 2020, Vol. 40
2020, Vol. 40文章信息
- 徐敏, 边红枫, 徐丽, 陈智, 何念鹏
- XU Min, BIAN Hongfeng, XU Li, CHEN Zhi, HE Nianpeng
- 脉冲式降水对不同类型草地土壤微生物呼吸碳释放量的影响
- Effects of precipitation pulse on soil carbon released by microbes in different grasslands
- 生态学报. 2020, 40(5): 1562-1571
- Acta Ecologica Sinica. 2020, 40(5): 1562-1571
- http://dx.doi.org/10.5846/stxb201901040032
-
文章历史
- 收稿日期: 2019-01-04
- 网络出版日期: 2019-12-17

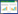




2. 中国科学院生态系统网络观测与模拟重点实验室, 北京 100101;
3. 中国科学院大学资源与环境学院, 北京 100049
2. Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China;
3. College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100049, China
降水事件是陆地生态系统生物地球化学循环的重要驱动因素, 会引起土壤水分的快速变化, 常使生态系统生物地球化学循环在短期内呈现出非常规现象。其中, 降水事件引起土壤在短期内释放大量CO2过程常被称为降水脉冲效应[1];在干旱半干旱区, 降水频率较低, 一次降水事件就会引起土壤有机质快速分解, 加快物质的循环, 因此这种快速的脉冲过程对于土壤养分循环和生物养分利用都具有重要意义[2-3]。当前, 在全球变化日益加剧的背景下, 已有的气候变化模型预测未来降雨模式将会从强度和频度两方面发生改变, 其中干旱半干旱地区可能会经历更长时间的干旱且降水频率也更难准确预测[4-5], 而降雨模式的变化也会对降水脉冲效应产生影响, 进而影响陆地生态系统碳循环。因此, 探讨降水频率变化、干旱程度增加等对降水脉冲效应的影响;以及分析降雨强度、降雨事件持续时间、土壤干旱历史和水分状况等因素的影响[6-7], 对全球碳循环研究具有重要意义。
降水事件通过快速提升土壤水分含量, 使土壤微生物呼吸显著增强。自然界中, 干旱程度、降水强度和降水频度的差异使得微生物经历不同的土壤水分变化过程, 形成了一套适合自身生存和发展的适应机制[8-10]。干旱时, 受土壤水分和基质扩散的限制, 土壤微生物采取休眠或部分死亡策略应对干旱胁迫, 微生物对有机质的利用减少;然而, 降水事件发生后, 土壤水分充足且可利用基质丰富, 土壤微生物充分利用这短暂的有利时机, 快速进行生理代谢和生长繁殖活动[11]。土壤先前干旱程度(或长期气候特征)将影响土壤微生物种类组成、多样性、生物量以及能量分配, 因而不同干旱区域(或干旱程度不同的区域)的土壤微生物可能会对降水事件产生不同响应[12], 但是, 目前人们对这种普遍存在的自然过程及其机制尚不清楚。此外, 降水强度与持续时间也会影响土壤微生物的响应, 有研究显示当降雨量适中时土壤CO2释放速率将明显增加, 大型降雨可使土壤CO2释放总量增加几倍到几百倍[13]。小降雨事件通常只能激发表层土壤微生物的分解过程, 引起表层土壤CO2大量释放[14-15], 但表层水分蒸发较快, 因此引起的土壤分解过程非常短暂;然而, 在干旱与半干旱地区, 5 mm或小于5 mm小降水事件发生频率非常高, 对总降雨的累积贡献超过70%;因此小降水事件的脉冲效应引起CO2排放可能占土壤碳释放总量的很大一部分, 并具有重要的生态意义[16]。
在已有研究中, 重湿后土壤微生物呼吸速率显著增高[17-19], 甚至可达到降雨前的几倍到几十倍[20-21];相对于持续湿润土壤, 降水脉冲后土壤微生物的呼吸速率明显高于降水前的土壤微生物呼吸速率[20]。土壤微生物呼吸速率与土壤CO2累积释放量是研究脉冲效应的重要指标。微生物对降水的响应是一个剧烈并且快速的过程, 呼吸速率可以在数十分钟内达到最大值[20, 22];然而, 在传统研究中, 土壤微生物呼吸速率的测定方法多为碱液吸收法或气相色谱法, 测量时间间隔多为小时、天[23-25], 难以捕捉到降水后短时间内的微生物动态变化过程。目前, 有关降水变化对土壤释放CO2的长期影响已开展了大量的研究工作[8, 26-27], 但对于快速释放CO2过程的研究却非常少、并且深入的机理性分析也十分匮乏。
降水事件后土壤释放的CO2可能来源于干旱时期土壤中积累的非生物有机碳和微生物生物量碳;前人研究表明, 重湿后土壤微生物呼吸的脉冲效应主要是由其细胞质溶质释放引起的[28-29]。然而, 也有研究提出干湿循环过程中释放CO2量大于土壤微生物生物量, 表明土壤释放CO2可能有其他来源。在降水事件发生后, 土壤团聚体结构遭到破坏, 使得原本受物理保护的非生物有机质通过水分运输作用成为微生物可利用的底物[30-31]。目前, 部分研究认为短期土壤呼吸速率提高主要是因为非生物底物供应的增加[32], 也有研究认为土壤进行连续的干燥-再湿循环后, 土壤碳矿化主要来源于微生物生物量碳[33];然而, 这两种来源对于土壤微生物呼吸脉冲的贡献尚不清楚。如果非生物有机碳是CO2脉冲效应的主要来源, 那么长期干燥再湿循环必定会减少土壤碳贮量;反之, 如果生物量碳是脉冲效应的主要贡献, 那么脉冲效应将会增加微生物的生理压力, 长时间范围内将减少碳矿化, 增加土壤碳固存[20];因此, 探究微生物呼吸脉冲的机制对于预测未来土壤碳循环非常重要。
