 2020, Vol. 40
2020, Vol. 40文章信息
- 尹国丽, 李亚娟, 张振粉, 李小龙, 张晓燕, 师尚礼
- YIN Guoli, LI Yajuan, ZHANG Zhenfen, LI Xiaolong, ZHANG Xiaoyan, SHI Shangli
- 不同草田轮作模式土壤养分及细菌群落组成特征
- Characteristics of soil nutrients and bacterial community composition under different rotation patterns in grassland
- 生态学报. 2020, 40(5): 1542-1550
- Acta Ecologica Sinica. 2020, 40(5): 1542-1550
- http://dx.doi.org/10.5846/stxb201901100088
-
文章历史
- 收稿日期: 2019-01-10
- 网络出版日期: 2019-12-17





2. 草业生态系统教育部重点实验室, 兰州 730070
2. Key laboratory of Grassland Ecosystem, Ministry of Education, Sino-U. S. Centers for Grazingland Ecosystem Sustainability, Lanzhou 730070, China
以紫花苜蓿(Medicago sativa L.)为代表的多年生豆科牧草是我国主要种植加工的一大类饲草作物, 在现代养殖业发展中有着不可或缺的作用, 但是连作障碍一直是多年生牧草种植的瓶颈问题。近些年已有大量学者进行了探索, 证实连作障碍的发生不仅与土壤理化性质改变有关, 还与土壤微生物组成有关[1-2], 土壤微生物作为土壤中的活性因子, 是反应土壤质量最敏感的指标, 近年来已有大量的研究表明在土壤微生物与连作障碍之间存在密切关系[3-4]。
草田轮作是农业生产中消减连作障碍最广泛的应用措施, 轮作通过增加作物组分丰富了地上、地下生物多样性, 改变了土壤微生态环境[5], 大量试验证明, 轮作不仅可以协调不同作物之间养分吸收的局限性, 避免养分失衡, 提高土壤养分的有效性和酶活性, 还可以通过根系分泌物调整根围微生物群落组成, 减少土传病虫害的发生, 提高经济效益[6-7], 但不同轮作模式对土壤理化性质的影响不尽相同。土壤微生物作为土壤生态系统重要组成部分, 其群落结构和多样性主要受植物物种组成和土壤养分状况的影响[8], 不同作物释放的根系分泌物存在差别, 也影响了土壤微生物群落生长代谢和群落结构[9-11]。细菌是土壤微生物中分布最广, 数量最多的部分[12], 占土壤微生物总数的70%—90%, 是土壤养分变化的敏感指标之一[13]。研究微生物群落组成及多样性一直是揭示植物-微生物互作关系机制的热点问题[14-15]。目前高通量测序技术的不断发展, 提高了研究者对环境中微生物群落组成和功能的认知水平[16-17]。
科学合理的轮作模式可有效缓解连作障碍引起的土壤环境恶化, 使土壤中微生物活性增强、多样性增加, 为作物生长提供健康稳定的土壤生态环境[18-19]。目前关于紫花苜蓿草田轮作的研究报道相对较多, 但前人的研究多集中在轮作对作物田土壤肥力及产量的影响[20-22], 缺乏紫花苜蓿-作物轮作系统中土壤细菌组成方面的研究报道。鉴于此, 本试验分析5龄紫花苜蓿草地和不同作物轮作模式下, 土壤养分因子含量和土壤细菌群落结构的组成及其相互关系, 旨在较深层次上探索紫花苜蓿连作障碍的形成机制, 为采用轮作措施减轻紫花苜蓿连作障碍提供实践依据, 并为半干旱地区草田轮作和草地生态农业的可持续发展提供理论基础。
1 材料与方法 1.1 试验地概况试验地位于在甘肃省武威市凉州区甘肃农业大学黄羊镇牧草站(103°5′E, 37°30′N, 海拔1776 m)。位于甘肃河西走廊东端, 属冷温带干旱区, 典型的大陆性气候, 日照充足, 春季多风沙, 夏季有干热风。降水年际变化不大, 但季节变化较大, 多年平均降水量160 mm左右, 主要集中在7、8、9月份, 冬春季干旱, 降水无法满足作物生长的需要。年蒸发量2400 mm, 年平均气温7.2 ℃, 一月最低气温-27.7 ℃, 七月最高气温34.0 ℃, 0 ℃积温3513.4 ℃, 10 ℃积温2985.4 ℃。全年无霜期156 d, 绝对无霜期118 d, 年日照时数2945 h。土壤以荒漠灌淤土为主, 粉沙壤质, 土层深厚。
