 2020, Vol. 40
2020, Vol. 40文章信息
- 周博, 范泽鑫, 杞金华
- ZHOU Bo, FAN Zexin, QI Jinhua
- 哀牢山中山湿性常绿阔叶林木径向生长季节动态及其对气候因子的响应
- Intra-annual radial growth of evergreen and deciduous tree species and their response to climatic factors in a montane moist evergreen broad-leaved forest in the Ailao Mountains, Southwest China
- 生态学报. 2020, 40(5): 1699-1708
- Acta Ecologica Sinica. 2020, 40(5): 1699-1708
- http://dx.doi.org/10.5846/stxb201901070057
-
文章历史
- 收稿日期: 2019-01-07
- 网络出版日期: 2019-12-17





2. 中国科学院大学, 北京 100049;
3. 中国科学院西双版纳热带植物园 哀牢山生态站, 景东 676209
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Ailaoshan Station of Subtropical Forest Ecosystem Studies, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Jingdong 676209, China
常绿阔叶林是我国亚热带气候区的典型植被, 分布范围较广, 森林面积约占我国陆地面积的26%[1], 其作为陆地重要的植被类型, 在固碳增汇、调节气候、保持水土、维持生物多样性等方面具有重要作用[2-3]。随着全球变化加剧, 森林生态系统如何响应气候变化在一定程度上反映了气候变化对陆地生态系统造成的影响。有研究表明气候变暖促进了高纬度高海拔地区树木的生长, 而没有影响或抑制低纬度低海拔地区树木的生长[4-11]。即使在降水没有明显改变的地区, 气温升高亦可能加剧一些植物的干旱胁迫[12-15]。近五十多年来, 云南省大部分地区气温逐渐升高而降水有所减少, 极端干旱事件发生频率增多[16-17]。气候变化如何影响森林生态系统树木生长速率变化, 是当前研究的热点问题。
基于我国热带亚热带地区13个长期森林动态监测样地资料的调查研究, 发现随着温度升高、降水格局改变、空气相对湿度降低以及土壤水分减少, 样地尺度所有个体的平均胸径呈减小趋势, 并且物种组成有所改变, 区域尺度的干旱可能是驱使该转变的重要原因[1]。基于不同气候区2409个森林样地树木径向生长与气候因子的研究表明, 树木生长随着降水量增加而增加, 当降水量增加到1400 mm时生长对降水的响应减弱, 随着温度梯度上升生长先逐渐增加, 当平均温度上升至11℃或最暖月最高温度上升至25—27℃时生长达到峰值, 而温度过低或过高都会限制树木的生长[18]。
不同物候类型树种(如常绿树种, 落叶树种)的径向生长速率和物候存在显著差异[19-20], 从树种的角度研究常绿树种与落叶树种对气候因子的响应有助于进一步了解气候变化对森林动态的影响。哀牢山中山湿性常绿阔叶林位于西南季风气候区的气候过渡地带, 具有雨热同期的气候特点。本研究旨在(1)分析哀牢山中山湿性常绿阔叶林中常绿树种与落叶树种的径向生长季节动态;(2)探讨气候因子对常绿树种与落叶树种径向生长的影响;进而从树种的角度为预测未来气候变化对常绿阔叶林树木生长动态的影响提供借鉴。
1 试验地概况本研究在云南省景东县境内哀牢山自然保护区(24°32′17′′ N, 101°01′45′′ E)进行, 因其地处云南亚热带中部和北部的气候过渡区, 加之其山地的垂直地带性特点, 而具有多样的气候与植被类型。研究地点海拔2507 m, 属亚热带季风气候, 雨热同期, 最热月份7月, 平均气温15.6℃, 最冷月份1月, 平均气温5.7℃, 年均温11℃, 年均降雨量1682 mm, 其中85%的降水集中分布在雨季即5月至10月, 年均相对湿度83%。独特的气候、充沛的降雨和湿润的空气使得山间常笼罩一股云雾[21]。林地内土壤为黄棕壤, 土质疏松孔隙度较高, 地表覆盖的大量凋落物使其具有丰富的土壤有机质和养分, 有机质含量为50—90 mg/g, 氮、磷、钾含量分别为3.5、0.7、3.7 mg/g, 土壤深度可达120—180 cm[21-23]。
哀牢山中山湿性常绿阔叶林作为亚热带常绿阔叶林植被类型, 林内分布着众多壳斗科(Fagaceae)、山茶科(Theaceae)、樟科(Lauraceae)和木兰科(Magnoliaceae)的优势种, 如壳斗科的木果柯(Lithocarpus xylocarpus)、硬壳柯(Lithocarpus hancei)、变色锥(Castanopsis rufescens);樟科的滇润楠(Machilus yunnanensis)、黄心树(Machilus bombycina);山茶科的南洋木荷(Schima noronhae)、折柄茶(Hartia sinensis);木兰科的红色木莲(Manglietia insignis)、多花含笑(Michelia floribunda);此外, 冬青科(Aquifoliaceae)、杜鹃花科(Ericaceae)、山矾科(Symplocaceae)及五加科(Araliaceae)等树种亦较为常见, 其中大部分为常绿树种, 同时散落分布有藏南槭(Acer campbellii)、珍珠花(Lyonia ovalifolia)、吴茱萸五加(Acanthopanax evodiaefolius)等落叶树种[21, 24]。
2 研究方法 2.1 试验对象据中山湿性常绿阔叶林6 hm2样地调查资料, 样地内共分布有25科49属共计68个树种(胸径≥ 1 cm), 其中79.4%为常绿阔叶树种(54种), 落叶阔叶树种仅占19.1%(13种), 及1个针叶树种[25]。
本研究连续观测了哀牢山生态站附近常绿阔叶林中生长良好的4个乔木树种9年的径向生长季节动态, 每个树种选择5个(珍珠花有1个体生长缺失年份较多, 仅分析其中4个体)个体。这些树种均为一般常见种, 其中两种为常绿树种, 厚皮香(Ternstroemia gymnanthera)、南亚枇杷(Eriobotrya bengalensis), 两种为落叶树种, 西桦(Betula alnoides)、珍珠花(Lyonia ovalifolia), 基本信息见下表 1。