鉴于干旱、半干旱地区降水脉冲效应的重要性, 本研究采用自主研发的能够快速、连续高频测定土壤呼吸速率的装置, 探讨内蒙古3种典型草地类型(草甸草原、典型草原和荒漠草原)在土壤复湿后土壤微生物CO2释放的快速响应过程。在此基础上, 我们提出实验假设:(1)草地土壤对突发降水会产生强烈快速的响应, 且不同草地类型土壤对相同强度降水响应强度不同;(2)降水脉冲效应与土壤长期经历的干旱状况有关, 越干旱地区土壤呼吸对降水脉冲的响应越强烈。
1 研究区概况内蒙古地处欧亚大陆内部, 地域辽阔, 气候类型从东北到西南依次为东北半湿润区、中部半干旱区和西部干旱气候区。在不同气候条件下, 草地植被类型形成明显的经度地带性, 从东到西由草甸草原向典型草原、荒漠草原过渡。草甸草原位于草原向山地针叶林过渡区, 土壤类型以黑钙土为主;多年生旱生、中旱生植物群落占优势, 主要建群种有狼针草(Stipa baicalensis)和羊草(Leymus chinensis)等。典型草原分布在内蒙古高原中部, 土壤主要有暗栗钙土和典型栗钙土;植被群落主要由旱生性多年生草本植物组成, 主要建群种包括大针茅(Stipa grandis)和羊草(Leymus chinensis)等。荒漠草原分布在内蒙古西部, 土壤主要是沙砾质荒漠土、灰漠土和灰钙土;植被群落主要由旱生性更强的多年生矮小草本植物组成, 其主要建群种为小针茅(Stipa klemenzii)和沙生针茅(Stipa glareosa)等。本研究沿内蒙古草原的东北草地样带, 由东向西以草甸草原、典型草原和荒漠草原为研究对象, 样地基本信息及采样点分布详见表 1和图 1。

|
| 图 1 研究区采样点分布 Fig. 1 Location of the study site |
| 草地类型 Grassland types |
海拔 Altitude/m |
经度 Longitude/E |
纬度 Latitude/N |
年降水量 MAP/mm |
年均温 MAT/℃ |
土壤理化性质 Soil physicochemical properties |
|||
| pH | OPR/mv | 砂粒/% | SOC/% | ||||||
| 草甸草原Meadow grassland | 269 | 44.52 | 121.04 | 320.32 | 5.8 | 8.01a(0.11) | 215 (8.96) | 76.26(1.48) | 0.84(0.18) |
| 典型草原Typical grassland | 1129 | 44.26 | 116.52 | 262.91 | 1.17 | 7.55(0.15) | 219 (7.55) | 61.64(3.00) | 1.64(0.20) |
| 荒漠草原Desert grassland | 955 | 43.63 | 112.15 | 140.25 | 3.69 | 8.24(0.16) | 203 (8.23) | 85.19(4.47) | 0.33(0.03) |
| MAP:年平均降水量Mean annual precipitation; MAT:年平均温度Mean annual temperature; OPR:氧化还原电位Oxidation-Reduction Potential; SOC:土壤有机碳Soil organic carbon; a数据以平均±标准差表示(n=5:括号内为标准差) | |||||||||
土壤样品采集于2018年7月。对各个草地类型, 我们选择地势平坦区域设置1条100 m调查带, 间隔20 m左右设置了4个1 m × 1 m植物群落调查样方;在去除表层凋落物后, 每个样方用土钻依次钻取0—10 cm的表层土5 kg。所有土壤样品带回实验室, 手工挑除可见根系, 过2 mm筛[34], 在阴凉通风处风干, 用于理化性质分析及土壤呼吸测定实验。
2.2 室内测定及实验设计土壤含水量及土壤饱和含水量使用烘干法测定[34-35], 即每种土壤随机设4个重复, 称取10 g风干土于滤网中, 将样品在蒸馏水中浸泡4 h左右, 然后将样品静置, 漏下多余水分使土壤饱和, 称重后将样品放在105 ℃烘箱中烘12 h以上, 直至重量不再发生变化, 取出后称重, 根据浸泡和烘干前后的重量来计算土壤饱和含水量及土壤含水量。用pH计测量土壤pH, 氧化还原电位(OPR), 土壤和蒸馏水的混合比例为1 : 2.5, 然后在振荡器上振荡2 h后静置, 再取上清液测定。土壤粒径采用激光粒度仪测定, 在使用30%过氧化氢和30%盐酸去除有机物和碳酸盐后, 然后上机测试。土壤有机碳采用容量分析法测定。
风干后的土壤样品用于测定土壤呼吸速率, 调节温度为20 ℃, 以适宜微生物的生长, 并选择60%饱和含水量作为土壤初始含水量。每类土壤称取30 g置于聚乙烯塑料瓶内(直径5 cm, 高10 cm), 设置4个重复, 土壤复湿之前先测干土的呼吸速率, 然后用称重法给土壤加水至60%饱和含水量, 测定加水后48 h内土壤呼吸速率;此外, 在再湿后的1、2、4、8、12、24、26、28、30、32、34、36 h和48 h采用称重法分别测量土壤含水量, 记录土壤含水量变化。
由于土壤微生物对降水脉冲事件的响应通常在几分钟内发生, 因此采用了可以快速、高频率测定土壤微生物呼吸速率的装置[35]。该装置主要包括气体进样装置、CO2起始浓度调节装置、自动控制装置和红外CO2浓度分析仪器;主要仪器Licor-7000红外CO2浓度分析仪、PRI-8800土壤培养前处理装置、恒温水浴锅和数据采集器等[34-35]。我们可利用该装置测定所有重复样品在48 h内每间隔10 min的土壤微生物呼吸速率(RSoil);在具体操作过程中, 单位土壤微生物呼吸速率通过CO2浓度变化的斜率进行计算, 计算公式如下:
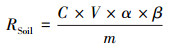
|
(1) |
式中, RSoil为单位土壤微生物呼吸速率(μg C g-1 soil h-1), C为测试时间内CO2浓度变化斜率, V是培养瓶和管线的总体积, m是培养瓶内土壤干重(g), α是CO2气体质量转化系数(由CO2到C), β是时间转化系数(从秒到小时)。
此外, 为了排除土壤有机质含量差异引起的误差, 计算了单位有机质微生物呼吸速率。