1.2 供试材料供试材料小麦品种为杂交种春宁4号(Triticum aestivum L.cv.Chunning No.4), 玉米品种为杂交种陇单4号(Zea mays L.cv.Longdan No.4), 紫花苜蓿品种为甘农3号(Medicago sativa L.cv.Gannong No.3)。小麦及玉米种子市售, 紫花苜蓿种子由甘肃农业大学草业学院提供。
1.3 试验设计选取地势平坦的紫花苜蓿草地进行草田轮作试验。设计如表 1所示, 设4个处理。试验小区面积为4 m×5 m, 不同小区间设1 m隔离带, 每个处理3个重复, 不同小区的土壤肥力及管理方式相同。
| 编号No. | 轮作模式Rotation patterns |
| A5W1 | 5a紫花苜蓿-1a小麦5a Alfalfa-1a Wheat |
| A5C1 | 5a紫花苜蓿-1a玉米5a Alfalfa-1a Corn |
| A5W2 | 5a紫花苜蓿-2a小麦5a Alfalfa-2a Wheat |
| A5C2 | 5a紫花苜蓿-2a玉米5a Alfalfa-2a Corn |
于2017年7月中旬采集土壤样品, 五点法取样, 用内径40 mm的土壤取样器取0—30 cm土层(耕作层)土样。每次采集后用70%的酒精对土壤采样器进行消毒处理, 以避免不同样品之间的污染。采得土样去除植物残体和石砾等杂物后放入灭菌袋, 低温运输至实验室, 尽快进行土壤DNA的提取和其余操作。
1.4.1 土壤养分测定土壤有机质含量采用重铬酸钾法测定;土壤全氮采用凯氏定氮法测定;土壤碱解氮采用碱解扩散法;土壤全钾采用火焰光度法;土壤有效钾含量先用醋酸铵浸提, 再用火焰光度法测定;土壤全磷采用HClO4-H2SO4消煮法;土壤速效磷采用碳酸氢钠浸提-铝锑抗比色法测定[23]。
1.4.2 土壤微生物DNA提取和16S rRNA基因高通量测序将采集的样品拣除杂质后, 同一样品充分混合使之均匀, 准确称取1.00 g新鲜土样。采用OMEGA土壤DNA提取试剂盒提取土壤总DNA, 提取步骤参考试剂盒说明书。1%的琼脂糖电泳和紫外分光光度法检测所提取DNA的质量和浓度, 检测合格后利用细菌16S rRNA基因的通用引物314F (5′-CCTACGGGNGGCWGCAG-3′)
-805R(5′-GACTACHVGGGTATCTAATCC-3′)扩增土壤微生物16S rRNA V3+V4区基因片段。PCR反应体系如下:Phusion Master Mix(1X)15 μL, DNA模版(1 ng/μL)10 μL, 正、反向引物(0.2 μmol/L)各3 μL, ddH2O补至30 μL。PCR扩增条件为:95℃, 3 min;(95 ℃, 30 s;55℃, 30 s;72 ℃, 30 s), 25个循环;16 ℃10 min[24]。每个样品3个重复。琼脂糖凝胶电泳法检测扩增结果, 检测合格后送交至上海生工生物科技公司进行后续实验和高通量测序。
1.5 序列分析获得原始序列数据后, 采用QIIME 1.8.0分析OTUs的质量分析、Alpha多样性分析等。Alpha多样性指数中, Chao1和Observed_species为菌群丰度指数。Chao1计算公式为: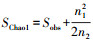
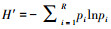
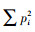
采用Microsoft Excel 2013软件对数据进行处理和作图;用SPSS 24.0进行单因素方差分析(one-way ANOVA), 采用最小显著差异法(LSD)比较数据组间的差异, 用Person相关系数评价不同因子间的相关性。用SPSS 24.0统计软件完成相关关系分析。