| 物种 Species |
科属 Family & Genera |
物候型 Phenology |
重要值 Importance value |
胸径范围 Initial DBH/cm |
树高 Height/m |
年均生长量 Annual growth/ (mm/a) |
| 厚皮香Ternstroemia gymnanthera | 山茶科厚皮香属 | 常绿 | 0.785 | 11.1±2.9 | 4.2±1.0 | 3.2±0.5 |
| 南亚枇杷Eriobotrya bengalensis | 蔷薇科枇杷属 | 常绿 | 3.121 | 13.9±1.9 | 10.1±1.4 | 2.7±0.5 |
| 西桦Betula alnoides | 桦木科桦木属 | 落叶 | 0.017 | 18.0±1.1 | 10.5±1.5 | 10.9±1.6 |
| 珍珠花Lyonia ovalifolia | 杜鹃花科珍珠花属 | 落叶 | 0.773 | 15.1±3.6 | 8.7±3.8 | 10.5±1.3 |
由树木生长环监测树木的径向生长, 安装方法参考Cattelino等提出的生长环安装监测方法, 在研究树种1.3 m胸径处安装金属生长环[26], 如1.3 m处不平整则选其上下平整处安装, 为尽可能减少因树皮水分动态变化引起的膨胀收缩对测量的干扰, 安装前清理安装位置即将脱落的树皮并测量起始胸围, 安装后标记起始刻度线, 随后每个月测量一次树木胸径生长, 目前已累计2009—2017年共计9年的观测数据。
生长观测期间的每月平均气温(Average temperature, ℃)、最高/最低气温(Maximum/Minimum temperature, ℃)、累积降水量(Precipitation, mm)、雨日(降雨天数Precipitation day, days)、雾日(雾日天数Fog day, days)、空气相对湿度(Relative humid, %)及光合有效辐射(Photosynthetically active radiation, PAR, mol m-2 s-1)等气象资料从哀牢山亚热带森林生态系统研究站获得。
2.3 数据处理借助McMahon和Parker (2015)提出的逻辑斯蒂生长模型(Logistic model)拟合树木径向生长进而推算其物候参数[27], 其原理为:分别将各个体每一年12个月累积生长量(dbhdoy)与其观测时间(天, doy)代入(1)中可分别构建该个体的生长模型, 拟合生长曲线的同时计算生长曲线的模型参数(如曲线上下部渐近线、拐点), 基于模型参数再通过(2)式设置对应的参数d(如该年生长量的50%, 对应天数doy即为生长量达全年一半的时间), 即可获取达到已知生长量的时间(如生长起止时间), 而生长曲线拐点对应的生长曲线斜率即最大生长速率, 对应时间即生长速率达到最大的时间, 该过程可根据McMahon和Parker (2015)提供的代码及“MASS”、“chron”、“RColorBrewer” 3个基础包在R Studio软件中完成[27]。
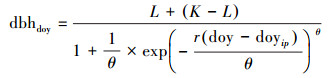
|
(1) |
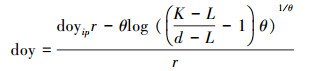
|
(2) |
doy为在一年365天中的天数; dbhdoy为某一时间记录的胸径;doyip为拟合生长曲线中拐点对应的天数; L为生长曲线下部渐近线;K为生长曲线上部渐近线;r为生长曲线拐点的斜率;θ为调整上部渐近线的拟合。
由前后相邻累积生长记录计算各树种年、季节和月净生长量以分析常绿树种和落叶树种生长动态, 其中雨季生长为每年5—10月净生长, 旱季生长则相应为剩余月份即1—4月与11—12月净生长, 采用双因素方差分析比较不同物候类群及不同树种的年际、季节生长差异。从哀牢山亚热带森林生态系统研究站获取年/月气温、降水、雨日、雾日、日照辐射、空气相对湿度等气候资料, 对月气候数据进行去趋势标准化处理, 再结合线性混合效应模型分析气候因子与树木生长的关系, 分别找出与常绿树和落叶树生长相关的影响因子。采用皮尔逊相关性分析气候因子常绿树和落叶树生长时间参数、最大生长速率的相关性, 研究气候变化对树种生长动态的影响, 同时采用单因素方差分析比较常绿树、落叶树生长时间参数和最大生长速率的差异, 由线性回归分析各树种最大生长速率与年生长速率的关系。数据处理和图表绘制在R Studio(R 3.4.1, http://www.R-project.org/)和Excel 2016软件中完成。
3 结果与分析 3.1 常绿和落叶树种年际径向生长动态与物候参数4个树种胸径年净生长量为2.7—10.9 mm/a, 平均年生长量为6.3 mm(表 1)。常绿和落叶树种之间生长速率的差异远大于常绿树种与落叶树种内的差异, 其中落叶树种胸径年净生长量10.6 mm, 显著高于常绿树种(3.0 mm, P < 0.001)。不同树种间年净生长量亦存在显著差异, 其中落叶树西桦最高, 可达10.9 mm;而常绿树南亚枇杷最低, 仅2.7 mm。研究期间, 4个树种的年际径向生长动态有着不同的变化趋势, 且落叶树表现出更大程度的年际变化。到研究后期2013—2017年, 常绿树和落叶树年径向生长呈现出增加趋势(图 1)。

|
| 图 1 哀牢山中山湿性常绿阔叶林常绿和落叶树种径向生长年际动态 Fig. 1 Annual growth of evergreen and deciduous tree species in a montane moist evergreen broad-leaved forest on the Ailao Mountains during 2009—2017 |
4个树种平均在一年中的第105天开始生长, 到第177天达到最大生长速率, 在第258天生长基本停止, 平均生长期长度为153天, 平均最大生长速率0.08 mm/d。在常绿和落叶树间, 常绿树种开始生长的时间(第102天)略早于落叶树种(第108天), 而停止生长的时间晚于落叶树种(第271天与第243天), 使得常绿树具有显著更长的生长期(169天)。尽管落叶树种生长期较短(137天), 但其达到最大生长速率的时间(第174天)却早于常绿树种(第180天), 且可达到的最大生长速率0.14 mm/d远高于常绿树的0.03 mm/d(P < 0.001)(表 2)。线性回归分析表明, 4个树种的最大生长速率与年生长速率成正比, 最大生长速率高的树其年生长速率也高(图 2)。
| 树种 Species |
最大生长速率 Gmax rate |
开始时间# Start day |