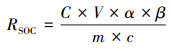
|
(2) |
式中, RSOC是单位有机质微生物呼吸速率(mg C g-1 SOC h-1), c是土壤有机碳含量百分比(%)。
2.3 降水脉冲效应的表征为了更加深刻的描述降水脉冲效应的强度及变化过程, 提出用3个参数来刻画降水脉冲事件中微生物呼吸速率的变化(图 2):RSOC-max:微生物最大呼吸速率(mg C g-1 SOC h-1);脉冲效应持续时间:突发降雨情况下, 微生物呼吸速率维持在1/3最大土壤微生物呼吸速率之内的时间(h);TRSOC-max:达到最大微生物呼吸速率所用时间(h)。另外,这个过程中土壤含水量逐渐下降(图 2)。

|
| 图 2 降水脉冲效应过程特征值及土壤含水量变化 Fig. 2 Characteristic of precipitation pulse effect and change in water content |
此外,还计算了48h内土壤微生物呼吸碳累积释放量, 根据土壤质量, 首先计算单位土壤微生物呼吸累积释放量ARSoil, 具体公式如下:
|
(3) |
ARSoil是单位土壤微生物呼吸累计释放量(μg C g-1 soil);ti和ti+1是前后测定的时间(h)。
由于3种草原土壤有机碳含量存在显著性差异, 为了排除有机碳含量差异的影响, 我们利用土壤有机碳含量将ARSoil标准化为ARSOC, 从而准确定量降水脉冲事件对单位有机质土壤微生物呼吸的影响。计算公式如下:
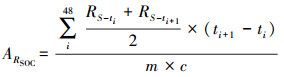
|
(4) |
ARSOC是单位有机质微生物呼吸累积释放量(mg C g-1 SOC);c是土壤有机碳含量百分比(%)。
2.4 统计分析利用IBM SPSS Statistics 19.0统计软件对实验土样的理化性质、微生物呼吸速率, 微生物累积呼吸量以及脉冲效应的特征值进行单因素方差分析, 所有检验显著性水平P=0.05。采用Sigmaplot 10.0软件及EXCEL分析软件对微生物呼吸速率和累积分解量在48 h内的变化作图。
3 结果分析 3.1 降水事件对草地土壤碳释放的脉冲效应由土壤在20 min内的单位有机质呼吸速率变化可得(图 3),土壤复湿后,RSOC显著提高, 在3 min内微生物呼吸开始出现响应, 并且这种响应过程强烈且快速(为了直观地描述加水前后速率的变化,以加水时间作为起始时间0 min,-10 min为加水前测量干土呼吸速率的时刻), 在这个过程中单位有机质微生物呼吸速率快速上升, 在10 min内达到了最大值,达到最大值时间不存在显著性差异。在复湿之前3种土壤的呼吸速率均较低, 降水后3种草原土壤的RSOC-max可达到干土呼吸速率的5—15倍。从图 3中48h内微生物呼吸变化可知, 单位有机质微生物呼吸速率达到峰值后呈指数下降, 在整个试验期间的后12h内,土壤微生物呼吸速率保持稳定波动,在此过程中单位有机质呼吸速率保持为荒漠草原最高,草甸草原次之,典型草原最低。48 h内, 3种草地类型的土壤呼吸速率下降到背景值的1—2倍。由图 3中土壤含水量/土壤饱和含水量的变化可知土壤含水量/土壤饱和含水量呈线性下降, 变化幅度荒漠草原>草甸草原>典型草原。

|
| 图 3 脉冲降水后20 min和48 h内单位有机质土壤微生物呼吸速率的变化以及48 h内土壤含水量/饱和含水量的变化 Fig. 3 Changes in soil microbial respiration rate per unit SOC within 20 min and 48 h after pulsed precipitation and changes in soil water content / saturated water content within 48 h |
三种土壤的降水脉冲效应特征不同,对脉冲效应的特征值比较分析发现单位有机质微生物最大呼吸速率及降水脉冲效应持续时间在三种草地类型间均存在显著性差异(图 4),其中单位有机质微生物最大呼吸速率表现为, 荒漠草原最大((1.58 ± 0.04) mg C g-1 SOC h-1), 草甸草原次之((0.73 ± 0.05) mg C g-1 SOC h-1), 典型草原最小((0.50 ± 0.03) mg C g-1 SOC h-1);3种草原土壤的脉冲效应持续时间则表现出相反的结果, 典型草原(2.5 h) >草甸草原(1.5 h) >荒漠草原(0.67 h)。显著性均为P < 0.05。

|
| 图 4 三种草地类型降雨脉冲过程特征值(单位有机质微生物最大呼吸速率及降水脉冲效应持续时间)比较 Fig. 4 Characteristic values of precipitation pulses of three grassland types (maximum microbial respiration rate per unit SOC and duration of precipitation pulse effect) 不同字母表示存在显著性差异(P < 0.05) |
土壤复湿48 h内, 3种草地单位土壤与单位有机质微生物呼吸碳累积量均存在差异,其中,单位土壤微生物呼吸碳累积量表现为典型草原最大(55.58 μg C g-1 soil), 荒漠草原最小(32.22 μg C g-1 soil), 草甸草原与典型草原不存在显著差异, ARSoil与土壤pH有强烈的负相关关系, 与土壤氧化还原电位(OPR)呈正相关关系。有趣的是, 将ARSoil用土壤有机碳含量标准化后, 发现ARSOC大小关系为荒漠草原(9.72 mg C g-1 SOC) >草甸草原(6.54 mg C g-1 SOC) >典型草原(3.54 mg C g-1 SOC)(图 5);这与土壤有机碳含量呈反比, 并且荒漠草原ARSOC远高于典型草原和草甸草原。ARSOC存在显著性差异(P < 0.05)。根据2017—2018年降水事件统计, 草甸草原, 典型草原和荒漠草原的降雨频次分别为52、82、34次, 降雨频率与ARSoil成正相关, 而与ARSOC成负相关。