2 结果与分析 2.1 不同轮作模式对土壤养分的影响如表 2所示, 紫花苜蓿轮作作物1年时, A5C1处理的有机质含量为12.3 g/kg, A5W1处理为11.85 g/kg, 轮作第2年, 无论后茬作物是玉米还是小麦, 有机质含量较轮作第1年均呈现下降趋势, 轮作玉米处理第2年比第1年下降3.66%, 而小麦处理下降了7.68%。其余指标均呈现出相似的规律, 就玉米轮作组而言, 第2年与第1年相比, 全氮下降了2.17%, 全磷下降了9.28%, 全钾、碱解氮、速效磷、速效钾呈现不同程度的下降, 其中, 下降幅度最大的是速效磷, 下降了25.8%, 下降幅度最小的是碱解氮, 下降了0.47%。轮作小麦和轮作玉米表现出了相似的规律, 都是轮作第2年各指标较第1年下降, 下降幅度最大的是速效磷, 下降了38.37%, 下降幅度最小的是速效钾, 为2.53%。
| 处理 Treatments |
有机质 Organic matter/(g/kg) |
全氮 TN/(g/kg) |
全磷 TP/(g/kg) |
全钾 TK/(g/kg) |
碱解氮 AN/(mg/kg) |
速效磷 AP/(mg/kg) |
速效钾 AK/(mg/kg) |
| A5W1 | 11.85±0.23a | 0.99±0.01a | 1.02±0.01a | 76.81±1.77a | 28.60±0.38a | 105.51±0.45a | 208.15±1.01a |
| A5C1 | 12.30±0.20a | 0.93±0.01b | 0.97±0.01a | 65.95±1.62a | 27.59±1.57b | 105.20±1.37b | 213.15±1.12a |
| A5W2 | 10.94±0.07c | 0.94±0.02b | 0.92±0.02a | 75.56±1.17a | 24.75±1.06c | 65.02±1.18d | 202.88±0.70a |
| A5C2 | 11.40±0.21b | 0.91±0.01c | 0.88±0.01b | 61.04±0.07a | 27.46±0.38b | 78.06±0.93c | 205.51±1.33a |
| 同列不同字母表示处理间差异显著(P < 0.05) | |||||||
测序得到的土壤细菌群落序列读数和OTUS数(表 3), 在相似度大于97%的水平上4个轮作模式共获得高质量目标序列42743条。Clean Reads Rate值均高于80%, 说明测序质量可以反映土壤细菌群落组成情况。不同轮作模式统计分析表明, 土壤细菌OTUS种类数无显著差异(P>0.05)。
| 轮作模式 Rotation patterns |
高质量目标序列 Number of valid sequences |
OTUS | Clean Reads Rate/% |
| A5W1 | 10421Aa | 1891Aa | 85 |
| A5C1 | 8514Ab | 1731Aa | 84 |
| A5W2 | 12410Ba | 2281Aa | 83 |
| A5C2 | 14398Aa | 2283Aa | 85 |
| 大写字母表示同行之间比较, 小写字母同列之间比较, 不同字母在0.05水平下差异显著 | |||
由表 4所示, 除ACE指数外, 其余3个指数都呈现出A5C2 > A5W1 > A5W2 > A5C1的趋势, ACE指数虽为A5W2 > A5W1, 但差异不显著。综上, 不同指数反映出了基本一致的趋势。
| 轮作模式 Rotation pattern |
Simpson | Chao1 | ACE | Shannon |
| A5W1 | 0.9841 | 1992.0 | 2350.4 | 8.67 |
| A5C1 | 0.9690 | 1569.0 | 2000.3 | 7.43 |
| A5W2 | 0.9827 | 1700.0 | 2363.2 | 8.24 |
| A5C2 | 0.9933 | 2127.0 | 2696.0 | 9.22 |
在门水平上, 除极少部分细菌(0.02%—0.05%)未被分类外, 4种轮作模式下土壤细菌主要分布在23个细菌门类。由图 1可知, 优势菌门为变形菌门(Proteobacteria)(25.30%—41.90%)和放线菌门(Actinobacteria)(14.50%—19.80%)以及厚壁菌门(Firmicutes)(3.70%—23.70%)、酸杆菌门(Acidobacteria)(5.40%—13.90%)。其次, 拟杆菌门(Bacteroidetes)、绿弯菌门(Chloroflexi)、浮霉菌门(Planctomycetes)和芽单胞菌门(Gemmatimonadetes)的丰度也相对较高, 其余菌门在各处理中均有分布, 相对丰度因处理不同而存在差异, 但差异不显著(P>0.05)。另外, 优势菌在不同轮作模式下的组成也不同, A5W2中变形菌门(Proteobacteria)含量很高, 而厚壁菌门(Firmicutes)含量却较低, 而这两种菌在其他轮作模式下含量较为稳定。