最大生长速率时间# Gmax day |
停止时间# Stop day |
持续时间/d Duration |
| 常绿Evergreen | 0.03 | 102 | 180 | 271*** | 169*** |
| 落叶Deciduous | 0.14*** | 108 | 174 | 243 | 137 |
| 厚皮香Ternstroemia gymnanthera | 0.04 | 102 | 177 | 261 | 158 |
| 南亚枇杷Eriobotrya bengalensis | 0.03 | 102 | 183 | 282 | 182 |
| 西桦Betula alnoides | 0.15 | 99 | 169 | 239 | 140 |
| 珍珠花Lyonia ovalifolia | 0.13 | 119 | 181 | 248 | 133 |
| Gmax rate是年内最大生长速率(mm/d);Start day, Gmax day, Stop day, Duration分别是模型拟合出的生长开始时间、最大生长速率时间、停止生长时间以及生长持续时间(天/d);***, P < 0.001;#:指一年中的日序 | |||||

|
| 图 2 常绿阔叶林4树种年生长速率与最大生长速率 Fig. 2 The annual growth rate and the max growth rate for 4 tree species in an evergreen broad-leaved forest |
哀牢山中山湿性常绿阔叶林树种的径向生长主要集中在雨季, 且其生长速率达到最大的时间第169—183天均在雨季的6—7月(约第150—210天, 表 2)。雨季树木胸径平均生长量为5.9 mm, 占全年生长的93%, 而旱季生长量仅占全年生长的7%。在不同物候型中, 落叶树种在雨季的生长量(9.6 mm)远高于常绿树种(2.7 mm)。相比旱季, 雨季生长量对其全年生长量的贡献更大(96%与86%)。常绿树种由于常年展叶而能在旱季条件较适宜时保持生长状态, 使其在旱季生长占全年的比例较落叶树种更高(14%与4%)。在不同树种间, 落叶树种珍珠花在雨季的生长占全年的比例最高为97%, 其次是落叶树种西桦95%, 常绿树种南亚枇杷和厚皮香分别占87%和86%(图 3)。此外, 旱季生长与生长期长度显著正相关(r=0.37, P < 0.001), 因而生长季长度较影响旱季生长。

|
| 图 3 常绿和落叶树种雨季旱季(雨季5—10月)占全年径向生长的百分比 Fig. 3 The percentage of seasonal growth for evergreen and deciduous tree species in a montane moist evergreen broad-leaved forest |
借助线性混合效应模型分析常绿树和落叶树与气候因子间的相关性发现, 低温对于常绿树种和落叶树种的生长速率均有显著影响(P < 0.05表 3), 此外雾日和光合有效辐射也是限制径向生长的重要气候因子。年径向生长量与年气候因子的回归分析显示, 温度相关的气候因子诸如平均气温、最低温、最高温及光合有效辐射对常绿树的生长存在显著影响, 水分相关的气候因子中仅雾日天数对常绿树生长的影响达到了显著水平(P < 0.05), 同时降水量亦存在一定影响(P=0.067), 可见相比于降水日数, 降水量以及平均气温对常绿树生长影响更大;与此不同, 最低温和光合有效辐射对落叶树的年生长有显著影响而平均气温和最高温几乎没有影响, 雾日天数、空气相对湿度、蒸汽压亏缺这类与水分状况相关的气候因子与落叶树的生长显著相关(P < 0.05), 并且与降雨天数也存在一定相关性(P=0.089), 可见降水天数与空气湿度对落叶树的生长更为重要。旱季气候对全年生长也有显著影响, 平均气温、最高温、蒸汽压亏缺等旱季气候因子对常绿树全年生长量具有显著影响, 暗示旱季水分、温度对常绿树生长的限制作用。而旱季气候因子对落叶树种全年生长的影响与年气候因子的影响一致, 表明常绿树年生长对旱季气候因子的响应相比落叶树更为敏感。最后, 由旱季生长与旱季气候因子的分析结果可知, 所有树种在旱季生长均受到旱季最低温的限制。
| 气候因子 Climate factors |
常绿树种Evergreen | 落叶树种Deciduous | ||||||||
| 估计系数 Estimate |
标准差 Std. Error |
t | 显著水平 Pr(>|t|) |
估计系数 Estimate |
标准差 Std. Error |
t | 显著水平 Pr(>|t|) |
|||
| 年生长量 | Temp | 0.087 | 0.034 | 2.537 | 0.013 | 0.000 | 0.062 | 0.002 | 0.998 | |
| Annual growth | Tmin | 0.073 | 0.035 | 2.102 | 0.039 | 0.124 | 0.060 | 2.061 | 0.043 | |
| Tmax | 0.103 | 0.034 | 3.034 | 0.003 | -0.045 | 0.062 | -0.716 | 0.477 | ||
| Prec | 0.065 | 0.035 | 1.856 | 0.067 | 0.094 | 0.062 | 1.519 | 0.134 | ||
| PreDay | 0.050 | 0.035 | 1.399 | 0.166 | 0.106 | 0.061 | 1.727 | 0.089 | ||
| FogDay | 0.073 | 0.035 | 2.074 | 0.041 | 0.138 | 0.060 | 2.279 | 0.026 | ||
| PAR | -0.108 | 0.034 | -3.197 | 0.002 | -0.148 | 0.060 | -2.490 | 0.015 | ||
| RH | 0.056 | 0.035 | 1.597 | 0.114 | 0.144 | 0.060 | 2.390 | 0.020 | ||
| VPD | -0.025 | 0.036 | -0.709 | 0.480 | -0.149 | 0.060 | -2.469 | 0.016 | ||
| 旱季生长量 | Temp | 0.015 | 0.011 | 1.357 | 0.179 | 0.016 | 0.028 | 0.572 | 0.570 | |
| Dry season′s growth | Tmin | 0.039 | 0.010 | 3.790 | 0 | 0.100 | 0.025 | 3.959 | 0 | |
