 2020, Vol. 40
2020, Vol. 40文章信息
- 李建, 李晓宇, 曹静, 谭凌照, 赵秀海
- LI Jian, LI Xiaoyu, CAO Jing, TAN Lingzhao, ZHAO Xiuhai
- 长白山次生针阔混交林群落结构特征及群落动态
- Community structure and dynamics of secondary coniferous and broad-leaved mixed forest in Changbai Mountains
- 生态学报. 2020, 40(4): 1195-1206
- Acta Ecologica Sinica. 2020, 40(4): 1195-1206
- http://dx.doi.org/10.5846/stxb201812232788
-
文章历史
- 收稿日期: 2018-12-23
- 网络出版日期: 2019-11-20






东北林区是我国主要的森林分布区之一, 其在保持水土、生产木材和为动物提供栖息地等方面发挥重要作用[1]。然而在20世纪, 由于过度采伐和保育不当等人为因素导致了天然林面积大量减少, 造成林分结构被严重破坏, 森林质量下降和生态系统功能严重退化等后果, 由此产生大面积次生林[2-3]。据第八次全国森林资源清查结果显示, 东北地区的次生林面积超过60%, 存在生长缓慢, 结构不合理, 更新能力弱和林分质量低下等问题亟待改善[4-5]。21世纪以来, 随着生态环境日益严峻和人们生态保护意识逐渐提高, 次生林的可持续经营和恢复逐渐成为研究重点[6-7]。
森林结构是实现森林经营目标必须测定和调控的属性, 其代表着一系列定量描述森林结构特性的指标[8-9]。传统的森林结构指标包括物种组成、物种多样性、胸径结构和密度结构等, 通常被用来描述森林特征的平均状态[10]。21世纪以来, 随着森林多目标经营理念的提出, 国内外学者相继提出若干基于林木个体空间坐标的林分空间结构指标, 从林木空间分布格局、树种混交和树木竞争等方面反映林分内部的结构变异, 助力森林质量精准提升[11-12]。结合森林非空间结构和空间结构, 将为探究群落动态、提高生态系统服务和功能、增强森林稳定性和可持续性提供全面系统的数据支撑[13-15]。
尽管国内学者已经开展了大量关于东北地区次生林群落结构的研究, 但是往往受限于调查面积过小或研究周期较短等原因而难以揭示一般性规律[16]。在分析群落动态时, 部分学者采用空间代替时间的办法, 通过构建演替序列描述群落发展态势[17]。这种方法虽然可在短期内获取群落演替的宏观动态, 但却不易消除空间异质性且不能反映干扰在群落动态中的作用, 导致研究结果存在局限性[18]。因此, 对森林群落动态进行长期监测成为科学研究的迫切需求。继巴拿马Barro Colorado岛内建立的首个50hm2森林动态监测样地之后[19], 我国学者于本世纪初期开始在全国范围内建立起一系列永久监测样地, 并构建了中国森林生物多样性监测网络(CForBio)[20]。十余年来, 国内学者依托于森林动态监测样地对多种群落类型的演替动态展开研究, 极大促进了物种共存和群落构建等生态学机理研究的蓬勃发展[21]。
温带次生针阔混交林是长白山地区重要的森林类型之一, 是对地带性顶极植被原始阔叶红松林进行采伐或火烧干扰后自然恢复形成的天然林群落。目前, 已有学者描述了温带次生针阔混交林群落的结构特征[22-24], 但对该群落在10年以上的长期结构动态鲜有报道。本文以一块面积为5.2hm2的长白山次生针阔混交林动态监测样地为基础, 利用2005年和2017年的调查数据, 研究次生针阔混交林的群落结构在较长时间内是否发生改变及变化趋势, 旨在揭示该地区的次生林演替规律, 为结构化森林经营提供理论依据。
1 研究区概况与研究方法 1.1 研究地概况研究地点位于吉林省白河林业局光明林场, 海拔899m, 该区域受温带大陆性季风气候影响, 年均气温3.6℃, 最冷月1月份的平均气温-15.4℃;夏季短暂且温暖多雨, 最热月7月份的平均气温19.6℃, 年均降水量707mm[25];土壤类型为富含有机质的山地暗棕色森林土, 土层厚度20—100cm。该地区物种属于长白山植物区系, 乔木树种主要包括臭松(Abies nephrolepis)、紫椴(Tilia amurensis)、白桦(Betula platyphylla)、假色槭(Acer pseudosieboldianum)、色木槭(Acer mono)、红松(Pinus koraiensis)、蒙古栎(Quercus mongolica)和水曲柳(Fraxinus mandschurica)等[26]。