|
| 图 1 不同轮作模式土壤在门水平上的细菌类群比较 Fig. 1 Comparison of bacteria groups at phylum level in the soils under different rotation sequences A5W1: 5a紫花苜蓿-1a小麦; 5a紫花苜蓿-2a小麦; A5C1: 5a紫花苜蓿-1a玉米;A5C2: 5a紫花苜蓿-2a玉米 |
在属的分类水平, 除未被分类的细菌属外(41.31%—53.78%), 各轮作模式土壤细菌相对丰度排序前13的细菌菌属分布见图 2。不同轮作模式下土壤优势属基本相同, 其中丙酸杆菌属(Propionibacterium)、芽孢杆菌属(Bacillus)、链球菌属(Streptococcus)以及奈瑟氏球菌属(Neisseria)为优势属。但不同轮作模式下土壤各细菌属所占相对丰度大小存在差异, 如芽孢杆菌属(Bacillus)在A5W1中相对丰度最高, 但在A5W2中却相对较低。

|
| 图 2 不同轮作模式土壤在属水平上的细菌类群比较 Fig. 2 Comparison of bacterial groups in the soils at the genus level under different rotation patterns |
由图 3可知, 4种轮作模式下土壤细菌物种相对丰度共提取4个主成分, 累积贡献率达96.82%, 其中第1主成分(PC1)和第2主成分(PC2)贡献率分别为68.25%和22.32%。从图 3中可以看出4种轮作模式中A5W1和A5C1分布在PC1轴负方向, A5W2和A5C2分布在正方向。A5W1和A5C2序列分布在PC2轴正方向, A5C1和A5W2分布在负方向。

|
| 图 3 不同轮作模式土壤在属水平细菌相对丰度主成分分析 Fig. 3 Principal component analysis of relative abundance of soil bacteria at genus level under different rotation patternss |
载荷因子反映主成分与细菌属水平丰度的相关系数, 载荷因子越高, 表示该细菌属对主成分的影响越大[12]。由表 5可知, 与第一主成分(PC1)具有较高相关性的细菌属主要有固氮菌属Azotobacter、硝化螺旋菌属Nitrospira、小梨形菌属Pirellula、Iamia菌属和芽孢杆菌属Bacillus, 与第二主成分(PC2)具有较高相关性的细菌属则主要有Flavisolibacter菌属、德沃斯氏菌属Devosia、德沃斯氏菌属、拜纳蒙纳斯属Balneimonas、Adhaeribacter菌属(表 5)。
| 细菌属Bacterial genus | PC1 | PC2 |
| Azotobacter | 0.91* | 0.35 |
| Flavisolibacter | -0.61 | 0.55* |
| Arthrobacter | -0.01 | 0.34 |
| Rhodoplanes | 0.11 | 0.15 |
| Nitrospira | 0.86* | -0.13 |
| Sphingomonas | 0.53 | 0.1 |
| Lysobacter | -0.22 | -0.49 |
| Fimbriimonas | -0.6 | -0.25 |
| Janthinobacterium | -0.59 | 0.48 |
| Pseudomonas | 0.9 | 0.3 |
| Devosia | 0.5 | 0.57* |
| Flavisolibacter | -0.61 | 0.55* |
| Arthrobacter | -0.01 | 0.34 |
| Rhodoplanes | 0.66 | 0.15 |
| DA101 | 0.12 | -0.13 |
| Sphingomonas | 0.53 | 0.1 |
| Bacillus | 0.88* | 0.54 |