| Tmax | 0.006 | 0.011 | 0.535 | 0.594 | -0.008 | 0.028 | -0.288 | 0.774 | ||
| Prec | 0.017 | 0.011 | 1.566 | 0.121 | 0.048 | 0.027 | 1.754 | 0.084 | ||
| PreDay | 0.019 | 0.011 | 1.788 | 0.078 | 0.050 | 0.027 | 1.817 | 0.074 | ||
| FogDay | 0.039 | 0.010 | 3.844 | 0 | 0.098 | 0.026 | 3.796 | 0 | ||
| PAR | -0.035 | 0.010 | -3.352 | 0.001 | -0.096 | 0.026 | -3.768 | 0 | ||
| RH | 0.033 | 0.011 | 3.096 | 0.003 | 0.078 | 0.027 | 2.920 | 0.005 | ||
| VPD | -0.028 | 0.011 | -2.671 | 0.009 | -0.076 | 0.027 | -2.846 | 0.006 | ||
| Temp为年均气温(Average annual temperature, ℃);Tmin为最低气温(Min temperature, ℃);Tmax为最高气温(Max temperature, ℃);Prec为年累积降水量(Precipitation, mm);PreDay为年累积雨日天数(Precipitation day, days);FogDay为年累积雾日数(Fog day, days);PAR为光和有效辐射(Photosynthetically active radiation, PAR, mol m-2 s-1);RH为空气相对湿度(Relative humid, %);VPD为蒸汽压亏缺(Vapor pressure deficit, VPD, kPa) | ||||||||||
生长参数与气候因子相关分析表明, 气候因子与常绿树种的最大生长速率及生长时间(开始、停止、持续时间)均有显著相关性, 而落叶树种生长时间对气候因子不敏感, 仅最大生长速率与气候因子呈显著相关。降水与常绿树种开始生长的时间显著负相关(P < 0.05), 雾日与常绿树种停止生长的时间正相关(P < 0.1), 降水、雾日与常绿树生长持续时间显著正相关(P < 0.05)。同时常绿树种最大生长速率与平均气温、最高温显著正相关(P < 0.05), 表明气温升高会增加常绿树种在雨季的生长速率。落叶树种最大生长速率与水分如雨日、雾日、空气相对湿度、蒸汽压亏缺显著相关(P < 0.1), 水分状况越好诸如雨日、雾日、空气相对湿度的增加都会增大落叶树的最大生长速率(表 4), 与落叶树种生长跟水分的关系一致(表 3), 印证水分条件如雨日、雾日和空气相对湿度而非温度对落叶树生长的重要性。
| 气候因子 Climate factors |
常绿树种Evergreen | 落叶树种Deciduous | |||||||||
| 开始时间 Start day |
最大生长 速率时间 Gmax day |
停止时间 Stop day |
最大生 长速率 Gmax rate |
持续时间 Duration |
开始时间 Start day |
最大生长 速率时间 Gmax day |
停止时间 Stop day |
最大生 长速率 Gmax rate |
持续时间 Duration |
||
| Temp | -0.09 | -0.05 | -0.04 | 0.22* | 0 | -0.13 | 0.01 | 0.03 | -0.05 | 0.02 | |
| Tmin | -0.02 | -0.03 | 0 | 0.17 | 0.04 | -0.1 | 0.15 | 0.11 | 0.18 | 0.08 | |
| Tmax | -0.08 | 0.09 | 0.04 | 0.26* | -0.04 | -0.13 | -0.08 | -0.02 | -0.1 | -0.01 | |
| Prec | -0.27* | -0.13 | 0.02 | 0.08 | 0.27* | 0.03 | 0.05 | 0.07 | 0.18 | -0.01 | |
| PreDay | -0.14 | -0.09 | 0.06 | 0.06 | 0.19 | 0.07 | 0.03 | 0.05 | 0.23 | -0.02 | |
| FogDay | -0.18 | 0.07 | 0.19 | 0.11 | 0.28* | -0.03 | 0.02 | 0.07 | 0.27* | 0.1 | |
| PAR | 0.13 | -0.02 | -0.14 | -0.24* | -0.21 | 0.08 | -0.07 | -0.08 | -0.26* | -0.1 | |
| RH | -0.12 | -0.01 | 0.13 | 0.08 | 0.23 | -0.04 | 0.02 | 0.08 | 0.26* | 0.12 | |
| VPD | 0.08 | 0.02 | -0.12 | 0 | -0.22 | 0 | -0.05 | -0.1 | -0.27* | -0.12 | |
| *, P < 0.05 | |||||||||||
不同树种的径向生长存在较大差异, 对于常绿和落叶树种的径向生长而言, 落叶树种年生长量(图 1)、雨季生长量与雨季生长量占全年生长的比例(图 3)均显著高于常绿树种, 与在浙江古田山及湖北神农架亚热带常绿阔叶林的研究结果一致[19-20]。树木的径向生长与其自身属性密切相关, 因而常绿与落叶树种径向生长的差异可能与其不同的生理特征有关[19, 28-31]。有研究发现, 常绿树种通常具有较高的防御特性, 如质地较厚且较硬的叶片(部分常绿树具革质或蜡质叶片)、较厚的栅栏组织和细胞壁, 从而可在一定程度上抵御诸如低温、干旱等恶劣环境[30], 因而在部分旱季时间仍可能继续生长, 研究结果显示常绿树种在旱季的生长量可达全年的14%, 而落叶树旱季生长量仅占全年4%。成本收益分析认为植物在构建成本与同化产出之间存在权衡, 常绿树种因在防御上的投入使其具有较大的构建成本, 而常常只有较低的同化能力, 常年维持较低的生长速率[30, 32-34];与此同时, 较高的防御特性也意味着较强的维持能力。与落叶树种相比, 常绿树种虽仅有较低的最大生长速率, 但常绿树种生长开始较早而结束较晚, 从而具有显著更长的生长季(169天, 表 2)。相反地, 落叶树具有较薄且较大的叶片以及较高的同化能力和叶片氮含量[30, 35], 因而具有较大的生长速率(图 1, 表 2);落叶树种在生理上的脆弱性使其对环境变化较为敏感, 当气温较低或水分趋于干旱时落叶以度过不良环境条件, 因而仅有短暂的生长季(137天, 表 2)。相应地, 落叶树种因为每年周期性落叶的特点, 决定了其只能在有限的展叶期尽可能提高生长速率, 同时尽快达到最大生长速率以积累足够多的碳(表 2), 满足当下生长的同时亦维持落叶期间的消耗[28-29, 36]。此外, 落叶树相对较低的叶片构建成本以及落叶前的养分回收减小了落叶带来的损失[30, 37-38]。