1.2 样地设置及植被调查方法2005年, 在吉林省白河林业局光明林场内, 选取处于演替中期的次生针阔混交林, 参照美国史密森热带研究所热带森林研究中心(Center for Tropical Forest Science, CTFS)建设标准和操作规范, 利用全站仪建立了投影面积为5.2 hm2(长260m, 宽200m)的森林动态监测样地, 样地坐标为东经128°0.7988′、北纬42°20.907′, 平均海拔为805m[27]。将样地进一步划分为20m×20m样方后, 于同年对样地内所有胸径≥1cm的木本植物开展调查, 记录物种名、胸径、树高、冠幅、枝下高和相对位置等信息并挂牌标记。此后, 分别于2010和2017年对样地复查。本研究以2005年和2017年的调查数据中所有胸径≥5cm的木本植物为研究对象。
1.3 非空间结构特征及动态计算方法 1.3.1 物种组成特征和动态重要值(IV)是衡量物种在群落中相对重要性的指标[28], 计算公式为:

|
式中:相对多度(%) =100×(某个物种的株数/所有物种的总株数);相对频度(%)=100×(某个物种在统计样方中出现的次数/所有物种出现的总次数);相对优势度(%)=100×(某个物种的胸高断面积/所有物种的胸高断面积和)。
计算各物种在2005—2017年间的多度、频度、胸高断面积和重要值的变化率, 并采用Wilcoxon符号秩检验分析变化趋势[9], 变化率公式如下:
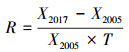
|
式中:R表示变化率, X2017和X2005分别表示物种在两次调查中的多度、频率、胸高断面积或重要值, T表示两次调查的时间间隔。
对物种的多度、频度、胸高断面积和重要值4个特征值进行对数转化后, 采用Speraman相关性检验分析在2005—2017年间的物种次序动态。假如物种的某个特征次序在某时段前后没有变化, 则Spearman相关系数为1;假如物种某个特征次序在某一时段前后差异极大, 则该相关系数为-1;总之, Spearman相关系数越大, 则物种特征变化越小;反之, 则该特征变化越大[12]。文中所有统计学显著性水平设为P<0.05。
1.3.2 物种多样性特征和动态采用物种丰富度(S)、Shannon-Weiner多样性指数(H)、Simpson优势度指数(D)和Pielou均匀度指数(J)4个指标分别计算样地水平和样方水平的物种多样性。物种多样性的计算公式如下[29]:
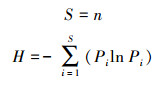
|
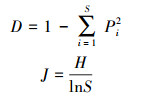
|
式中:n表示某物种的数量;Pi表示样方内第i个种的数量占总体数量的比例。采用Wilcoxon符号秩检验分析样方水平上物种多样性的变化趋势, 对于其他指标也采用相同方法分析动态变化[27]。
1.3.3 胸径结构特征和动态胸径结构是一个容易获取且精确度较高的指标, 实际研究中常用胸径结构代替年龄结构。径级结构描述了林木直径大小分布特征, 可在一定程度上反映群落的可持续性。本文以2cm为径级分析群落胸径结构特征, 再对胸径进行对数转换并采用Pearson和Spearman相关性检验分析群落径级结构动态[28]。
采用胸径变异系数(VarD)和胸径基尼系数(GiniD)描述样地整体和样方水平的胸径结构离散程度。其中, 胸径变异系数是描述林木分布频率的常用指标, 系数值越大则林木大小分化程度越高;基尼系数是描述个体胸径差异偏离绝对均匀程度的指标, 当个体间没有差异时取值为0, 当差异最大化时取值无限接近1[30]。两个指标的计算公式为:
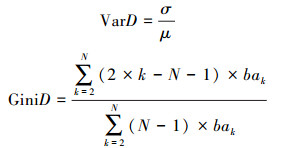
|
式中:σ表示胸径标准差, μ表示胸径平均值, bak表示按升序第k个个体的胸高断面积。
1.3.4 密度结构样地或样方水平的林分密度(stand density, StaD, 单位:m2/hm2)和株数密度(strains density, StrD, 单位:株/m2)分别用个体的胸高断面积和与数量表示。两个指标的计算公式为:
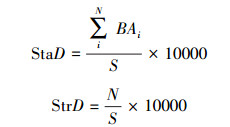
|
式中:BAi表示某植株的胸高断面积, N表示某一尺度下所有个体数量;S表示样地或样方面积(m2)。
1.4 空间结构特征及动态计算方法 1.4.1 空间分布格局采用角尺度(Wi)描述林分空间分布格局[31], 计算公式为:
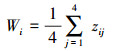
|
式中:zij表示参照木i与相邻木j所构成的夹角α与标准角α0(α0=72°)的关系, 当α>α0时, zij=0;否则zij=1。用平均角尺度(W)表征群落水平分布格局, W < 0.475表示均匀分布, W∈0.475, 0.517表示随机分布, W>0.517表示聚集分布[32]。
1.4.2 树种混交通过计算混交度(Mi)表征树种隔离程度[33], 计算公式如下:
