中国淡水鱼类检索. 南京: 江苏科学技术出版社, 1995.
|
| [11] |
陈宜瑜, 褚新洛, 罗云林. 中国动物志:硬骨鱼纲:鲤形目(中卷). 北京: 科学出版社, 1998.
|
| [12] |
茹辉军, 王海军, 赵伟华, 沈亚强, 王勇, 张晓可. 黄河干流鱼类群落特征及其历史变化. 生物多样性, 2010, 18(2): 179-186. |
| [13] |
Pinkas L, Oliphant M S, Iverson I L K. Food Habits of Albacore, Bluefin Tuna, and Bonito in California Waters. California:Department of Fish and Game, 1971, 152: 1-105. |
| [14] |
陈国宝, 李永振, 陈新军. 南海主要珊瑚礁水域的鱼类物种多样性研究. 生物多样性, 2007, 15(4): 373-381. DOI:10.3321/j.issn:1005-0094.2007.04.006 |
| [15] |
Shannon C E, Weaver W. The Mathematical Theory of Communication:Theory of Communication. Urbana: University of Illinois Press, 1949.
|
| [16] |
Wilhm J L. Use of biomass units in Shannon's formula. Ecology, 1968, 49(1): 153-156. DOI:10.2307/1933573 |
| [17] |
Pielou E C. Ecological Diversity. New York:Wiley, 1975, 1-165. |
| [18] |
严云志, 郭丽丽, 李国龙. 长江铜陵段老洲水域鱼类资源的初步调查研究. 安徽师范大学学报:自然科学版, 2006, 29(6): 575-578. |
| [19] |
邓朝阳, 朱仁, 严云志. 长江芜湖江段鱼类多样性及其群落结构的时空格局. 淡水渔业, 2013, 43(1): 28-36. DOI:10.3969/j.issn.1000-6907.2013.01.006 |
| [20] |
毛志刚, 谷孝鸿, 曾庆飞, 周露洪, 王小林, 吴林坤, 曹萍, 孙明波. 太湖鱼类群落结构及多样性. 生态学杂志, 2011, 30(12): 2836-2842. |
| [21] |
Pauly D, Christensen V, Dalsgaard J, Froese R, Torres F Jr. Fishing down marine food webs. Science, 1998, 279(5352): 860-863. DOI:10.1126/science.279.5352.860 |
| [22] |
Rochet M J, Trenkel V M. Which community indicators can measure the impact of fishing? A review and proposals. Canadian Journal of Fisheries and Aquatic Sciences, 2003, 60(1): 86-99. DOI:10.1139/f02-164 |
| [23] |
Magurran A E. Why diversity?//Magurran A E, ed. Ecological Diversity and Its Measurement. Dordrecht: Springer, 1988: 1-5.
|
| [24] |
于道平, 黄敏毅, 赵凯, 陈寿文. 长江东流河道整治对长江江豚种群数量的影响. 兽类学报, 2012, 32(4): 330-334. |
| [25] |
杜浩, 班璇, 张辉, 危起伟, 陈大庆. 天然河道中鱼类对水深、流速选择特性的初步观测——以长江江口至涴市段为例. 长江科学院院报, 2010, 27(10): 70-74. DOI:10.3969/j.issn.1001-5485.2010.10.015 |
| [26] |
Grenouillet G, Pont D, Hérissé C. Within-basin fish assemblage structure:the relative influence of habitat versus stream spatial position on local species richness. Canadian Journal of Fisheries and Aquatic Sciences, 2004, 61(1): 93-102. DOI:10.1139/f03-145 |
| [27] |
Allan J D, Castillo M M. Stream Ecology:Structure and Function of Running Waters. 2nd ed. Netherlands: Springer, 2007.
|
| [28] |
Grossman G D, Moyle P B, Whitaker Jr J O. Stochasticity in structural and functional characteristics of an Indiana stream fish assemblage:a test of community theory. The American Naturalist, 1982, 120(4): 423-454. DOI:10.1086/284004 |
| [29] |
王文剑, 储玲, 司春, 朱仁, 陈文豪, 陈方明, 严云志. 秋浦河源国家湿地公园溪流鱼类群落的时空格局. 动物学研究, 2013, 34(4): 417-428. |
| [30] |
陈小华, 李小平, 程曦. 黄浦江和苏州河上游鱼类多样性组成的时空特征. 生物多样性, 2008, 16(2): 191-196. DOI:10.3321/j.issn:1005-0094.2008.02.013 |
| [31] |
李捷, 李新辉, 谭细畅, 李跃飞, 何美峰, 罗建仁, 林建志, 苏少芳. 广东肇庆西江珍稀鱼类省级自然保护区鱼类多样性. 湖泊科学, 2009, 21(4): 556-562. DOI:10.3321/j.issn:1003-5427.2009.04.015 |
| [32] |
胡忠军, 吴昊, 陈立侨, 刘其根. 长江口青草沙水库正式供水前的鱼类群落结构特征. 水产学报, 2012, 36(8): 1234-1241. |
| [33] |
殷名称. 鱼类生态学. 北京:中国农业出版社, 1995, 188-189. |
| [34] |
施炜纲, 刘凯, 张敏莹, 徐东坡. 春季禁渔期间长江下游鱼虾蟹类物种多样性变动(2001-2004年). 湖泊科学, 2005, 17(2): 169-175. DOI:10.3321/j.issn:1003-5427.2005.02.013 |
| [35] |
王银平.太湖鲚种群特征及鱼类种间关系研究[D].南京: 中国科学院南京地理与湖泊研究所, 2016.
|
| [36] |
朱轶, 吕偲, 胡慧建, 王忠锁, 贾亦飞, 何木盈, 黄小军, 雷光春. 三峡大坝运行前后西洞庭湖鱼类群落结构特征变化. 湖泊科学, 2014, 26(6): 844-852. |