4.2 常绿和落叶树种径向生长对气候的响应19世纪末全球尺度的降温趋势结束以后, 气候变化趋势开始逆转[39], 气温逐渐升高[40], 长期降水趋势变化较小而地区降水格局发生改变, 极端降雨事件和干旱事件增多[41-44]。国内热带亚热带地区的气候变化呈现一致的趋势, 近六十年来该地区平均气温每十年上升0.2℃, 平均降水没有明显变化, 无雨日每十年增加13.1天, 空气相对湿度每十年降低11%[1]。分析云南省119个气象站气象观测资料后, 发现过去五十多年以来云南省气候变化趋势与全国乃至全球基本一致, 平均温度每十年升高0.3℃, 相比夏季(0.26℃), 冬季升温更明显(0.33℃)[17]。与分布在东部亚热带地区低海拔的常绿阔叶林不同, 中山湿性常绿阔叶林位于较低纬度但较高海拔(2460 m)地区, 研究期间最低温0.3℃, 自有气象观测(1982年)以来极端低温可低至零下, 部分年份甚至出现冰雪天气。研究结果表明最低温与树木生长显著正相关(表 3), 而研究地点气温变化呈现与云南一致的升温趋势, 研究期间年平均最低温升高, 在此气候变化趋势下未来一段时间气温升高可能促进中山湿性常绿阔叶林树木的生长。
研究表明低温会降低植物根系与土壤微生物活性, 减缓土壤养分的释放进程[45-46], 进而限制根系对土壤养分的吸收, 植物生长速率降低, 加上低温胁迫下植物光合作用强度减弱, 植物在枝条及叶片部分的生物量分配减少, 反过来又影响其光合作用以至生长过程[6, 47-48]。升温则可能缓解了高纬度或高海拔地区低温胁迫对树木生长的影响, 延长了生长季长度[8, 49], 或促进了土壤有机质降解和养分矿化进而提高土壤养分有效性[6, 50-51]。一项基于过去22年55个温带森林样地的胸径资料、土地利用和气候变化的研究表明, 干扰后自然恢复驱动了温带森林生物量增长趋势, 且这一增加趋势与气温变化趋势一致, 在存在土地利用转换历史的地区, 干扰之后的自然恢复可能是促进树木生长主要原因[52]。但哀牢山中山湿性常绿阔叶林为保存完好的原生亚热带常绿阔叶林, 且徐家坝地区早在1981年就已建立自然保护区与生态系统定位研究站进行观测研究, 人为干扰较小[21-22]。
树木的生长受多种环境因子的共同影响, 以及树种本身的生理特征, 使得树木生长对气候变化可能呈现不同的响应[53-55]。对常绿树种与落叶树种而言, 尽管低温和雾日同时影响常绿树与落叶树种的径向生长, 但相比之下常绿树对温度的响应更为敏感(表 3), 可能与其生理特征有关, 常绿树种由于较低的生长速率, 仅需消耗相对较少的水分, 因而可能对水分的依赖较小[31], 气温变化则可能影响常绿树的生理活动以至于生长, 通常常绿树木能借助如气孔调节、渗透调节等机制在一定程度内抵御低温, 但温度过低则直接会抑制常绿树的生理活性从而限制生长[56];落叶树则较可能受水分如雨日、雾日限制(表 3), 在快速生长的时期(比如雨季)落叶树会消耗更多的水分, 因而可能较为依赖环境中的水分条件[31, 56], 在气温较低的旱季, 由于落叶树以休眠的状态度过其中大部分时间而可能影响较小。除升温对树木生长的正效应外, 极端气候事件如干旱、极端高温、极端低温都可能对树木的生长造成较大干扰[16-17]。
5 结论常绿树木生长季较长而落叶树木生长速率较高, 低温是限制中山湿性常绿阔叶林树木径向生长的主要气候因子, 相比之下常绿树对温度的响应更为敏感, 而落叶树可能受水分条件限制, 且常绿树生长时间(开始、停止、持续时间)与气候因子显著相关而落叶树生长时间对气候因子不敏感;气候变化因而可能促进常绿阔叶林树木生长的同时改变不同物候类型树种在常绿阔叶林中的生长状态与分布格局。由于生长变异与气候波动造成的不稳定性, 今后应开展基于树木生长与气象的长期观测并结合树木年轮与功能性状研究常绿阔叶林对气候变化的响应。
致谢: 感谢中国科学院哀牢山亚热带森林生态系统研究站、中国生态系统研究网络提供气象数据资料。| [1] |
Zhou G Y, Houlton B Z, Wang W T, Huang W J, Xiao Y, Zhang Q M, Liu S Z, Cao M, Wang X H, Wang S L, Zhang Y P, Yan J H, Liu J X, Tang X L, Zhang D Q. Substantial reorganization of China's tropical and subtropical forests:Based on the permanent plots. Global Change Biology, 2014, 20(1): 240-250. DOI:10.1111/gcb.12385 |
| [2] |
Dixon R K, Solomon A M, Brown S, Houghton R A, Trexier M C, Wisniewski J. Carbon pools and flux of global forest ecosystems. Science, 1994, 263(5144): 185-190. DOI:10.1126/science.263.5144.185 |
| [3] |
Pan Y, Birdsey R A, Fang J Y, Houghton R, Kauppi P E, Kurz W A, Phillips O L, Shvidenko A, Lewis S L, Canadell J G, Ciais P, Jackson R B, Pacala S W, McGuire A D, Piao S L, Rautiainen A, Sitch S, Hayes D. A large and persistent carbon sink in the world's forests. Science, 2011, 333(6045): 988-993. DOI:10.1126/science.1201609 |
| [4] |