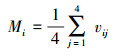
|
式中:vij表示参照木i与相邻木j的身份关系, 当i与j为同一物种时, vij=0, 否则vij=1。平均混交度(M)可被用来判断群落水平的混交程度, 混交度越高则群落越稳定。
1.4.3 个体竞争相邻林木之间的大小分化差异和所处地位优劣可通过计算胸径大小比数得知[34], 计算公式如下:
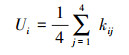
|
式中:kij表示参照木i与相邻木j的大小关系, 当i>j时, kij=0;否则kij=1。平均大小比数(U)能反映群落水平的优势木比例。
1.4.4 林分空间结构状态根据上述3个空间结构指标的定义, 理想的林分空间结构为林分平均混交度是100, 林分平均大小比数是0, 林分平均角尺度是50。林分空间结构距离指数(FSSD)用于描述林分实际空间结构向理想空间结构逼近或远离的趋势, FSSD值越小则林分越接近理想状态[35], 其计算公式为:
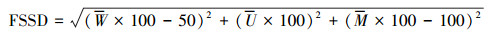
|
式中, 0≤FSSD≤150。
1.5 数据处理本文应用R3.4.3、Winkelmass和Excel软件进行数据的统计与分析。
2 结果与分析 2.1 非空间结构特征和动态 2.1.1 群落物种组成2005和2017年的群落调查分别发现13科23属40种和13科25属42种木本植物, 共有植物13科23属39种。两次调查期间, 物种组成几乎没有变化, 仅4个偶见种(总株数≤3)出现变动[36], 其中毛山楂(Crataegus maximowiczii), 拧筋槭(Acer triflorum)和山丁子(Malus baccata)进入群落, 沙松(Abies holophylla)退出群落。
温带群落的物种数量相对有限, 处于不同演替阶段的优势种群差异明显。2005—2017年, 重要值排在前10位的优势物种组成基本保持不变, 仅香杨替代山杨排在末位。优势物种在群落内的优势度略微扩大, 重要值累计由77.79提高至78.78。大部分优势物种的胸高断面积均提高, 其中增长量排在前5位的树种分别是臭松(0.45 m2/hm2)、紫椴(0.41 m2/hm2)、水曲柳(0.29m2/hm2)、红松(0.24 m2/hm2)和白桦(0.18 m2/hm2)。仅怀槐和山杨的胸高断面积缩减, 分别减少0.09 m2/hm2和0.53 m2/hm2。
2005—2017年, 群落全部个体数量由6614株降至5884株, 减少730株;全部优势种群数量由5630株降至5042株, 减少588株。大部分优势物种数量减少, 其中白桦、紫椴和怀槐的种群数量损失较多, 分别减少180株、142株和106株(表 1)。12年间, 大部分优势种群动态表现为多度增加但胸高断面积减少, 说明种群内小径级个体损失较多;怀槐和山杨的多度和胸高断面积均减少, 说明该种群内的大径级个体也出现大量死亡现象。
| 树种Species | 重要值Important value | 胸高断面积Basal area/(m2/hm2) | 多度Abundance | ||||||||
| 2005 | 2017 | 变化率/% | 2005 | 2017 | 变化率/% | 2005 | 2017 | 变化率/% | |||
| 紫椴Tilia amurensis | 13.32 | 13.26 | -0.04 | 6.35 | 6.76 | 0.54 | 806 | 664 | -1.62 | ||
| 臭松Abies nephrolepis | 12.36 | 13.50 | 0.77 | 2.58 | 3.03 | 1.45 | 1370 | 1338 | -0.20 | ||
| 白桦Betula platyphylla | 11.19 | 10.58 | -0.45 | 5.53 | 5.70 | 0.26 | 683 | 503 | -2.55 | ||
| 假色槭Acer pseudosieboldianum | 8.88 | 10.22 | 1.26 | 1.14 | 1.29 | 1.10 | 1004 | 1075 | 0.57 | ||
| 色木槭Acer mono | 7.54 | 8.10 | 0.62 | 1.95 | 2.06 | 0.47 | 563 | 577 | 0.20 | ||
| 红松Pinus koraiensis | 6.68 | 6.92 | 0.30 | 2.19 | 2.43 | 0.91 | 394 | 353 | -0.92 | ||
| 蒙古栎Quercus mongolica | 5.61 | 5.59 | -0.03 | 2.51 | 2.60 | 0.30 | 201 | 168 | -1.49 | ||