|
| 图 5 脉冲降水后48h内三种草地类型的单位有机质微生物呼吸碳累积量变化 Fig. 5 Change in microbial respiration carbon accumulation per unit SOC of three grassland types within 48 h after pulsed precipitation |
土壤复湿后, 土壤CO2释放速率显著增加, 在10 min内观察到了3种草地类型的微生物呼吸峰值, 且降水脉冲后的微生物呼吸速率是降水前的5—15倍, 这与先前部分研究报道结果相似[32, 36]。Fierer和Schimel[20]的研究表明:土壤再湿后其呼吸速率是干土测量值的4.7—5.75倍, 有的研究发现森林土壤在模拟降水后的呼吸速率可达到干土10—100倍[32]。同时, 随着时间的推移, 可利用基质不断被消耗, 水分也快速的蒸发散失, 微生物呼吸速率呈指数下降, 在12 h之后保持平稳的波动, 并在48 h内恢复到了微生物呼吸的背景水平, 与Sponseler[21]的发现相一致。土壤微生物呼吸对脉冲降水的这种快速响应来源于干燥时期土壤中稳定积累的有机质, 水分限制使得溶质扩散率降低, 微生物摄取的有机质的减少, 再加上持续的胞外酶活性, 导致土壤中可溶性有机物的增加, 因此干旱促进了土壤中有机碳库含量的增加。而当长期干旱的土壤经历降水刺激后, 且土壤水分含量超过土壤微生物呼吸的干旱阈值, 土壤中库存底物的扩散性也不断增强, 并成为微生物可用的基质同时, 由于微生物的脉冲效应迅速且短暂, 在数分钟内呼吸速率快速增长, 测量的时间间隔可能影响观测到的脉冲特征值, 因此, 传统的以小时或天尺度进行的类似观测很难捕捉到该特征曲线。此外, 降水强度、土壤的干旱情况、土壤的初始水分状况与呼吸速率的测量时间都可能会对脉冲效应产生影响, 在未来研究中需要加以重视。
4.2 不同草地类型土壤的降水脉冲效应差异显著降水脉冲效应在不同草地类型间存在显著性差异。荒漠草原单位有机质的脉冲效应比草甸草原和典型草原更加显著, 具体表现为荒漠草原单位有机质土壤最大呼吸速率以及48 h内单位有机质累积碳释放量显著高于草甸草原和典型草原, 其中, 土壤有机碳含量的差异可能是上述脉冲效应差异的重要原因。降水后, 微生物可用底物的增加, 加快了其土壤呼吸速率, 但受土壤有机碳含量影响, 不同类型草原的降水脉冲效应强度存在一定差异, 其中有机碳含量最低的荒漠草原的降水脉冲效应强度最小。Jones和Murphy[22]在探讨底物的重要性时发现, 微生物可在60 s内对加入的底物做出反应, 此外, He和Yu[37]也发现在草地和森林土壤中, 微生物呼吸速率与底物呈指数关系, 这也进一步说明不同草地类型土壤有机质含量的差异是影响降水后脉冲效应的重要因素。此外, 底物的化学计量比(碳氮比)也可能会影响微生物对底物分解的有效性[22, 37]。然而, 由于本研究缺乏微生物生物量碳氮比与可溶性有机碳氮比的测试数据, 因此不能从化学计量的角度去准确解释其对底物有效性的影响。未来研究中, 可以通过测量降水脉冲过程中微生物生物量碳氮比与可溶性有机碳氮比, 探究底物有效性对脉冲效应的影响。在降水脉冲事件发生后, 除可用性底物增加外, 微生物生物量的快速增加也能一定程度解释脉冲效应中呼吸速率的增加;Lundquist等人[12]研究发现随着微生物生物量的增加, 呼吸正向增加。此外, 有研究显示微生物的生长与最初的呼吸脉冲并不耦合[38], 本研究也发现有机质含量较低的荒漠草原其在降水事件后, 达到呼吸峰值所用时间也仅为几分钟, 说明微生物呼吸作用的增强可能来源于微生物生物量的增加, 本研究有关ARSOC的结果表明荒漠草原在48 h内微生物呼吸累积量最高, 因此可推测初始阶段微生物生物量与其呼吸是正相关增加的, 但仍需求未来实验大量测定微生物生物量变化数据进行验证。
3种类型草地在土壤复湿后脉冲效应持续时间差异显著, 其中典型草原持续时间显著高于草甸草原和荒漠草原。原因在于脉冲效应的持续时间主要受土壤碳、氮含量以及土壤质地等因素的影响。较多有机质以及砂粒含量更少的土壤, 脉冲效应的持续时间可能更长。Wang等[32]发现有机质含量较高的天然林比次生林的脉冲持续时间更长。Kaiser等[39]的研究表明粘粒含量高的土壤比高沙粒含量的土壤失去水分的速度慢, 微生物有更多的时间适应或响应土壤含水量的下降。本文中荒漠草原土壤具有较高的砂粒含量(表 1), 失去水分的速度更快, 脉冲效应的持续时间较短, 而典型草原相比草甸草原具有更高的土壤有机碳和全氮含量, 脉冲效应的持续时间更长。
4.3 有机质含量、干旱程度共同影响了微生物呼吸对降水脉冲事件的响应土壤有机质含量影响微生物呼吸对降水脉冲事件的响应。降水后单位土壤微生物的最大呼吸速率以及单位土壤累积碳释放量的结果显示, 典型草原土壤对于再湿的响应最为强烈, 草甸草原次之, 而荒漠草原最低。考虑到脉冲效应与土壤中可用基质密切相关, 我们测量了土壤本底有机碳含量, 发现3种草地类型RSoil-max结果与有机碳含量一致, 典型草原有机碳含量高于草甸草原, 荒漠草原有机碳含量最低。现有研究表明微生物的呼吸速率与底物呈正相关关系[23, 31], 部分研究结果显示微生物呼吸速率与底物呈指数关系[37]。本研究发现有机质含量较低的荒漠草原单位有机质土壤呼吸累积释放量ARSOC显著高于有机质含量相对较高的草甸草原与典型草原, 进一步揭示了土壤有机质含量是降水脉冲效应的重要影响因素。