Black T A, Chen W J, Barr A G, Arain M A, Chen Z, Nesic Z, Hogg E H, Neumann H H, Yang P C. Increased carbon sequestration by a boreal deciduous forest in years with a warm spring. Geophysical Research Letters, 2000, 27(9): 1271-1274. DOI:10.1029/1999GL011234 |
| [5] |
Bogaert J, Zhou L, Tucker C J, Myneni R B, Ceulemans R. Evidence for a persistent and extensive greening trend in Eurasia inferred from satellite vegetation index data. Journal of Geophysical Research:Atmospheres, 2002, 107(D11): 4119. |
| [6] |
Lie G W, Xue L. Biomass allocation patterns in forests growing different climatic zones of China. Trees, 2016, 30(3): 639-646. DOI:10.1007/s00468-015-1306-0 |
| [7] |
Luo D W, Huang J G, Jiang X Y, Ma Q Q, Liang H X, Guo X L, Zhang S K. Effect of climate and competition on radial growth of Pinus massoniana and Schima superba in China's subtropical monsoon mixed forest. Dendrochronologia, 2017, 46: 24-34. DOI:10.1016/j.dendro.2017.08.001 |
| [8] |
Rustad L, Campbell J, Marion G, Norby R, Mitchell M, Hartley A, Cornelissen J, Gurevitch J, GCTE-NEWS. A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia, 2001, 126(4): 543-562. DOI:10.1007/s004420000544 |
| [9] |
Sebastià M T. Plant guilds drive biomass response to global warming and water availability in subalpine grassland. Journal of Applied Ecology, 2007, 44(1): 158-167. |
| [10] |
Zhang W T, Jiang Y, Dong M Y, Kang M Y, Yang H C. Relationship between the radial growth of Picea meyeri and climate along elevations of the Luyashan mountain in North-central China. Forest Ecology and Management, 2012, 265: 142-149. DOI:10.1016/j.foreco.2011.10.017 |
| [11] |
Zhao C Z, Liu Q. Growth and physiological responses of Picea asperata seedlings to elevated temperature and to nitrogen fertilization. Acta Physiologiae Plantarum, 2009, 31(1): 163-173. DOI:10.1007/s11738-008-0217-8 |
| [12] |
Barber V A, Juday G P, Finney B P. Reduced growth of Alaskan white Spruce in the twentieth century from temperature-induced drought stress. Nature, 2000, 405(6787): 668-673. DOI:10.1038/35015049 |
| [13] |
Lévesque M, Siegwolf R, Saurer M, Eilmann B, Rigling A. Increased water-use efficiency does not lead to enhanced tree growth under xeric and mesic conditions. New Phytoligist, 2014, 203(1): 94-109. DOI:10.1111/nph.12772 |
| [14] |
Peñuelas J, Canadell J G, Ogaya R. Increased water-use efficiency during the 20th century did not translate into enhanced tree growth. Global Ecology and Biogeography, 2011, 20(4): 597-608. DOI:10.1111/j.1466-8238.2010.00608.x |
| [15] |
Liu H Y, Park Williams A, Allen C D, Guo D L, Wu X C, Anenkhonov O A, Liang E Y, Sandanov D V, Yin Y, Qi Z H, Badmaeva N K. Rapid warming accelerates tree growth decline in semi-arid forests of Inner Asia. Global Change Biology, 2013, 19(8): 2500-2510. DOI:10.1111/gcb.12217 |
| [16] |