| 水曲柳Fraxinus mandschurica | 4.38 | 4.44 | 0.11 | 2.00 | 2.28 | 1.17 | 205 | 168 | -1.66 | ||
| 怀槐Maackia amurensis | 4.05 | 3.08 | -2.00 | 0.82 | 0.73 | -0.91 | 247 | 141 | -4.67 | ||
| 山杨Populus davidiana | 3.78 | 2.50 | -2.82 | 1.80 | 1.27 | -2.45 | 157 | 77 | -5.94 | ||
| 香杨Populus koreana | 3.24 | 3.09 | -0.39 | 1.97 | 2.09 | 0.51 | 72 | 55 | -2.24 | ||
总体上, 所有物种的多度显著减小(P<0.01), 频度显著减小(P<0.05), 胸高断面积显著增大(P<0.05), 重要值无明显变化(P>0.05)(图 1)。物种多度、频度、胸高断面积和重要值的排序无显著变化(P<0.001), 种间差异保持相对稳定。

|
| 图 1 2005—2017物种多度、频度、胸高断面积和重要值变化率 Fig. 1 Changes in species abundance, frequency, area of basal area and the important value between 2005 and 2017 a—d分别代表物种多度、频度、胸高断面积和重要值的变化率, 虚线左侧的变化率为负值 |
两次调查中, 群落的物种丰富度分别为40和42, Shannon-Wiener指数分别为2.59和2.50, Simpson指数分别为0.89和0.88, Pielou均匀度指数分别为0.70和0.69(表 2)。在20m×20m的样方的尺度上, 物种丰富度、Shannon-Wiener多样性和Simpson优势度均显著减小(P<0.001), 而Pielou均匀度没有显著变化(P>0.05, 图 2)。
| 非空间结构 Non-spatial structure |
非空间结构指数 Non-spatial structure index |
2005年 | 2017年 |
| 物种多样性Species diversity | 物种丰富度 | 40 | 42 |
| Shannon-Wiener多样性 | 2.59 | 2.50 | |
| Simpson优势度 | 0.89 | 0.88 | |
| Pielou均匀度 | 0.70 | 0.69 | |
| 胸径结构DBH structure | 胸径变异系数 | 0.77 | 0.79 |
| 胸径基尼系数 | 0.37 | 0.38 | |
| 密度结构Density structure | 林分密度(m2/hm2) | 33.12 | 34.74 |
| 株数密度(株/hm2) | 1272 | 1132 |

|
| 图 2 2005—2017年次生针阔混交林群落空间结构动态变化 Fig. 2 Dynamics of spatial structure of secondary coniferous and broad-leaved mixed forest community from 2005 to 2017 箱形图中加粗实线表示样本的中位数, 箱形的上下边分别表示四分之一位数和四分之三位数;每个对比图上方字母代表组间差异是否显著, 如果差异显著则字母不同(P<0.05), 如果差异不显著则字母相同(P>0.05) |
由图 3可知, 群落径级结构均呈明显的倒“J”型分布。2005—2017年, 整体上群落各径级的个体数量显著减少(P<0.001), 但径级间的相对个体数差异没有显著变化(ρ=0.98, P<0.001), 群落径级结构保持稳定。

|
| 图 3 2005年和2017年群落径级结构 Fig. 3 DBH size class community structure in 2005 and 2017 |
2005年样地内木本植物的胸径平均值为14.4cm, 群落胸径变异系数是0.77, 胸径基尼系数是0.37。2017年全部个体的平均胸径为15.5cm, 群落胸径的变异系数是0.79, 胸径基尼系数是0.38(表 2)。12年间, 样方尺度上的胸径变异系数和基尼系数均显著增加(P<0.001)(图 2), 与样地整体的变化趋势一致。
2.1.4 密度结构次生针阔混交林群落样地在2005年的林分密度为33.12m2/hm2, 株数密度为1272株/ hm2;2017年的林分密度为34.74m2/hm2, 株数密度为1132株/hm2。12年间, 林分密度提升1.62m2/hm2, 株数密度降低140株/ hm2(表 2)。在样方尺度和样地尺度的密度结构变化趋势一致, 林分密度显著增加(P<0.001), 株数密度显著减小(P<0.001)(图 2)。