| Iamia | 0.81* | -0.13 |
| Pirellula | 0.96* | 0.19 |
| Variovorax | -0.5 | 0.53 |
| Balneimonas | 0.18 | 0.75* |
| Adhaeribacter | -0.56 | 0.57* |
| 表中带*的为两个主成分中相关性前5的细菌属 | ||
由图 4可见, 在属水平上, A5W1和A5C1的组成最接近, 其次是A5C2, A5W2的组成与前两者相差较大, 说明作物轮作的时间越长, 细菌组成差异越大。

|
| 图 4 不同轮作模式土壤在属水平细菌组成聚类分析 Fig. 4 Cluster analysis of genus level composition of soil bacteria under different rotation patterns |
由表 6可知, 土壤养分因子与各细菌属的相关性不同, 有机质与丙酸杆菌属Propionibacterium及固氮菌属Azotobacter呈现显著相关关系(P < 0.05), 而与芽孢杆菌属Bacillus为极显著正相关关系(P < 0.01)。全钾与链球菌属Streptococcus呈现显著相关关系(P < 0.05), 土壤碱解氮与丙酸杆菌属Propionibacterium呈显著正相关关系(P < 0.05), 与固氮菌属Azotobacter呈现极显著负相关关系(P < 0.01);而速效磷与丙酸杆菌属Propionibacterium、芽孢杆菌属Bacillus及固氮菌属Azotobacter均呈显著正相关关系(P < 0.05)。
| 有机质 Organic matter |
全氮 TN |
全磷 TP |
全钾 TK |
碱解氮 AN |
速效磷 AP |
速效钾 AK |
|
| 丙酸杆菌属Propionibacterium | 0.826* | 0.752 | 0.417 | -0.762 | 0.866* | 0.886* | 0.751 |
| 芽孢杆菌属Bacillus | 0.913** | 0.722 | 0.658 | 0.715 | 0.715 | 0.826* | 0.762 |
| 链球菌属Streptococcus | 0.514 | 0.325 | 0.726 | 0.952** | 0.569 | 0.741 | 0.763 |
| 固氮菌属Azotobacter | 0.889* | -0.526 | 0.688 | 0.652 | -0.956** | 0.862* | 0.755 |
| 硝化螺旋菌属Nitrospira | 0.714 | 0.924 | 0.627 | 0.657 | 0.689 | -0.715 | 0.627 |
| P < 0.05;**P < 0.01 | |||||||
土壤细菌作为土壤中最重要的活性组分, 其多样性水平也是反映土壤环境的最重要指标。通过本研究结果可以看出, 后茬作物种类和轮作时间对土壤细菌群落结构都有明显的影响。其他学者的研究也发现了相似的规律, 如王芳等[25]利用Illumina Mi Seq技术对不同连作及轮作方式下大豆根际细菌进行研究, 发现连作时间、轮作作物种类和轮作顺序3个因素对对土壤细菌多样性都会产生影响。梁志婷[26]在甘肃陇东地区进行了不同粮草轮作模式对土壤微生物多样的研究结果也与本研究相似。本研究的结果显示, 轮作作物和轮作时间2个因子对土壤细菌多样性的影响是普遍的, 不同作物上的研究均支持这一结论。因此, 在利用轮作改善土壤微生态时, 应充分考虑2个因子, 使其发挥最佳效果。另一方面, 本研究也提出了新的科学问题, 即在轮作体系中, 作物种类和轮作时间这2个因子, 哪一个对土壤微生物多样性的组成影响更大, 这将是值得深入研究的新课题。