此外, 降水前的土壤干旱状况和降雨频率也会影响土壤CO2释放。干旱一段时间后的土壤会对降水产生快速强烈的响应, 但是其响应强度与干旱时长的关系并未得到验证。我们发现2017—2018年降水次数为典型草原(82)>草甸草原(52)>荒漠草原(34), 降雨次数最少的荒漠草原ARSOC最高, 而降雨频率最高的典型草原ARSOC最低, 据此可推测干旱时间越长、降水频率越低的地区, 土壤对突发降水的反应越强烈。由于不同地区的微生物对资源的获取策略不同, 干旱时期由于缺少食物, 微生物一般选择休眠并积累胞内渗透溶质来应对干旱环境, 而不同气候区的微生物在经历不同的干-湿循环后, 可能对水分形成了不同的机制。本研究结果发现当发生降水事件时, 干旱区微生物相对于湿润区微生物的响应更为强烈, 呼吸速率变化幅度更大, 且在48h内的碳累积释放量也最大, 因此, 干旱区的微生物可能形成特殊的干旱-水分适应机制(“聪明”的微生物), 当经历长时间干旱后, 面对突发降水这一“来之不易”的水资源, 能迅速适应土壤水分的变化, 充分利用短期内快速增加的资源进行生长繁殖;Ellen等的研究也表明微生物能够在短时间尺度上适应土壤水分的变化[25], 这是微生物应对干旱环境所采取的生存策略。有关三类草原在达到峰值后呼吸速率下降的幅度显示, 荒漠草原微生物呼吸速率降低的幅度最大, 且48 h内荒漠草原的微生物对水分的利用最彻底, 这可能是由于荒漠草原在经历长时间干旱后微生物的再湿后生长模式发生改变[25-26], 即随着干旱程度的增加, 微生物对脉冲式降水的响应增强, 碳累积释放量也随之增加。因此, 未来研究应重视土壤的干旱程度对降水脉冲效应的影响[21, 38]。
4.4 降水脉冲效应的研究展望降水脉冲效应作为影响土壤碳氮循环的关键过程, 其内在控制机制以及对生态系统碳循环的贡献等还远未弄清。微生物对降水脉冲事件响应的强烈程度和持续时间与降雨的强度、土壤初始水分状况和降雨前土壤干旱持续时间等密切相关。本文研究了不同草地类型土壤对降水脉冲的响应过程, 并定性分析了土壤有机质含量和干旱程度对降水脉冲事件的影响。相对于其他研究, 我们利用自主研发的微生物呼吸速率自动测定装置, 实现了每个样品快速高频测定(第一次3 min, 随后每隔10 min测量一次)[35], 从而揭示了微生物呼吸速率在降水后的短期动态变化, 既大大提高实验效率, 也为未来探讨类似科学问题提供了坚实的方法学基础。当然, 除了干旱程度外, 土壤其他理化性质、土壤碳氮比(C : N比)、土壤微生物类群及微生物量等也会一定程度影响降水事件的脉冲效应;然而, 如何量化这些因素的影响, 需要进一步的实验探究。自然界中土壤水分处于不断变化的状态, 目前有关不同降水强度对土壤脉冲效应影响的研究相对较少, 未来亟须加强这一方面的研究工作。此外, 土壤干旱和再湿是一个反复的过程, 降水不连续和不同强度导致了土壤干燥和再湿过程的复杂性, 虽然学者们针对关于干燥-再湿循环的过程开展了大量的研究工作, 并在一定程度上揭示了干旱程度以及降水强度对土壤有机质分解的影响, 但这些研究大多采用碱液吸收法在日尺度或间隔几个小时测定土壤呼吸速率, 不能准确地捕捉到短暂的脉冲效应[40]。因此, 未来有关干湿交替的研究应注重实验方法的改进, 同时开展多频度脉冲效应效应的比对实验[8, 24]。在研究内容和研究对象方面, 未来可从区域尺度的探讨多个草地和森林样点的降水脉冲变化规律, 研究经度地带性规律或土壤有机质梯度变化的影响等;同时, 也通过可以增加对脉冲过程中基质有效碳氮的测量与微生物生物量的测定, 进一步解析微生物对脉冲效应的响应机理。
5 结论降水事件对土壤CO2释放的脉冲效应, 对该地区草地生态系统土壤碳循环以及土壤碳平衡具有重要意义。本文利用自主研发的土壤微生物呼吸速率测定装置, 初步探究了内蒙古3种不同类型草地土壤的降水脉冲效应, 发现草甸草原, 典型草原和荒漠草原降水脉冲效应快速而强烈, 其中土壤基质含量和干旱程度(降水频率)等是降水脉冲效应的重要影响因素。其研究思路和研究技术方法可为未来从全国范围开展相关研究提供方法和基础理论的储备。未来应加强相关研究, 以深入揭示降水脉冲效应的时空格局及其控制机制, 并阐明降水脉冲效应对干旱和半干旱地区生态系统碳循环过程的重要作用。
| [1] |
Birch H F. The effect of soil drying on humus decomposition and nitrogen availability. Plant and Soil, 1958, 10(1): 9-31. DOI:10.1007/BF01343734 |
| [2] |
Blazewicz S J, Schwartz E, Firestone M K. Growth and death of bacteria and fungi underlie rainfall-induced carbon dioxide pulses from seasonally dried soil. Ecology, 2014, 95(5): 1162-1172. DOI:10.1890/13-1031.1 |
| [3] |
Collins S L, Sinsabaugh R L, Crenshaw C, Green L, Porras-Alfaro A, Stursova M, Zeglin L H. Pulse dynamics and microbial processes in aridland ecosystems. Journal of Ecology, 2008, 96(3): 413-420. DOI:10.1111/j.1365-2745.2008.01362.x |
| [4] |
Barrow E M, Hulme M. Changing probabilities of daily temperature extremes in the UK related to future global warming and changes in climate variability. Climate Research, 1996, 6: 21-31. DOI:10.3354/cr006021 |
| [5] |
Murray V, Ebi K L. IPCC special report on managing the risks of extreme events and disasters to advance climate change adaptation (SREX). Journal of Epidemiology and Community Health, 2012, 66(9): 759-760. DOI:10.1136/jech-2012-201045 |