2.2 空间结构特征和动态2005年和2017年的林分空间结构距离保持在55.47不变(表 3), 群落空间结构的综合情况较为理想, 群落空间结构基本保持稳定(图 2)。
| 空间结构 Spatial structure |
空间结构指数 Spatial structure index |
2005年 | 2017年 |
| 空间分布格局Spatial distribution pattern | 角尺度 | 0.51 | 0.51 |
| 树种混交Species spatial heterogeneity | 混交度 | 0.76 | 0.76 |
| 个体竞争Individual competition | 大小比数 | 0.50 | 0.50 |
| 空间结构理想态Ideal spatial structure | 空间结构距离 | 55.47 | 55.47 |
采用角尺度描述群落的水平分布格局, 2005和2017年群落的平均角尺度均为0.50, 群落整体处于随机分布(表 3)。从分布频率上看, 群落内大部分个体处于随机分布(W=0.5), 处于聚集分布(W=0或0.25)和均匀分布(W=0.75或1)的个体比例比较接近(图 4)。样方尺度上, 群落角尺度在12年间无显著差异(P>0.05)(图 2)。

|
| 图 4 2005和2017年林木空间结构频率分布 Fig. 4 Frequency distribution of individuals spatial structure in 2005 and 2017 |
采用混交度指标描述种间隔离度, 2005年和2017年的群落平均混交度均为0.76, 林分处于强度混交(表 3)。各取值的分布频率随取值增大呈单调递增趋势, 处于完全混交的个体比例最高, 处于完全不混交的个体比例最低(图 4)。12年间, 样方水平混交度无显著差异(P>0.05)(图 2)。
2.2.3 林木个体竞争采用大小比数描述群落内个体间的大小分化程度, 2005和2017年的群落平均大小比数均为0.50(表 3), 说明林分内个体分布比较均匀, 处于不同竞争强度的个体比例较为接近, 林分整体呈中庸分布(图 4)。12年间, 样方水平大小比数无显著差异(P>0.05)(图 2)。
3 讨论和结论 3.1 群落非空间结构动态群落次生演替过程一直是生态学理论的研究热点, 演替过程中群落的结构动态和稳定性仍然存在争论[37]。2005—2017年, 次生针阔混交林群落的物种组成呈增加趋势, 物种丰富度的变化主要由偶见种引起, 体现了偶见种在维持群落生物多样性的重要作用[38]。12年间, 优势物种在群落内的相对重要性发生改变, 白桦、山杨和香杨等先锋树种的重要值降低, 臭松、红松、水曲柳、假色槭和色木槭等顶级树种和林下层树种的重要值提高。郭利平等研究表明, 长白山阔叶红松林群落演替的实质是白桦、山杨、黄檗和春榆等阳性或半阴性树种随演替进程逐渐被紫椴、水曲柳、红松和色木槭等阴性树种取代的过程[39], 本研究结果基本符合这一变化趋势, 表明群落处于正向演替过程。此外, 怀槐和山杨的种群数量和胸高断面积均明显降低, 可预见该种群将由优势转为劣势;假色槭和色木槭的种群数量和胸高断面积均增加, 反映了林下层树种的竞争优势逐渐增强, 将可能对林冠层树种的更新造成阻碍。
群落径级结构在两次调查中均呈倒“J”型, 群落保持较为稳定的更新态势, 侧面反映了该群落在12年内未曾受到严重的人为干扰, 大部分种群能完成自我更新。群落个体之间的大小差异还能反映群落的复杂度和稳定性[40], 已有研究结果表明, 林木大小分化程度随演替进程逐渐提高, 如Lei等对不同林龄的云杉混交林的结构多样性研究表明, 林木胸径基尼系数随着林分生长发育呈增大趋势但差异不显著[41];白超等结合6个指标比较多种混交林类型发现:天然林和老龄林比人工林和中龄林具有更高的林木大小多样性[40]。与上述观点一致, 本文研究群落的胸径变异系数和基尼系数均提高, 反映了群落的正向演替进程。
密度结构与种群周转速率密切相关, 是传统森林经营评价和调控的必要指标[42]。株数密度和林分密度均会随着群落发展而改变, 张春雨等对长白山处于不同演替阶段群落特征的研究表明, 株数密度随演替进程逐渐减少, 林分密度逐渐增加[43], 本文研究结果也体现了这一变化趋势。然而一项对处于不同演替阶段的16个热带森林群落的5年监测研究表明, 林分密度随演替进程逐渐升高, 株数密度仅在演替初期呈快速上升态势并在演替中后期保持平稳[44]。造成上述差异的原因可能是热带群落具有更高的物种多样性、更长的生长季和更快的更新速率, 温带群落在温湿度、降水量和光周期等环境条件上受到更多局限[17]。