Fan Z X, Bräuning A, Cao K F, Zhu S D. Growth-climate responses of high-elevation conifers in the central Hengduan Mountains, Southwestern China. Forest Ecology and Management, 2009, 258(3): 306-313. DOI:10.1016/j.foreco.2009.04.017 |
| [17] |
Fan Z X, Bräuning A, Thomas A, Li J B, Cao K F. Spatial and temporal temperature trends on the Yunnan Plateau (Southwest China) during 1961-2004. International Journal of Climatology, 2011, 31(14): 2078-2090. DOI:10.1002/joc.2214 |
| [18] |
Bowman D M J S, Williamson G J, Keenan R J, Prior L D. A warmer world will reduce tree growth in evergreen broadleaf forests:evidence from Australian temperate and subtropical eucalypt forests. Global Ecology and Biogeography, 2014, 23(8): 925-934. DOI:10.1111/geb.12171 |
| [19] |
Chi X L, Tang Z Y, Xie Z Q, Guo Q, Zhang M, Ge J L, Xiong G M, Fang J Y. Effects of size, neighbors, and site condition on tree growth in a subtropical evergreen and deciduous broad-leaved mixed forest, China. Ecology and Evolution, 2015, 5(22): 5149-5161. DOI:10.1002/ece3.1665 |
| [20] |
Chi X L, Guo Q, Fang J Y, Schmid B, Tang Z Y. Seasonal characteristics and determinants of tree growth in a Chinese subtropical forest. Journal of Plant Ecology, 2017, 10(1): 4-12. |
| [21] |
邱学忠. 哀牢山森林生态系统研究. 昆明:云南科技出版社, 1998, 1-15. |
| [22] |
朱华, 闫丽春. 云南哀牢山种子植物. 昆明:云南科技出版社, 2009, 3-6. |
| [23] |
白雪, 范泽鑫. 哀牢山中山湿性常绿阔叶林水青树年轮宽度对气候变化的响应. 林业科学, 2018, 54(3): 161-167. |
| [24] |
朱华. 云南中山湿性常绿阔叶林起源的探讨. 植物科学学报, 2016, 34(5): 715-723. |
| [25] |
巩合德, 杨国平, 鲁志云, 刘玉洪. 哀牢山常绿阔叶林树种多样性及空间分布格局. 生物多样性, 2011, 19(2): 143-150. |
| [26] |
Cattelino P J, Becker C A, Fuller L G. Construction and installation of homemade dendrometer bands. Northern Journal of Applied Forestry, 1986, 3(2): 73-75. DOI:10.1093/njaf/3.2.73 |
| [27] |
McMahon S M, Parker G G. A general model of intra-annual tree growth using dendrometer bands. Ecology and Evolution, 2015, 5(2): 243-254. DOI:10.1002/ece3.1117 |
| [28] |
Antúnez I, Retamosa E C, Villar R. Relative growth rate in phylogenetically related deciduous and evergreen woody species. Oecologia, 2001, 128(2): 172-180. DOI:10.1007/s004420100645 |
| [29] |
Chaturvedi R K, Raghubanshi A S, Singh J S. Leaf attributes and tree growth in a tropical dry forest. Journal of Vegetation Science, 2011, 22(5): 917-931. DOI:10.1111/j.1654-1103.2011.01299.x |
| [30] |
Eamus D. Ecophysiological traits of deciduous and evergreen woody species in the seasonally dry tropics. Trends in Ecology & Evolution, 1999, 14(1): 11-16. |
| [31] |
白坤栋, 蒋得斌, 曹坤芳, 万贤崇, 廖德宝. 哀牢山和猫儿山中山常绿和落叶阔叶树光合特性对季节温度变化的响应. 生态学报, 2010, 30(4): 905-913. |
| [32] |
Kikuzawa K. A cost-benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern. The American Naturalist, 1991, 138(5): 1250-1263. DOI:10.1086/285281 |
| [33] |
Kikuzawa K, Lechowicz M J. Theories of leaf longevity//Kikuzawa K, Lechowicz M J, eds. Ecology of Leaf Longevity. Tokyo: Springer, 2011: 41-56.