| [6] |
Borken W, Matzner E. Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Global Change Biology, 2009, 15(4): 808-824. DOI:10.1111/j.1365-2486.2008.01681.x |
| [7] |
Shi A D, Marschner P. Drying and rewetting frequency influences cumulative respiration and its distribution over time in two soils with contrasting management. Soil Biology and Biochemistry, 2014, 72: 172-179. DOI:10.1016/j.soilbio.2014.02.001 |
| [8] |
Fierer N, Schimel J P. Effects of drying-rewetting frequency on soil carbon and nitrogen transformations. Soil Biology and Biochemistry, 2002, 34(6): 777-787. DOI:10.1016/S0038-0717(02)00007-X |
| [9] |
Lado-Monserrat L, Lull C, Bautista I, Lidón A, Herrera R. Soil moisture increment as a controlling variable of the "Birch effect". Interactions with the pre-wetting soil moisture and litter addition. Plant and Soil, 2014, 379(1/2): 21-34. |
| [10] |
Xiang S R, Doyle A, Holden P A, Schimel J P. Drying and rewetting effects on C and N mineralization and microbial activity in surface and subsurface California grassland soils. Soil Biology and Biochemistry, 2008, 40(9): 2281-2289. DOI:10.1016/j.soilbio.2008.05.004 |
| [11] |
Fischer T. Substantial rewetting phenomena on soil respiration can be observed at low water availability. Soil Biology and Biochemistry, 2009, 41(7): 1577-1579. DOI:10.1016/j.soilbio.2009.04.009 |
| [12] |
Lundquist E J, Jackson L E, Scow K M. Wet-dry cycles affect dissolved organic carbon in two California agricultural soils. Soil Biology and Biochemistry, 1999, 31(7): 1031-1038. DOI:10.1016/S0038-0717(99)00017-6 |
| [13] |
Davidson E A, Verchot L V, Cattânio J H, Ackerman I L, Carvalho J E M. Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia. Biogeochemistry, 2000, 48(1): 53-69. DOI:10.1023/A:1006204113917 |
| [14] |
Huxman T E, Snyder K A, Tissue D, Leffler A J, Ogle K, Pockman W T, Sandquist D R, Potts D L, Schwinning S. Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia, 2004, 141(2): 254-268. DOI:10.1007/s00442-004-1682-4 |
| [15] |
Schwinning S, Sala O E. Hierarchy of responses to resource pulses in arid and semi-arid ecosystems. Oecologia, 2004, 141(2): 211-220. DOI:10.1007/s00442-004-1520-8 |
| [16] |
Sala O E, Lauenroth W K. Small rainfall events:an ecological role in semiarid regions. Oecologia, 1982, 53(3): 301-304. DOI:10.1007/BF00389004 |
| [17] |
Cook F J, Orchard V A. Relationships between soil respiration and soil moisture. Soil Biology and Biochemistry, 2008, 40(5): 1013-1018. DOI:10.1016/j.soilbio.2007.12.012 |