3.2 群落空间结构动态植物个体的生长、死亡和更新动态将直接影响群落空间结构, 良好的林分空间结构是判断群落能否维持稳定发展的重要前提[8]。通常认为, 当水平分布格局为随机分布、大小分化程度为中庸分布并且树种混交度较强时, 群落具有比较稳定的群落结构[9]。两次调查中, 本文研究的次生针阔混交林群落的水平分布格局为随机分布(W=0.51), 大小分化程度为中庸分布(U=0.50), 混交度为强度混交(M=0.76), 体现了群落具有良好且稳定的空间结构。与夏富才等对长白山北坡阔叶红松林(W=0.58, M=0.53)[45]和次生杨桦林(W=0.58, M=0.54)[46]的空间结构的研究相比, 本文研究的处于演替中期的群落空间结构更加理想和稳定。关于群落演替过程中群落空间结构是否会发生变化还需持续监测研究。
彭辉等对热带BCI50hm2样地活立木空间结构的研究表明, 群落的平均角尺度、平均大小比数和平均混交度分别为0.504、0.504和0.911, 其中群落平均混交度明显高于本文研究的温带森林群落, 其原因可能是热带森林具有更高的物种多样性和株数密度(第7次调查发现297个物种, 约4028株/hm2), 提高了处于强度混交(M=0.75或1)的个体比例[47]。未来探讨群落混交度时应考虑加入物种多样性和株数密度等非空间结构指标, 增强对不同群落类型空间结构差异的分辨力。
3.3 群落动态的尺度效应群落结构受生境异质性、植物繁殖体传播范围和种间相互作用等因素影响具有尺度依赖性, 通过不同尺度的群落结构动态反映群落稳定性和演替规律更具说服力[48-49]。因此, 本文不仅探讨了样地尺度下的群落结构整体水平, 还在20m×20m样方尺度下对比群落12年间动态, 使群落动态趋势更加明确且更具说服力。研究发现, 大部分指标在上述两个尺度上呈现一致的变化趋势, 表明不同尺度上的群落结构动态具有一致性。然而也发现个别指标出现不一致的变化趋势, 其中物种丰富度在样地和样方尺度上具有相反的变化趋势, 样地尺度上调查到偶见种的增减可能会影响对群落演替态势的判断, 小尺度上物种丰富度显著降低更能反映群落整体演替态势, 群落物种丰富度在小尺度上的减少趋势也与前人研究观点一致[43];Pielou均匀度指数虽然在样地尺度上呈现降低态势, 但是在样方尺度上并无显著变化, 体现出物种分布格局在12年间无明显改变。
3.4 结论综上, 本文结合群落非空间结构和空间结构指标, 综合分析长白山次生针阔混交林群落结构动态。12年间, 群落非空间结构指标具有明显变化而空间结构并无明显变化, 且大部分指标在样地和样方两个尺度上变化趋势一致, 体现群落演替过程中“稳中有变”的特点。对该群落经营时可考虑适当伐除色木槭和假色槭等林下层树种, 为红松、蒙古栎和紫椴等顶级树种的更新提供空间, 在保证群落稳定性的前提下促进群落正向演替。本文研究结果虽然体现出群落具有正向演替态势, 但并未发现群落演替中“质”的变化, 未来还需对该次生针阔混交林群落持续开展动态监测, 并结合环境因素从不同角度进一步探究群落演替规律。
| [1] |
李明, 柴旭荣, 张连芝. 中国东北三省森林生物量及其空间格局. 东北师大学报:自然科学版, 2017, 49(3): 131-135. |
| [2] |
刘国华, 傅伯杰, 陈利顶, 郭旭东. 中国生态退化的主要类型、特征及分布. 生态学报, 2000, 20(1): 13-19. |
| [3] |
Deng X Z, Jiang Q O, Zhan J Y, He S J, Lin Y Z. Simulation on the dynamics of forest area changes in Northeast China. Journal of Geographical Sciences, 2010, 20(4): 495-509. DOI:10.1007/s11442-010-0495-0 |
| [4] |
国家林业局. 第八次全国森林资源清查结果. 林业资源管理, 2014(1): 1-2. |
| [5] |
宋庆丰.中国近40年森林资源变迁动态对生态功能的影响研究[D].北京: 中国林业科学研究院, 2015. http://cdmd.cnki.com.cn/Article/CDMD-82201-1015622494.htm
|
| [6] |
Yu D P, Zhou L, Zhou W M, Ding H, Wang Q W, Yue W, Wu X Q, Dai L M. Forest management in Northeast China:history, problems, and challenges. Environmental Management, 2011, 48(6): 1122-1135. DOI:10.1007/s00267-011-9633-4 |
| [7] |
刘世荣, 代力民, 温远光, 王晖. 面向生态系统服务的森林生态系统经营:现状、挑战与展望. 生态学报, 2015, 35(1): 1-9. DOI:10.3969/j.issn.1673-1182.2015.01.001 |