|
| [34] |
Sobrado M A. Cost-benefit relationships in deciduous and evergreen leaves of tropical dry forest species. Functional Ecology, 1991, 5(5): 608-616. DOI:10.2307/2389479 |
| [35] |
Poorter L, Bongers F. Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology, 2006, 87(7): 1733-1743. DOI:10.1890/0012-9658(2006)87[1733:LTAGPO]2.0.CO;2 |
| [36] |
Poorter L, Wright S J, Paz H, Ackerly D D, Condit R, Ibarra-Manríquez G, Harms K E, Licona J C, Martínez-Ramos M, Mazer S J, Muller-Landau H C, Peña-Claros M, Webb C O, Wright I J. Are functional traits good predictors of demographic rates? Evidence from five neotropical forests. Ecology, 2008, 89(7): 1908-1920. DOI:10.1890/07-0207.1 |
| [37] |
Huang J J, Wang X H, Yan E R. Leaf nutrient concentration, nutrient resorption and litter decomposition in an evergreen broad-leaved forest in Eastern China. Forest Ecology and Management, 2007, 239(1/3): 150-158. |
| [38] |
宁秋蕊, 李守中, 姜良超, 陶晶晶, 陈涵睿, 刘聪, 杨贤宇. 植物叶片养分再吸收特征及其影响因子. 应用与环境生物学报, 2017, 23(5): 811-817. |
| [39] |
PAGES 2k Consortium. Continental-scale temperature variability during the past two millennia. Nature Geoscience, 2013, 6(5): 339-346. DOI:10.1038/ngeo1797 |
| [40] |
IPCC. Climate Change 2013: The Physical Science Basis. The Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2013: 3-32.
|
| [41] |
Adler R F, Gu G J, Sapiano M, Wang J J, Huffman G J. Global precipitation:means, variations and trends during the satellite era (1979-2014). Surveys in Geophysics, 2017, 38(4): 679-699. DOI:10.1007/s10712-017-9416-4 |
| [42] |
Potter N J, Chiew F H S, Frost A J. An assessment of the severity of recent reductions in rainfall and runoff in the murray-darling basin. Journal of Hydrology, 2010, 381(1/2): 52-64. |
| [43] |
Verdon-Kidd D C, Kiem A S. Nature and causes of protracted droughts in Southeast Australia:Comparison between the federation, WWII, and big dry droughts. Geophysical Research Letters, 2009, 36(22): L22707. DOI:10.1029/2009GL041067 |
| [44] |
Sun F B, Roderick M L, Farquhar G D. Rainfall statistics, stationarity, and climate change. Proceedings of the National Academy of Sciences of the United States of America, 2018, 115(10): 2305-2310. DOI:10.1073/pnas.1705349115 |
| [45] |
Aerts R, Chapin III F S. The mineral nutrition of wild plants revisited:a re-evaluation of processes and patterns. Advances in Ecological Research, 1999, 30: 1-67. DOI:10.1016/S0065-2504(08)60016-1 |
| [46] |
Hobbie S E, Schimel J P, Trumbore S E, Randerson J R. Controls over carbon storage and turnover in high-latitude soils. Global Change Biology, 2000, 6(S1): 196-210. DOI:10.1046/j.1365-2486.2000.06021.x |
| [47] |
Lambers H, Chapin III F S, Pons T L. Plant Physiological Ecology. 2nd ed. New York: Springer, 2008.
|
| [48] |
Reich P B, Luo Y J, Bradford J B, Poorter H, Perry C H, Oleksyn J. Temperature drives global patterns in forest biomass distribution in leaves, stems, and roots. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(38): 13721-13726. DOI:10.1073/pnas.1216053111 |
| [49] |
Jarvis P, Linder S. Constraints to growth of boreal forests. Nature, 2000, 405(6789): 904-905. DOI:10.1038/35016154 |
| [50] |
Lin D L, Xia J Y, Wan S Q. Climate warming and biomass accumulation of terrestrial plants:a meta-analysis. New Phytologist, 2010, 188(1): 187-198. DOI:10.1111/j.1469-8137.2010.03347.x |
| [51] |
Miller A E, Schimel J P, Sickman J O, Meixner T, Doyle A P, Melack J M. Mineralization responses at near-zero temperatures in three Alpine soils. Biogeochemistry, 2007, 84(3): 233-245. DOI:10.1007/s10533-007-9112-4 |
| [52] |
McMahon S M, Parker G G, Miller D R. Evidence for a recent increase in forest growth. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(8): 3611-3615. DOI:10.1073/pnas.0912376107 |
| [53] |
Allen C D, MaCalady A K, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears D D, Hogg E H, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J H, Allard G, Running S W, Semerci A, Cobb N. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest Ecology and Management, 2010, 259(4): 660-684. DOI:10.1016/j.foreco.2009.09.001 |
| [54] |
Anderegg W R, Schwalm C, Biondi F, Camarero J J, Koch G, Litvak M, Ogle K, Shaw J D, Shevliakova E, Williams A P, Wolf A, Ziaco E, Pacala S. Forest ecology. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science, 2015, 349(6247): 528-532.
|
| [55] |
Dulamsuren C, Hauck M, Leuschner C. Recent drought stress leads to growth reductions in Larix sibirica in the Western Khentey, Mongolia. Global Change Biology, 2010, 16(11): 3024-3035. |
| [56] |
刘金玉, 付培立, 王玉杰, 曹坤芳. 热带喀斯特森林常绿和落叶榕树的水力特征和水分关系与抗旱策略. 植物科学学报, 2012, 30(5): 484-493. |