| [18] |
Schimel J, Balser T C, Wallenstein M. Microbial stress-response physiology and its implications for ecosystem function. Ecology, 2007, 88(6): 1386-1394. DOI:10.1890/06-0219 |
| [19] |
Wang L X, Manzoni S, Ravi S, Riveros-Iregui D, Caylor K. Dynamic interactions of ecohydrological and biogeochemical processes in water-limited systems. Ecosphere, 2015, 6(8): 133. DOI:10.1890/ES15-00122.1 |
| [20] |
Fierer N, Schimel J P. A proposed mechanism for the pulse in carbon dioxide production commonly observed following the rapid rewetting of a dry soil. Soil Science Society of America Journal, 2003, 67(3): 798-805. DOI:10.2136/sssaj2003.0798 |
| [21] |
Sponseller R A. Precipitation pulses and soil CO2 flux in a Sonoran Desert ecosystem. Global Change Biology, 2007, 13(2): 426-436. DOI:10.1111/j.1365-2486.2006.01307.x |
| [22] |
Jones D L, Murphy D V. Microbial response time to sugar and amino acid additions to soil. Soil Biology and Biochemistry, 2007, 39(8): 2178-2182. DOI:10.1016/j.soilbio.2007.03.017 |
| [23] |
Esch E H, Lipson D, Cleland E E. Direct and indirect effects of shifting rainfall on soil microbial respiration and enzyme activity in a semi-arid system. Plant and Soil, 2017, 411(1/2): 333-346. |
| [24] |
Sun Q Q, Meyer W S, Marschner P. Repeated rainfall in summer induces prolonged high soil respiration in a semi-arid floodplain woodland. Ecohydrology, 2018, 11(7): e1984. DOI:10.1002/eco.1984 |
| [25] |
Wu J, Brookes P C. The proportional mineralisation of microbial biomass and organic matter caused by air-drying and rewetting of a grassland soil. Soil Biology and Biochemistry, 2005, 37(3): 507-515. DOI:10.1016/j.soilbio.2004.07.043 |
| [26] |
Morillas L, Roales J, Portillo-Estrada M, Gallardo A. Wetting-drying cycles influence on soil respiration in two Mediterranean ecosystems. European Journal of Soil Biology, 2017, 82: 10-16. DOI:10.1016/j.ejsobi.2017.07.002 |
| [27] |
Waring B G, Powers J S. Unraveling the mechanisms underlying pulse dynamics of soil respiration in tropical dry forests. Environmental Research Letters, 2016, 11(10): 105005. DOI:10.1088/1748-9326/11/10/105005 |
| [28] |
Bottner P. Response of microbial biomass to alternate moist and dry conditions in a soil incubated with 14C- and 15N-labelled plant material. Soil Biology and Biochemistry, 1985, 17(3): 329-337. DOI:10.1016/0038-0717(85)90070-7 |
| [29] |
Kieft T L, Soroker E, Firestone M K. Microbial biomass response to a rapid increase in water potential when dry soil is wetted. Soil Biology and Biochemistry, 1987, 19(2): 119-126. DOI:10.1016/0038-0717(87)90070-8 |
| [30] |