| [8] |
汤孟平. 森林空间结构研究现状与发展趋势. 林业科学, 2010, 46(1): 117-122. |
| [9] |
惠刚盈. 基于相邻木关系的林分空间结构参数应用研究. 北京林业大学学报, 2013, 35(4): 1-8. |
| [10] |
王均伟.长白山阔叶红松林主要树种空间格局及环境解释[D].北京: 北京林业大学, 2016. http://cdmd.cnki.com.cn/Article/CDMD-10022-1016140196.htm
|
| [11] |
邓红兵, 陈喆菲, 许东, 赵小汎. 森林多目标经营单元的时空尺度扩展. 生态学报, 2015, 35(1): 65-70. |
| [12] |
曹小玉, 李际平, 封尧, 胡园杰, 张彩彩, 房晓娜, 邓超. 杉木生态公益林林分空间结构分析及评价. 林业科学, 2015, 51(7): 37-48. |
| [13] |
Shugart H H, Saatchi S, Hall F G. Importance of structure and its measurement in quantifying function of forest ecosystems. Journal of Geophysical Research:Biogeosciences, 2010, 115(G2): G00E13. |
| [14] |
Forrester D I. The spatial and temporal dynamics of species interactions in mixed-species forests:from pattern to process. Forest Ecology and Management, 2014, 312: 282-292. DOI:10.1016/j.foreco.2013.10.003 |
| [15] |
Seidl R, Schelhaas M J, Rammer W, Verkerk P J. Increasing forest disturbances in Europe and their impact on carbon storage. Nature Climate Change, 2014, 4(9): 806-810. DOI:10.1038/nclimate2318 |
| [16] |
宋厚娟, 叶吉, 蔺菲, 原作强, 郝占庆, 王绪高. 取样面积对森林木本植物空间分布格局分析的影响. 科学通报, 2014, 59(24): 2388-2395. |
| [17] |
Lohbeck M, Poorter L, Martínez-Ramos M, Rodriguez-Velázquez J, Van Breugel M, Bongers F. Changing drivers of species dominance during tropical forest succession. Functional Ecology, 2014, 28(4): 1052-1058. DOI:10.1111/1365-2435.12240 |
| [18] |
徐玮泽, 刘琪璟, 孟盛旺, 周光. 长白山阔叶红松林树木种群动态的长期监测. 应用生态学报, 2018, 29(10): 3159-3166. |
| [19] |
Condit R. Research in large, long-term tropical forest plots. Trends in Ecology & Evolution, 1995, 10(1): 18-22. |
| [20] |
米湘成, 郭静, 郝占庆, 谢宗强, 郭柯, 马克平. 中国森林生物多样性监测:科学基础与执行计划. 生物多样性, 2016, 24(11): 1203-1219. DOI:10.17520/biods.2015313 |
| [21] |
马克平. 森林动态大样地是生物多样性科学综合研究平台. 生物多样性, 2017, 25(3): 227-228. |
| [22] |
闫琰, 张春雨, 赵秀海. 长白山不同演替阶段针阔混交林群落物种多度分布格局. 植物生态学报, 2012, 36(9): 923-934. |
| [23] |
陈亚南, 杨华, 马士友, 任玫玫. 长白山2种针阔混交林空间结构多样性研究. 北京林业大学学报, 2015, 37(12): 48-58. |
| [24] |
孙越, 夏富才, 赵秀海, 轩志龙, 徐敏敏, 李良. 张广才岭温带次生针阔混交林物种组成和群落结构特征. 生态学报, 2017, 37(10): 3425-3436. |
| [25] |
张春雨.长白山针阔混交林种群结构及环境解释[D].北京: 北京林业大学, 2009.