3.2 不同轮作模式下土壤细菌群落组成土壤细菌群落结构受种植作物的影响[27], 作物种类不同其土壤微生物所处环境不同, 细菌组成和丰度大小有差异, 不同的农田土壤中存在共有和特异的细菌种群[28]。本研究发现变形菌门(Proteobacteria)和放线菌门(Actinobacteria)以及厚壁菌门(Firmicutes)、酸杆菌门(Acidobacteria)为各轮作顺序丰度较高的菌, 这一结果与Sapp等[29]、Liu等[30]和王伏伟等[31]对土壤细菌的研究结果一致。在属的水平, 芽孢杆菌属(Bacillus)在耕层丰度较高, 该属为不产氧光合细菌, 其功能与禾本科作物联合固氮作用紧密相关[32], 这与4个轮作顺序取样阶段均为禾本科作物相符。研究表明, 某些植物能够与土壤微生物产生互作作用, 对植物病害控制产生一定效果, 同时也可能会引起某种作物感染病害[33]。本研究发现, 5a紫花苜蓿-1a玉米序列土壤中芽孢杆菌属(Bacillus)和奈瑟氏球菌属(Neisseria)丰度较高, 该菌属能够通过产生几丁质酶或细菌素的方式对土壤线虫发育产生抑制作用, 有生防效果[34-35]。研究结果在揭示在河西走廊灌区连作苜蓿后轮作禾本科作物在改善土壤肥力的同时, 对胞土壤害虫防治效果较好。轮作后Fimbriimonas菌属丰度较高, 此菌具有固氮和脱氮特征, 在维持氮平衡方面起重要作用[36]。
3.3 轮作时间与土壤细菌群落细菌群落是土壤微生物中最主要的一类微生物, 是土壤微生物多样性的重要指标。应用高通量测序分析技术, 可以比较准确地反映细菌群落特征[37-38]。本研究通过高通量测序技术对比研究了不同轮作模式下土壤细菌多样性, 结果发现, 不同的轮作模式下细菌群落结构具有一定的差异。但是主要优势菌群保持稳定, 都以变形菌门(Proteobacteria)丰度最高, 土壤中每一特定性的菌类均具有不同的功能特性, 变形菌是一类适应性很强的细菌, 该类菌中即包含动植物的病原菌也存在抑制致病菌的有益菌[39-40], 但是不同轮作模式的细菌组成也有不同, 轮作1年的比轮作2年的芽单胞菌门的丰度要高。芽单胞菌类是有益于作物生长的细菌, 轮作1年时, 该类菌群数量明显高于第2年, 这可以从土壤微生物特定菌类的增加来解释轮作栽培有益于作物生物生长。轮作栽培由于不同的作物对养分和水分吸收以及根系分泌物不同, 因而能够改变土壤微生物的生态环境。吴宏亮对砂田土壤进行轮作栽培研究, 发现轮作改变砂田土壤微生物区系结构, 提高微生物多样性[41]。本研究中, 无论后茬作物玉米还是小麦, 轮作2年实际上又造成了禾本科作物的连作, 因此, 其土壤微生态的组成要劣于轮作1年的。
4 结论在多年种植苜蓿地和不同禾本科作物复种轮作模式下, 不同轮作模式对土壤养分各指标有不同程度的影响, 后茬作物无论是玉米还是小麦, 轮组第2年土壤有机质、全氮、全磷、全钾、碱解氮、速效磷和速效钾的含量都较第1年有明显的下降。4种轮作模式下占优势的菌门为变形菌门、放线菌门和厚壁菌门。土壤细菌群落属组成中以丙酸杆菌属(Propionibacterium)、芽孢杆菌属(Bacillus)、链球菌属(Streptococcus)以及奈瑟氏球菌属(Neisseria)为优势属等为主。土壤细菌多样性以A5C2轮作模式下最高, A5C1最低;聚类分析显示A5W1和A5C1的组成最接近, 其次是A5C2, A5W2的组成与前两者相差较大。土壤养分因子含量与几种优势属呈现了不同程度的相关性。研究结果揭示了河西走廊灌溉区种植多年紫花苜蓿后复种小麦和玉米改善土壤肥力和防治某些土传植物病害微生物的机理。
| [1] |
谢泽宇.陇中黄土高原苜蓿-作物轮作对土壤碳氮组分及其相关微生物活性的影响[D].甘肃农业大学, 2017.
|
| [2] |
Yang N, Wang Z H, Gao Y J, Li K, Sukhdev S. Malhi. Effects of planting soybean in summer fallow on wheat grain yield, total N and Zn in grain and available N and Zn in soil on the Loess Plateau of China. European Journal of Agronomy, 2014, 58: 63-72. DOI:10.1016/j.eja.2014.05.002 |
| [3] |
秦舒浩, 曹莉, 张俊莲, 师尚礼, 王蒂. 轮作豆科植物对马铃薯连作田土壤速效养分及理化性质的影响. 作物学报, 2014, 40(8): 1452-1458. |
| [4] |
周岚, 杨永, 王占海, 陈阜, 曾昭海. 玉米-大豆轮作及氮肥施用对土壤细菌群落结构的影响. 作物学报, 2013, 39(11): 2016-2022. |
| [5] |
梁志婷.陇东旱塬不同粮草轮作模式下土壤微生物群落特征及土壤养分的研究[D].兰州大学, 2018.