Appel T. Non-biomass soil organic N- the substrate for N mineralization flushes following soil drying-rewetting and for organic N rendered CaCl2-extractable upon soil drying. Soil Biology and Biochemistry, 1998, 30(10/11): 1445-1456. |
| [31] |
Denef K, Six J, Bossuyt H, Frey S D, Elliott E T, Merckx R, Paustian K. Influence of dry-wet cycles on the interrelationship between aggregate, particulate organic matter, and microbial community dynamics. Soil Biology and Biochemistry, 2001, 33(12/13): 1599-1611. |
| [32] |
Wang Q, He N P, Liu Y, Li M L, Xu L. Strong pulse effects of precipitation events on soil microbial respiration in temperate forests. Geoderma, 2016, 275: 67-73. DOI:10.1016/j.geoderma.2016.04.016 |
| [33] |
Scheu S, Parkinson D. Changes in bacterial and fungal biomass C, bacterial and fungal biovolume and ergosterol content after drying, remoistening and incubation of different layers of cool temperate forest soils. Soil Biology and Biochemistry, 1994, 26(11): 1515-1525. DOI:10.1016/0038-0717(94)90093-0 |
| [34] |
Liu Y, He N P, Zhu J X, Xu L, Yu G R, Niu S L, Sun X M, Wen X F. Regional variation in the temperature sensitivity of soil organic matter decomposition in China's forests and grasslands. Global Change Biology, 2017, 23(8): 3393-3402. DOI:10.1111/gcb.13613 |
| [35] |
He N P, Wang R M, Gao Y, Dai J Z, Wen X F, Yu G R. Changes in the temperature sensitivity of SOM decomposition with grassland succession:implications for soil C sequestration. Ecology and Evolution, 2013, 3(15): 5045-5054. DOI:10.1002/ece3.881 |
| [36] |
Miller A E, Schimel J P, Meixner T, Sickman J O, Melack J M. Episodic rewetting enhances carbon and nitrogen release from chaparral soils. Soil Biology and Biochemistry, 2005, 37(12): 2195-2204. DOI:10.1016/j.soilbio.2005.03.021 |
| [37] |
He N P, Yu G R. Stoichiometrical regulation of soil organic matter decomposition and its temperature sensitivity. Ecology and Evolution, 2016, 6(2): 620-627. DOI:10.1002/ece3.1927 |
| [38] |
Meisner A, Bååth E, Rousk J. Microbial growth responses upon rewetting soil dried for four days or one year. Soil Biology and Biochemistry, 2013, 66: 188-192. DOI:10.1016/j.soilbio.2013.07.014 |
| [39] |
Kaiser M, Kleber M, Berhe A A. How air-drying and rewetting modify soil organic matter characteristics:an assessment to improve data interpretation and inference. Soil Biology and Biochemistry, 2015, 80: 324-340. DOI:10.1016/j.soilbio.2014.10.018 |
| [40] |
李凌浩, 韩兴国, 王其兵, 陈全胜, 张焱, 杨晶, 闫志丹, 李鑫, 白文明, 宋世环. 锡林河流域一个羊草群落中土壤呼吸与生物量之间的相关性分析(英文). 植物学报, 2002, 44(5): 593-597. DOI:10.3321/j.issn:1672-9072.2002.05.016 |