|
| [26] |
赵云萍, 刘玉安, 李广祥, 孙淑艳. 长白山林区复层异龄林垂直结构的研究. 吉林林学院学报, 1995, 11(2): 105-108. |
| [27] |
倪瑞强.长白山典型针阔混交林群落结构与动态研究[D].北京: 北京林业大学, 2014. http://cdmd.cnki.com.cn/Article/CDMD-10022-1014324722.htm
|
| [28] |
方精云, 王襄平, 沈泽昊, 唐志尧, 贺金生, 于丹, 江源, 王志恒, 郑成洋, 朱江玲, 郭兆迪. 植物群落清查的主要内容、方法和技术规范. 生物多样性, 2009, 17(6): 533-548. |
| [29] |
曹梦, 潘萍, 欧阳勋志, 臧颢, 宁金魁, 郭丽玲, 李杨. 飞播马尾松林林下植被组成、多样性及其与环境因子的关系. 生态学杂志, 2018, 37(1): 1-8. |
| [30] |
谭凌照, 范春雨, 范秀华. 吉林蛟河阔叶红松林木本植物物种多样性及群落结构与生产力的关系. 植物生态学报, 2017, 41(11): 1149-1156. |
| [31] |
Hui G, Pommerening A. Analysing tree species and size diversity patterns in multi-species uneven-aged forests of Northern China. Forest Ecology and Management, 2014, 316: 125-138. DOI:10.1016/j.foreco.2013.07.029 |
| [32] |
王静, 梁军, 焦一杰, 张星耀. 杨树人工林林木个体大小比数与溃疡病发生程度的关系. 林业科学, 2012, 48(11): 57-62. DOI:10.11707/j.1001-7488.20121109 |
| [33] |
周永斌, 殷有, 殷鸣放, 张飞. 白石砬子国家级自然保护区天然林的自然稀疏. 生态学报, 2011, 31(21): 6469-6480. |
| [34] |
汤孟平, 娄明华, 陈永刚, 徐文兵, 赵明水. 不同混交度指数的比较分析. 林业科学, 2012, 48(8): 46-53. |
| [35] |
董灵波, 刘兆刚, 马妍, 倪宝龙, 李元. 天然林林分空间结构综合指数的研究. 北京林业大学学报, 2013, 35(1): 16-22. |
| [36] |
游诗雪, 张超, 库伟鹏, 朱弘, 赵明水, 余树全. 1996-2012天目山常绿落叶阔叶混交林乔木层群落动态. 林业科学, 2016, 52(10): 1-9. DOI:10.11707/j.1001-7488.20161001 |
| [37] |
Morin X, Fahse L, De Mazancourt C, Scherer-Lorenzen M, Bugmann H. Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics. Ecology Letters, 2014, 17(12): 1526-1535. DOI:10.1111/ele.12357 |
| [38] |
葛结林, 熊高明, 邓龙强, 赵常明, 申国珍, 谢宗强. 湖北神农架山地米心水青冈-多脉青冈混交林的群落动态. 生物多样性, 2012, 20(6): 643-653. |
| [39] |
郭利平, 姬兰柱, 王珍, 王智轩. 长白山红松阔叶林不同演替阶段优势种的变化. 应用生态学报, 2011, 22(4): 866-872. |
| [40] |
白超, 惠刚盈. 林木直径大小多样性量化测度指数的比较研究. 林业科学研究, 2016, 29(3): 340-347. DOI:10.3969/j.issn.1001-1498.2016.03.005 |
| [41] |
Lei X D, Wang W F, Peng C H. Relationships between stand growth and structural diversity in spruce-dominated forests in New Brunswick, Canada. Canadian Journal of Forest Research, 2009, 39(10): 1835-1847. DOI:10.1139/X09-089 |
| [42] |
张璐, 唐建军, 叶宝兴, 王根轩, 陈欣. 植物群落密度调控研究进展. 生态学报, 2010, 30(2): 455-461. |
| [43] |
张春雨, 赵秀海, 赵亚洲. 长白山温带森林不同演替阶段群落结构特征. 植物生态学报, 2009, 33(6): 1090-1100. DOI:10.3773/j.issn.1005-264x.2009.06.009 |
| [44] |
Lebrija-Trejos E, Meave J A, Poorter L, Pérez-García E A, Bongers F. Pathways, mechanisms and predictability of vegetation change during tropical dry forest succession. Perspectives in Plant Ecology, Evolution and Systematics, 2010, 12(4): 267-275. DOI:10.1016/j.ppees.2010.09.002 |
| [45] |
夏富才, 姚大地, 赵秀海, 潘春芳, 汪金松, 张杨. 长白山北坡阔叶红松林空间结构. 东北林业大学学报, 2009, 37(10): 5-7. DOI:10.3969/j.issn.1000-5382.2009.10.002 |
| [46] |
夏富才, 赵秀海, 彭道黎, 倪瑞强, 汪金松, 范娟. 长白山次生杨桦林林分结构. 华南农业大学学报, 2013, 34(3): 383-387. |
| [47] |
彭辉, 周红敏, 王宏翔, 赵中华, 惠刚盈. BCI 50 hm2森林大样地2010年活立木空间结构分析. 林业科学研究, 2017, 30(6): 895-901. |
| [48] |
李晓宇, 廖嘉星, 侯嫚嫚, 范秀华. 不同尺度下长白山次生杨桦林群落系统发育结构研究. 北京林业大学学报, 2016, 38(12): 14-20. |
| [49] |
李建, 彭鹏, 何怀江, 谭凌照, 张新娜, 吴相菊, 刘兆刚. 采伐对吉林蛟河针阔混交林空间结构的影响. 北京林业大学学报, 2017, 39(9): 48-57. |