|
| [6] |
虎德钰, 毛桂莲, 许兴. 不同草田轮作方式对土壤微生物和土壤酶活性的影响. 西北农业学报, 2014, 23(09): 106-113. |
| [7] |
安志刚, 郭凤霞, 陈垣, 白刚. 连作自毒物质与根际微生物互作研究进展. 土壤通报, 2018, 49(03): 750-756. |
| [8] |
Tiemann L K, Grandy A S, Atkinson E E, Marin-Spiotta E, McDaniel M D. Crop rotational diversity enhances belowground communities and functions in an agroecosystem. Ecology Letters, 2015, 18(8): 761. DOI:10.1111/ele.12453 |
| [9] |
毕博远, 韩凤朋. 黄土高原不同种植年限紫花苜蓿根系分泌物GC-MS分析. 草地学报, 2018, 26(03): 611-617. |
| [10] |
李国斌, 李光跃, 孙窗舒, 贾鑫, 陈贵林. 干旱胁迫对蒙古黄芪生物量及其根际微生物种群数量的影响. 西北植物学报, 2015, 35(9): 1868-1874. |
| [11] |
刘丹丹, 李敏, 刘润进. 我国植物根围促生细菌研究进展. 生态学杂志, 2016, 35(3): 815-824. |
| [12] |
李金融, 侯湖平, 王琛, 张绍良, 马静, 丁忠义, 黄磊, 董健, 杨永均. 基于高通量测序的复垦土壤细菌多样性研究. 环境科学与技术, 2018, 41(12): 148-157. |
| [13] |
Chen D M, Cheng J H, Chu P F, Hu S J, Xie Y C, Tuvshintogtokh I, Bai Y F. Regional-scale patterns of soil microbes and nematodes across grasslands on the Mongolian plateau:relationships with climate, soil, and plants. Ecography, 2015, 38(6): 622-631. DOI:10.1111/ecog.01226 |
| [14] |
Fierer N, Leff J W, Adams B J, Caporaso J G, Fierer N. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. Proceedings of the National Academy of Sciences, 2012, 109(52): 21390-21395. DOI:10.1073/pnas.1215210110 |
| [15] |
时鹏, 高强, 王淑平, 张妍. 玉米连作及其施肥对土壤微生物群落功能多样性的影响. 生态学报, 2010, 30(22): 6173-6182. |
| [16] |
Nannipieri P, Ascher J, Ceccherini M T, Landi L, Pietramellara G, Renella G. Microbial diversity and soil functions. European Journal of Soil Science, 2003, 54(4): 655-670. DOI:10.1046/j.1351-0754.2003.0556.x |
| [17] |
Roesch L F, Fulthorpe R R, Riva A, Casella G, Hadwin A K, Kent A D, Daroub S H, Camargo F A, Farmerie W G, Triplett E W. Pyrosequencing enumerates and contrasts soil microbial diversity. Isme Journal, 2007, 1(4): 283-290. DOI:10.1038/ismej.2007.53 |
| [18] |
Navarro-Noya Y E, Gómez-Acata S, Montoya-Ciriaco N, Montoya-Ciriaco N, Rojas-Valdez A, Suárez-Arriaga M C, Valenzuela-Encins C, Jiménez-Bueno N, Verhulst N, Govaerts B, Dendooven L. Relative impacts of tillage, residue management and crop-rotation on soil bacterial communities in a semi-arid agroecosystem. Soil Biology & Biochemistry, 2013, 65: 86-95. |
| [19] |
吴凤芝, 王学征. 黄瓜与小麦和大豆轮作对土壤微生物群落物种多样性的影响. 园艺学报, 2007, 34(6): 1543-1546. DOI:10.3321/j.issn:0513-353x.2007.06.033 |
| [20] |
虎德钰.苜蓿后茬不同草田轮作方式对土壤水分和养分的影响[D].宁夏大学, 2014.
|
| [21] |
丁静.不同年限苜蓿草田轮作的作物生长及土壤环境研究[D].西北农林科技大学, 2013.
|
| [22] |
马伦兰, 马培杰, 苏生. 紫花苜蓿-小麦轮作对小麦产量与土壤有机质的影响. 现代农业科技, 2017(18): 1-2. DOI:10.3969/j.issn.1007-5739.2017.18.001 |
| [23] |
鲍士旦.土壤农化分析.北京: 中国农业出版社, 2001.
|
| [24] |
刘玮琦, 茆振川, 杨宇红, 谢丙炎. 应用16S rRNA基因文库技术分析土壤细菌群落的多样性. 微生物学报, 2008, 48(10): 1344-1350. DOI:10.3321/j.issn:0001-6209.2008.10.012 |
| [25] |
王芳, 陈井生, 刘大伟. 不同种植方式大豆根际土壤细菌多样性分析. 作物学报, 2018, 44(10): 1539-1547. |
| [26] |
梁志婷.陇东旱塬不同粮草轮作模式下土壤微生物群落特征及土壤养分的研究[D].兰州大学, 2018.
|
| [27] |
Wen X, Dubinsky E, Yao W U, Yu R, Chen F. Wheat, maize and sunflower cropping systems selectively influence bacteria community structure and diversity in their and succeeding crop's rhizosphere. Journal of Integrative Agriculture, 2016, 15(8): 1892-1902. DOI:10.1016/S2095-3119(15)61147-9 |
| [28] |
Niu J, Chao J, Xiao Y, Xiao Y H, Chen W, Zhang C, Liu X D, Rang Z W, Yin H Q, Dai L J. Insight into the effects of different cropping systems on soil bacterial community and tobacco bacterial wilt rate. Journal of Basic Microbiology, 2017, 57(1): 3-11. |
| [29] |
Sapp M, Harrison M, Hany U, Charlton A, Thwaites R. Comparing the effect of digestate and chemical fertiliser on soil bacteria. Applied Soil Ecology, 2014, 86(86): 1-9. |
| [30] |
Liu J J, Sui Y Y, Yu Z H, Shi Y, Chu H Y, Jin J, Liu X B, Wang G H. High throughput sequencing analysis of biogeographical distribution of bacterial communities in the black soils of northeast China. Soil Biology & Biochemistry, 2014, 70(2): 113-122. |
| [31] |
王伏伟, 王晓波, 李金才, 叶爱华, 王妍, 车威, 朱林. 施肥及秸秆还田对砂姜黑土细菌群落的影响. 中国生态农业学报, 2015, 23(10): 1302-1311. |
| [32] |
Sangwan P, Chen X, Hugenholtz P, Janssen P H. Chthoniobacter flavus gen. nov., sp. nov., the First Pure-Culture Representative of Subdivision Two, Spartobacteria classis nov., of the Phylum Verrucomicrobia. Applied & Environmental Microbiology, 2004, 70(10): 587-5881. |
| [33] |
Togashi J, Ueda K, Namai T. Overwintering of Erwinia carotovora subsp. carotovora in Diseased Tissues in Soil and Its Role as Inoculum for Soft Rot of Chinese Cabbage (Brassica campestris, Pekinensis Group). Journal of General Plant Pathology, 2001, 67(1): 45-50. |
| [34] |
武云鹏, 史凤玉, Hamid M.Imran, 张瑞敬, 朱英波. 野生大豆抗感大豆孢囊线虫材料内生细菌多样性分析. 微生物学报, 2014, 54(8): 926-935. |
| [35] |
姬广海. 溶杆菌属及其在植物病害防治中的研究进展. 云南农业大学学报自然科学, 2011, 26(1): 124-130. |
| [36] |
Im W T, Hu Z Y, Kim K H, Rhee S K, Meng H, Lee S T, Quan Z X. Description of Fimbriimonas ginsengisoli gen. nov., sp. nov. within the Fimbriimonadia class nov., of the phylum Armatimonadetes. Antonie van Leeuwenhoek, 2012, 102(2): 307-317. DOI:10.1007/s10482-012-9739-6 |
| [37] |
许颖, 马德胜, 宋文枫, 魏小芳. 采用16S r DNA高通量测序技术分析油藏微生物多样性. 应用与环境生物学报, 2016, 22(3): 409-414. |
| [38] |
陈重军, 张海芹, 汪瑶琪, 喻徐良, 王建芳, 沈耀良. 基于高通量测序的ABR厌氧氨氧化反应器各隔室细菌群落特征分析. 环境科学, 2016, 37(7): 2652-2658. |
| [39] |
朱英波, 史凤玉, 张瑞敬, 武云鹏. 黑龙江大豆轮作和连作土壤细菌群落多样性比较. 植物保护学报, 2014, 41(4): 403-409. |
| [40] |
李新, 焦燕, 代钢, 杨铭德;温慧洋. 内蒙古河套灌区不同盐碱程度的土壤细菌群落多样性. 中国环境科学, 2016, 36(1): 249-260. DOI:10.3969/j.issn.1000-6923.2016.01.041 |
| [41] |
吴宏亮, 康建宏, 陈阜, 许强, 张海林, 赵亚慧. 不同轮作模式对砂田土壤微生物区系及理化性状的影响. 中国生态农业学报, 2013, 21(6): 674-680. |